Infection characteristics among Serratia marcescens capsule lineages
Mark T. Anderson, Stephanie D. Himpsl, Leandra G. Kingsley, Sara N. Smith, Michael A. Bachman, Harry L. T. Mobley

TL;DR
This study explores how different capsule types in Serratia marcescens affect infection and survival in the host, focusing on clinical strains and their resistance to immune defenses.
Contribution
The study identifies specific capsule lineages in S. marcescens that confer resistance to macrophage phagocytosis, impacting infection fitness.
Findings
S. marcescens capsule types KL1 and KL2 show resistance to macrophage internalization due to sialylation.
Clinical isolates of multiple capsule types demonstrate capsule-dependent colonization in organs during bacteremia.
Capsule diversity influences infection outcomes in both lung and bloodstream infections.
Abstract
Serratia marcescens is a healthcare-associated pathogen that can cause severe infections, including bacteremia and pneumonia. The capsule polysaccharide of S. marcescens is a bacteremia fitness determinant, and previous work defined capsule locus (KL) diversity within the species. Strains belonging to KL1 and KL2 capsule clades produce sialylated polysaccharides and represent the largest subpopulation of isolates from clinical origin. In this study, the contribution of these and other S. marcescens capsules to infection was determined in animal and cellular models. Using a murine model of primary bacteremia, clinical isolates of multiple KL types demonstrated capsule-dependent colonization of the spleen, liver, and kidney following tail vein inoculation. Similar results were observed using a bacteremic pneumonia model, in that all tested strains of clinical origin demonstrated a…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
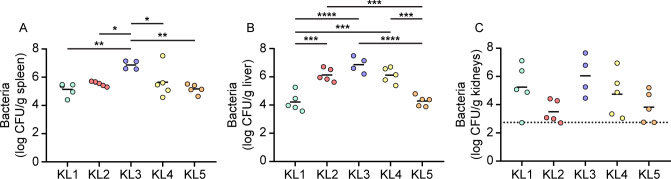 Fig 1
Fig 1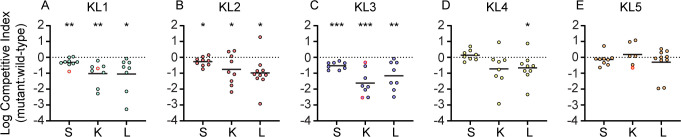 Fig 2
Fig 2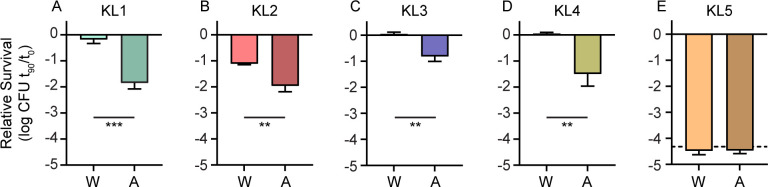 Fig 3
Fig 3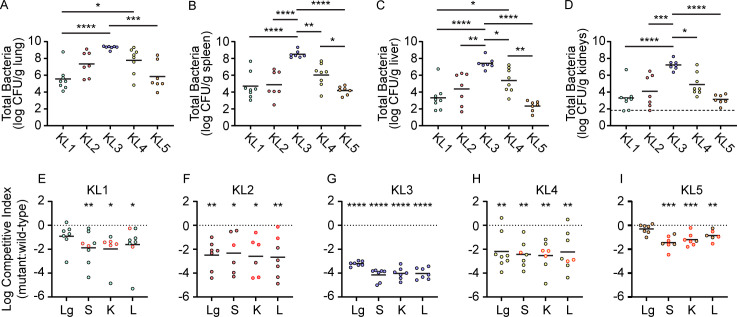 Fig 4
Fig 4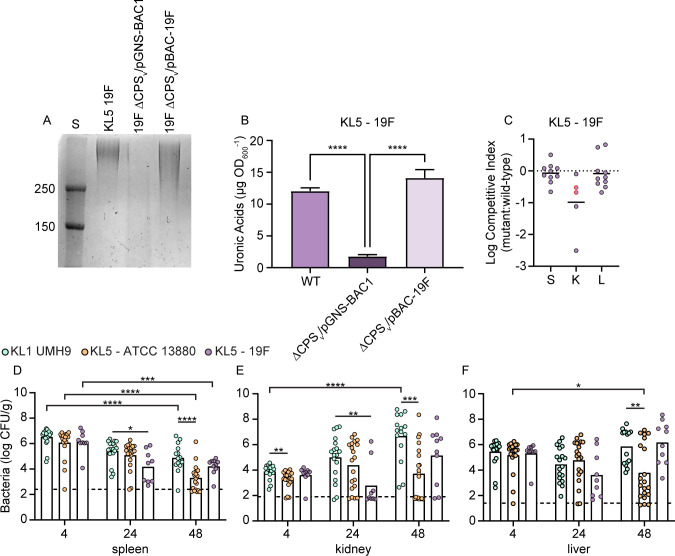 Fig 5
Fig 5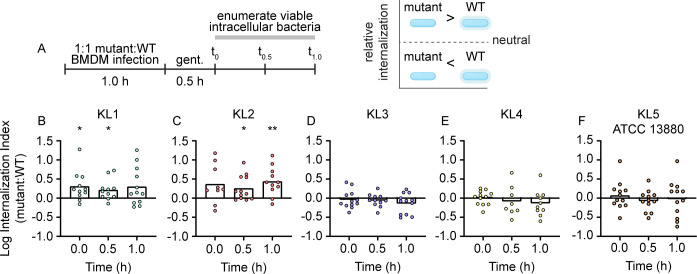 Fig 6
Fig 6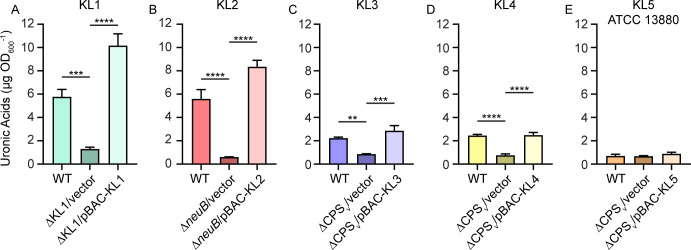 Fig 7
Fig 7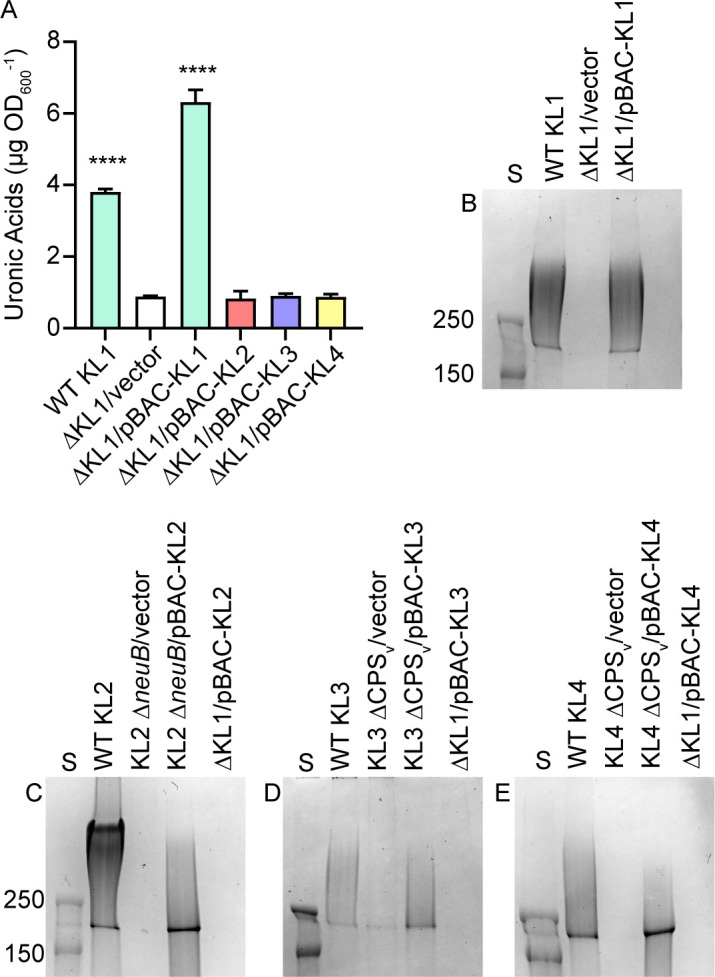 Fig 8
Fig 8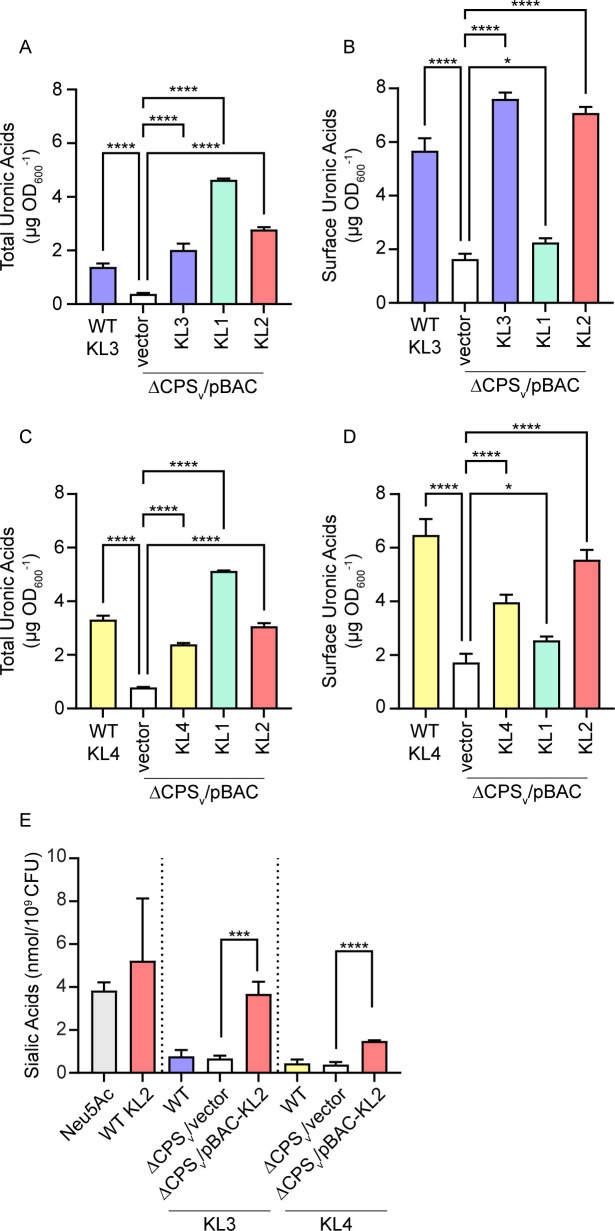 Fig 9
Fig 9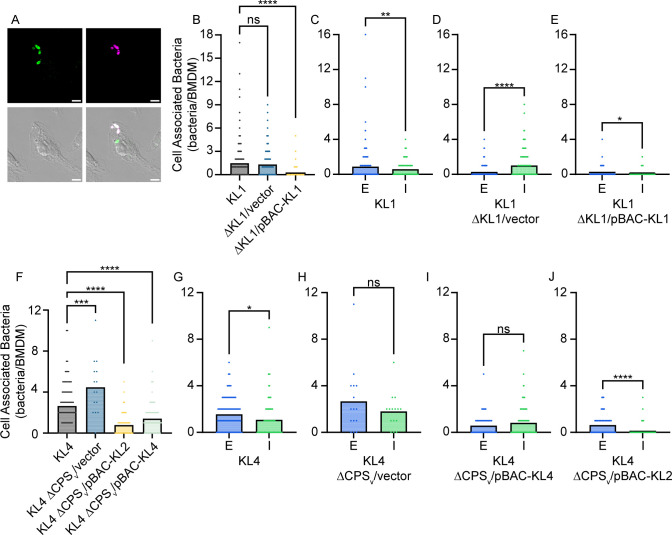 Fig 10
Fig 10| Name | Genotype/description | Source or reference |
|---|---|---|
| Bacteria, KL clade | ||
| UMH9, KL1 | Wild type | ( |
| ΔCPSv:: | ( | |
| ΔKL1 | This study | |
| Δ | ( | |
| gn773, KL2 | Wild type | ( |
| Δ | This study | |
| UMH7, KL3 | Wild type | ( |
| ΔCPSv:: | This study | |
| UMH11, KL4 | Wild type | ( |
| ΔCPSv:: | This study | |
| ATCC 13880, KL5 | Wild type | Pond water, ATCC |
| ΔCPSv:: | This study | |
| 19F, KL5 | Wild type | Frog, USDA-ARS Culture Collection (NRRL) |
| ΔCPSv:: | This study | |
| KZ2, ND | Wild type | Honeybee ( |
| KZ11, ND | Wild type | Honeybee ( |
| KZ19, ND | Wild type | Honeybee ( |
| Plasmids | ||
| pBAC-KL1 | 23.0 kb, 18-ORF insert encoding KL1 | This study |
| pBAC-KL2 | 20.5 kb, 16-ORF insert encoding KL2 | This study |
| pBAC-KL3 | 16.7 kb, 13-ORF insert encoding KL3 | This study |
| pBAC-KL4 | 18.3 kb, 15-ORF insert encoding KL4 | This study |
| pBAC-KL5 (ATCC 13880) | 15.4 kb, 11-ORF insert encoding KL5 | This study |
| pBAC-KL5 (19F) | 15.4 kb, 11-ORF insert encoding KL5 | This study |
| pBAD | pBAD18-km with a 1.7 kb insert containing the | This study |
- —National Institutes of Healthhttp://dx.doi.org/10.13039/100000002
- —National Institutes of Healthhttp://dx.doi.org/10.13039/100000002
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsStreptococcal Infections and Treatments · Infections and bacterial resistance · Microbial infections and disease research
INTRODUCTION
Serratia marcescens is one of the common causes of bacteremia and pneumonia among gram-negative bacterial species (1, 2), and an estimated 100,000 deaths (42,000 associated with drug resistance) were due to Serratia species in 2019 alone (3, 4). Many of these infections occur in individuals with pre-existing conditions or during prolonged hospital stays. Pediatric populations are also vulnerable to severe S. marcescens infections, often occurring as nosocomial outbreaks (5–7). In addition to the systemic and life-threatening infections that are the focus of this work, S. marcescens is capable of a wide range of other pathogenic interactions with both human and non-human hosts (8–11). The clinical significance of S. marcescens is contrasted by the prevalence of the species in many environments, with isolation sources ranging from soil, water, plants, and insects (8) and highlighting the range of niches in which the organism thrives.
Genomic studies investigating the population structure of S. marcescens have defined the species-level diversity and have identified distinct lineages within the species. Some lineages have stringent correlation with clinical sources, consistent with niche adaptation to the infection environment and supported by a discrete repertoire of accessory genomic elements enriched within these clades (12–15). Other lineages are conversely associated with non-clinical or environmental sources. The clinical lineages have a higher proportion of antimicrobial resistance genes, and there is evidence for substantial propagation of drug-resistant clades over time and geographic location (12, 14, 16). The major infection-associated genotypes are now represented by hundreds of sequenced strains, enabling experimental examination of phenotypes that are predicted to impact S. marcescens pathogenesis. Our own work has also demonstrated a distinction between clinical and environmental S. marcescens lineages strictly through comparison of the locus encoding capsule polysaccharide (CPS) (17).
The CPS for one S. marcescens capsule type was characterized as a critical fitness determinant during bloodstream infection (18). Like other encapsulated Enterobacterales species, the CPS of S. marcescens is encoded in a genomic locus that varies extensively between isolates (17). Our comparison of capsule loci (KLs) from infection isolates determined that clades KL1 and KL2 were overrepresented among a cohort of >300 genomes and that strains from both clades produced sialylated CPS. Ketodeoxynonulonic acid (KDN) was the predominant sialic acid identified from KL1 and KL2 strains, but a minor proportion of N-acetylneuraminic acid (Neu5Ac) was also detected. In addition to these predominant clinical capsule types, less abundant capsule clades from clinical or non-infection sources were also defined. In this work, we sought to determine the infection characteristics of S. marcescens strains representing five different capsule clades using both animal and cellular model systems.
RESULTS
Infectivity of S. marcescens isolates following bloodstream inoculation
Capsule was previously determined to be an important fitness factor based on experiments using a single S. marcescens bacteremia clinical isolate belonging to clade KL1 (17, 18). As an initial assessment of infection capability for isolates differentiated by capsule type, strains selected from clades KL1–KL5 (Table 1) were inoculated into the bloodstream of mice via tail vein injection (TVI), and bacterial survival was measured at 24 h. KL1 strain UMH9 stably colonized the spleen, liver, and kidneys in a manner consistent with previous results (19) (Fig. 1). The KL2–KL5 strains also colonized blood-filtering organs at 24 h, but significant variation in bacterial burdens was observed among the strains in both the spleen and liver (Fig. 1A and B), with KL3 bacteremia isolate UMH7 consistently achieving the highest density in both organs. The KL2 (gn773) and KL4 (UMH11) bacteremia isolates were similarly elevated in liver compared to KL1 and the S. marcescens type strain ATCC 13880 (KL5), a pond water isolate. Although variability in the kidneys was higher than the other organs (Fig. 1C), the KL2 and KL5 strains trended toward lower colonization levels, approaching the limit of detection. Thus, while all clinical and non-clinical S. marcescens strains were capable of bacteremia, significant organ-specific colonization differences were observed.
*Strain variations in organ colonization following TVI bacteremia. S. marcescens strains were inoculated into C57BL/6J mice (n = 5) via TVI, and bacterial colonization in spleen (A), liver (B), and kidneys (C) was determined by viable counts. Log-transformed mean bacterial burdens are indicated by the solid lines. Statistical significance was assessed by ordinary one-way analysis of variance with Tukey’s multiple comparison test. *P < 0.05, **P < 0.01, ***P < 0.001, ***P < 0.0001. The dotted line in panel C represents the highest value among samples that were at or below the limit of detection.
Capsule contributions to bacterial survival following TVI
Acapsular mutants of each KL type were next generated to test the contribution of CPS to bacteremia across strains (Table 1). KL1, KL3, KL4, and KL5 strains were mutated such that the variable and cladespecific regions of each KL (CPS_v_) (Fig. S1A) (17) were deleted and replaced with an insert fragment encoding kanamycin resistance. Attempts to construct a similar ΔCPS_v_ mutation in the KL2 strain were unsuccessful, despite multiple efforts using different mutagenic systems. As an alternative, a capsule-null phenotype was achieved by disrupting the neuB gene encoding the sialic acid synthase within the KL2 CPS_v_ region, similar to a previously described capsule-null neuB mutant of KL1 (17). Initial assessments of these five mutants confirmed that the KL mutations disrupted high-molecular-weight CPS production, eliminated extracellular CPS uronic acids, and did not prevent O-antigen synthesis (Fig. S1). The only exception was KL5 ATCC 13880, which did not yield detectable CPS or O-antigen from either the wild-type or the ΔCPS_v_ mutant.
The relative fitness of each capsule mutant was first measured in comparison to the wild-type parent strain in TVI mixed infections via competitive index (CI). All KL types under these mixed inoculum parameters exhibited similar total burdens in the spleen, kidneys, and liver of infected mice (Fig. S2). For each strain with detectable CPS production in vitro (Fig. S1), a significant competitive disadvantage in survival after 24 h was observed for mutants lacking capsular genes (Fig. 2A through D). The fitness advantage provided by KL4 CPS was only significant for bacteria in the liver but showed a similar trend in kidneys. Combined, these results demonstrate that the capsule is important for bacterial survival across multiple clinical S. marcescens isolates and capsular clades. In contrast, the KL5 strain demonstrated no significant change in fitness upon CPS_v_ mutation (Fig. 2E). This result was anticipated, given the lack of CPS associated with this strain. However, the six genes present in the ATCC 13880 KL5 CPS_v_ region (Fig. S1) all appeared to be uninterrupted in the genome sequence (NZ_CP072199.1), and thus, their functional significance was unclear. To further evaluate contributions of each CPS type to bloodstream fitness, strains were exposed to human serum for 90 min followed by enumeration of viable bacteria. KL1–KL4 CPS provided resistance to the bactericidal activity of serum that was at least sixfold greater than acapsular mutant derivatives (Fig. 3A through D). No significant difference in serum susceptibility was observed between the KL5 strains (Fig. 3E).
*Requirement for capsule during TVI bacteremia. (A–E) CI for bacteria recovered from 24 h mixed strain competition TVI infections in C57BL/6J mice (two cohorts of five mice, n = 10). Solid lines represent the mean of log-transformed values. Red outlined symbols indicate CI for which mutant bacteria were recovered at or below the limit of detection. Statistical significance was determined by one-sample t-test with a hypothesized mean value of zero (dotted line), representing neutral fitness. Points below the dotted line represent samples in which the mutant strain was outcompeted by wild-type bacteria. *P < 0.05, **P < 0.01, **P < 0.001. Abbreviations: K, kidney; L, liver; S, spleen.
*Capsules of clinical isolates protect from serum bactericidal activity. (A–E) The survival of wild-type (W) and acapsular (A) mutant strains was determined in the presence of 40% human serum after 90 min exposure relative to time 0 with bars representing mean values (n = 3). The dashed line indicates the limit of detection, where relevant. Statistical significance was assessed by Student’s t-test. **P < 0.01, **P < 0.001.
Capsule contributions to bacteremic pneumonia
Bacteremia frequently originates from localized infections that disseminate and become systemic (20), as opposed to primary bacteremia in which organisms gain direct access to the bloodstream via an exogenous source such as a hypodermic needle or an intravenous catheter. A pneumonia model with secondary bacteremia was therefore developed based on a previously described K. pneumoniae model (21). Following retropharyngeal co-inoculation of wild-type and capsule mutant strains into mice, colonization of the lungs was observed at 24 h post-inoculation (Fig. 4A). The spleen, kidneys, and liver were also colonized at this time point (Fig. 4B through D) and to levels that approximated those observed in the primary bacteremia model (Fig. 1). Escape of S. marcescens from the lungs therefore occurs readily and results in stable organ colonization. The overall trends in bacterial burdens of the spleen, kidneys, and liver between this dissemination-dependent route and the TVI route were also similar for individual KL types in that KL3 bacteria exhibited the highest density followed by KL2 and KL4, then KL1 and KL5 ([Fig. 1A through C and 4B through D](#F1 F4)). Bacterial accumulation at systemic sites also correlated to primary lung burden when assayed at the time of sacrifice (Fig. 4A). Therefore, primary lung burden may influence dissemination kinetics and subsequent organ colonization, in addition to any differences in infection capacity between strains.
*Requirement for capsule during bacteremic pneumonia. Combined wild-type and capsule mutant bacteria recovered from the lung (A), spleen (B), liver (C), and kidneys (D) of C57BL/6J mice (two cohorts of five mice, n = 10) following mixed strain competition infections (24 h). Solid lines represent the mean of log-transformed viable bacteria, and the dotted line in panel D indicates the highest value among samples that were at or below the limit of detection from kidneys. Differences in bacterial burdens between strains were assessed by one-way analysis of variance with Tukey’s multiple comparison test. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001. (E–I) CIs comparing relative survival of capsule mutant and wild-type bacteria in the lung (Lg), spleen (S), kidneys (K), and liver (L) for the infections shown in panels A–D. Symbols with red outlines denote CIs from which capsule mutant strains were recovered at or below the limit of detection. Fitness defects were assessed by one-sample t-test against the hypothetical null value of 0 (dotted lines) representing neutral fitness. Points below the dotted line represent samples in which the mutant strain was outcompeted by wild-type bacteria. *P < 0.05, **P < 0.01, ***P < 0.001, ***P < 0.0001.
To define the requirement for capsule in pneumonia, the relative recovery of capsule mutant and wild-type bacteria for KL1-5 strains was determined. The four acapsular strains of clinical origin all demonstrated a severe competitive disadvantage compared to parental strains (Fig. 4E through H), with the mean recovery of capsule-deficient strains being ca. 100-fold or less than wild type for most organ and strain combinations. Thus, in both primary and secondary bacteremia models, the capsule is a critical fitness determinant across human-associated S. marcescens. Unexpectedly, a significantly competitive disadvantage for the capsule mutant derivative of KL5 ATCC 13880 was also observed in the spleen, kidneys, and liver (Fig. 4I). Given the contrast between these results and those from TVI (Fig. 2E), the mutated KL5 capsular genes could play an active role in bacterial survival during lung dissemination. However, wild-type ATCC 13880 bacteria lacked a competitive advantage in the lung, whereas the clinical strains demonstrated at least an eightfold advantage in the lung compared to isogenic acapsular derivatives (Fig. 4E through H). Additional efforts to identify ATCC 13880 CPS by microscopy during repeated passage under selective pressure in human serum failed to provide evidence that CPS synthesis could be induced in this strain (Fig. S3).
KL5 CPS does not contribute to primary bacteremia
The lack of KL5 CPS from ATCC 13880 was a limitation to understanding the role of CPS from non-clinical S. marcescens. To assess whether other KL5 strains produce CPS, a second representative designated 19F (Table 1) was acquired and characterized. A high-molecular-weight polysaccharide was detected from 19F (Fig. 5A), and this strain had abundant surface-associated uronic acids (Fig. 5B), demonstrating that 19F synthesizes CPS. Three additional environmental strains that were originally isolated from honeybees (Table 1) also produce acidic CPS (Fig. S4), solidifying the conclusion that the acapsular phenotype of ATCC 13880 is not conserved among environmental strains. No significant fitness difference in TVI CI was observed between wild-type 19F and a ΔCPS_v_ mutant derivative (Fig. 5C), supporting the conclusion that the KL5 CPS contributes minimally to 19F bacteremia fitness. We hypothesized that the lack of a CPS-mediated fitness advantage for either ATCC 13880 or 19F would result in enhanced clearance of these strains compared to an encapsulated strain such as KL1 UMH9, and therefore, survival following TVI was assessed over time. Both KL5 strains trended toward enhanced clearance compared to KL1, with ATCC 13880 showing significantly lower bacterial burdens than KL1 in all organs at 48 h post-inoculation and an overall loss of bacteria over time in the liver (Fig. 5D through F). Recovery of strain 19F was also lower than KL1 at 24 h in the spleen and kidney but did not show significant differences in the liver at any time point. Additionally, KL1 was the only strain capable of significant expansion over the course of the experiment, with high levels of bacteria found in the kidney by 48 h (Fig. 5E). Clearance of all strains appeared to be most effective in the spleen since bacterial burdens decreased significantly over time (Fig. 5D).
*CPS does not contribute to the fitness of KL5 strain 19F. (A) Total polysaccharides prepared from wild-type and capsule-null derivatives of KL5 strain 19F were separated by SDS-PAGE and stained with alcian blue. The 19F capsule mutant harbored the pGNS-BAC1 vector control plasmid or recombinant plasmid containing a cloned copy of 19F KL. Pre-stained protein molecular weight standards (S) of known molecular weight are shown (kDa). (B) Cell-associated uronic acids were quantitated from 19F strains in comparison to a glucuronic acid standard curve. Statistical significance was calculated by one-way analysis of variance (ANOVA) with Dunnett’s multiple comparison test relative to 19F ΔCPSv/pGNS-BAC1. (C) Strains 19F and 19F ΔCPSv were inoculated at an equal ratio into C57BL/6J mice (n = 10), and CI was determined from bacteria recovered from the spleen (S), kidney (K), and liver (L). Red outlined symbols indicate the CI for which mutant bacteria were recovered at or below the limit of detection. Points below the dotted line represent samples in which the mutant strain was outcompeted by wild-type bacteria. Mean log competitive indices (solid lines) were not significantly different from the hypothesized value of 0 representing neutral fitness, as determined by one-sample t-test. (D–F) Mice from at least two independent cohorts (n = 10–20) were inoculated with the indicated strains via the TVI route, and bacterial burdens in the spleen, kidneys, and liver were determined. The mean values of log-transformed numbers of viable bacteria recovered are indicated by the bars, and dashed lines denote the highest value among samples that were at or below the limit of detection. Statistical significance was assessed by one-way ANOVA with Dunnett’s multiple comparison test. Adj. P values: *P < 0.05, **P < 0.01, ***P < 0.001, ***P < 0.0001.
Sialylated CPS protects S. marcescens during macrophage interactions
We previously concluded that KL1 CPS has anti-phagocytic properties based on data demonstrating that a KL1 acapsular derivative was internalized more readily by the U937 monocytic cell line compared to wild-type bacteria (17). To determine whether CPS from other KL types also inhibited cellular uptake, intracellular bacteria were quantitated after incubation with murine bone marrow-derived macrophages (BMDMs). The relative number of viable intracellular acapsular mutants was compared to wild-type bacteria at three time points following treatment with gentamicin to kill extracellular bacteria and was calculated as an internalization index (Fig. 6A). For both KL1 and KL2 sialylated capsule types, higher numbers of viable and internalized acapsular bacteria were recovered compared to wild type (Fig. 6B and C). Therefore, both KL1 and KL2 CPS contribute to macrophage phagocytosis resistance. In contrast, none of the non-sialylated CPS KL types exhibited a significant difference under the same conditions, as evidenced by neutral internalization indices for KL3, KL4, and KL5 bacteria (Fig. 6D through F). The comparative lack of CPS-dependent phagocytosis resistance for the KL3 and KL4 clinical strains, in particular, suggests an important role for sialylated S. marcescens CPS in innate immune interactions and may be one of multiple contributing factors to the successful adaptation of these lineages to infection.
*Sialylated CPS protects S. marcescens from macrophage internalization. (A) Schematic for the design and interpretation of BMDM competition infection experiments. (B–F) Murine BMDMs were co-infected at a 1:1 ratio with wild-type (WT) and capsule mutant derivatives of each clade followed by enumeration of viable intracellular bacteria. The relative number of intracellular mutant and WT bacteria was calculated as an internalization index. Bars represent the mean of log-transformed internalization indices, and significant deviation from the hypothetical value of 0 representing equivalent internalization was determined by one-sample t-test. *P < 0.05, *P < 0.01.
KL cloning and non-native CPS synthesis
KL1–KL4 and two KL5 loci were cloned into the bacterial artificial chromosome (BAC) pGNS-BAC1 (22). The cloned regions ranged from 15 to 23 kb in length (Table 1) and consisted of the entire intergenic sequence upstream of galU, the five-gene conserved KL region, and all clade-specific KL open reading frames (Fig. S1A). Transformation of the pBAC-KL plasmids into their respective capsule mutant strains resulted in complete restoration of extracellular uronic acid production for strains KL1–KL4 (Fig. 7A through D) and the 19F strain of KL5 (Fig. 5B), but not ATCC 13880 (Fig. 7E). Thus, the engineered pBAC-KL constructs are functional and sufficient to restore CPS production in their cognate strains.
*Genetic complementation of KL mutations. (A–E) Capsule production by wild-type (WT) and capsule mutant S. marcescens strains representative of clades KL1–KL5 was measured by quantifying extracellular uronic acids and based on a standard curve of glucuronic acid. Statistical significance was assessed relative to mutant strains harboring the vector control plasmid by one-way ANOVA with Dunnett’s multiple comparison test. Adj. P values: **P < 0.01, ***P < 0.001, ***P < 0.0001.
Heterologous CPS synthesis was first attempted in KL1 UMH9 because it is the strain that we have characterized most extensively. A second acapsular KL1 derivative was generated for this purpose that harbored a deletion of the entire KL1 from the five-gene conserved region (galU, galF, wza, wzb, and wzc) through the clade-specific variable locus (Fig. S1A). BAC constructs harboring KL1–KL4 were transformed into the UMH9 ΔKL1 strain, and CPS production was quantitated via uronic acids. Unexpectedly, none of the BAC constructs harboring KL2–KL4 yielded significant increases in uronic acids compared to the ΔKL1 vector control (Fig. 8A). Total extracellular polysaccharides were also isolated from these strains and resolved by SDS-PAGE. Consistent with the uronic acid quantitation, each of the pBAC-KL constructs was able to restore production of CPS when introduced into the native acapsular strains but not in the ΔKL1 background (Fig. 8B through E). We hypothesized that the Wzi protein, required for surface attachment of CPS and encoded outside the KL (17), may be involved in strainspecific CPS display. However, a cloned copy of the KL3 wzi gene expressed in the ΔKL1/pBAC-KL3 strain does not restore production of either cell-free or cell-associated uronic acids in this background (Fig. S5A) but does restore surface association of KL1 CPS in a KL1 Δwzi mutant (Fig. S5B). Therefore, the inability to heterologously produce CPS in KL1 UMH9 is due to a presently unknown limitation but may occur prior to polysaccharide surface translocation and attachment.
*Synthesis of non-native CPS is restricted in KL1. (A) Extracellular uronic acids were quantitated from the wild-type (WT) KL1 strain and a KL1 deletion mutant (ΔKL1) harboring the vector control plasmid pGNS-BAC1 or plasmids containing the KL1–KL4 regions. Uronic acids were quantitated in comparison to a glucuronic acid standard curve. Statistical significance was determined using ordinary one-way ANOVA with Dunnett’s multiple comparison test against the negative control strain. ***Adj. P < 0.0001. (B–E) Total bacterial polysaccharides from WT and capsule-null derivatives were separated by SDS-PAGE and stained with alcian blue. Capsule mutants harbored either the pGNS-BAC1 vector control plasmid or a recombinant plasmid with a cloned copy of the native KL. Recombinant KL plasmids were also expressed from a capsule-null mutant derivative of KL1 (ΔKL1). Pre-stained protein molecular weight standards (S) were electrophoresed on each gel with molecular weights shown in kilodaltons.
To determine if non-native CPS production was possible in other lineages, BAC constructs containing KL1 and KL2 were transformed into the KL3 and KL4 ΔCPS_v_ mutants. KL1 and KL2 CPS were quantitated at or above the level of wild-type strains, and the native pBAC-KL3 or pBAC-KL4 complemented ΔCPS_v_ mutants as measured by total cell-associated plus extracellular uronic acids in both backgrounds (Fig. 9A and C). However, only a minor fraction of KL1 CPS was surface associated compared to CPS from the other BAC constructs (Fig. 9B and D). Since abundant non-native KL1 CPS is likely released from the surface in these scenarios, further KL1 genetic combinations were not pursued. The pBAC-KL2 construct, in contrast, yielded surface-associated KL2 CPS at levels similar to the native pBAC-KL3 and pBAC-KL4 constructs. Sialic acids were next quantitated as an additional measure of KL2 CPS in KL3 and KL4 strains. Wild-type KL3 and KL4 strains yielded background levels of sialic acids by the thiobarbituric acid assay, and both strains carrying pBAC-KL2 yielded a significant increase in extracellular sialic acids compared to capsule mutant bacteria harboring the vector control plasmid (Fig. 9E). These combined results demonstrate that sialylated and surface-associated KL2 CPS can be synthesized in both KL3 and KL4 strains.
*Non-native synthesis of sialylated CPS in KL3 and KL4 strains. (A–D) Total uronic acids (A and C) and surface-associated uronic acids (B and D) were quantitated from wild-type (WT) and ΔCPSv mutant strains. Mutant bacteria harbored either the control plasmid pGNS-BAC (vector) or recombinant BAC plasmids containing the indicated KL. Quantitation was based on a standard curve with glucuronic acid. Statistical significance was assessed by one-way ANOVA with Dunnett’s multiple comparison test against ΔCPSv mutants carrying the vector control plasmid. Adj. P values: *P < 0.05, ***P < 0.001, ***P < 0.0001. (E) Sialic acids were quantitated by thiobarbituric acid assay and normalized by CFU between strains. Purified Neu5Ac and CPS from wild-type KL2 bacteria were used as positive controls. KL3 and KL4 ΔCPSv mutants harbored either the empty pGNS-BAC plasmid or recombinant plasmids expressing KL2. Statistical analysis was constrained within the dotted lines and was performed as described for panels A–D.
Non-native KL2 CPS limits macrophage internalization
To further investigate the specific impact of S. marcescens sialylated CPS on macrophage interactions, immunofluorescence microscopy was used to quantify intracellular and extracellular bacteria associated with BMDM (Fig. 10A). KL1 bacteria with and without the native KL1 CPS were first analyzed to establish the approach. Both wild-type and the ΔKL1 mutant had a similar number of total bacteria (extracellular + intracellular) associated with BMDM on a per-cell basis (Fig. 10B). However, a significantly higher proportion of these bacteria were extracellular for wild type, whereas the ΔKL1 derivative was predominantly intracellular (Fig. 10C and D; Fig. S6). These results confirm the previous gentamicin protection assay findings (Fig. 6) and solidify the role of KL1 CPS in resisting macrophage phagocytosis. Genetic complementation with pBAC-KL1 restored the extracellular-to-intracellular relationship observed for the wild-type strain (Fig. 10E) but substantially reduced the total number of bacteria per BMDM (Fig. 10B), potentially due to CPS hyperproduction (Fig. 7A).
*Ectopic expression of KL2 genes confers phagocytosis resistance to a non-sialylated strain. (A) Representative images of BMDM infected with the wild-type KL1 strain for 60 min followed by differential immunofluorescence and microscopy. Extracellular bacteria were labeled with an Alexa Fluor-647 conjugated secondary antibody. BMDM were then permeabilized, and all bacteria were exposed to an Alexa Fluor-488 conjugated secondary antibody. Extracellular bacteria fluoresce in both channels and appear white in the composite image, while intracellular bacteria appear green. Scale bars are 5 µm. (B and F) Quantitation of total BMDM-associated bacteria (extracellular + intracellular). Points represent the number of bacteria associated with individual BMDM cells, and statistical significance was assessed by one-way ANOVA with Dunnett’s multiple comparison test against the KL1 and KL4 wild-type strains, respectively. Adj. P values: ***P < 0.001, ****P < 0.0001. (C–E and G–J) Quantitation of extracellular (E) and intracellular (I) BMDM-associated bacteria for KL1 and KL4 wild-type and CPS mutant derivative strains. Statistical significance was assessed by unpaired t-test. P values: *P < 0.05, **P < 0.01, ***P < 0.001, ***P < 0.0001. ns, non-significant.
The ability to synthesize KL2 CPS in both the KL3 and KL4 strains allowed us to test the hypothesis that sialylated KL2 CPS confers phagocytosis resistance to these normally non-sialylated lineages. Loss of the native KL3 and KL4 CPS significantly increased the per-cell number of total BMDM-associated bacteria compared to the parental strains (Fig. 10F; Fig. S7), but as expected, the relationship between the number of extracellular and intracellular bacteria remained largely unchanged between wild-type and ΔCPS_v_ mutant bacteria (Fig. 10G and H; Fig. S7B and C). Thus, the KL3 and KL4 CPS have minimal impact on BMDM phagocytosis, as demonstrated in Fig. 6. Unfortunately, complementation of the KL3 ΔCPS_v_ mutant with either pBAC-KL3 or pBAC-KL2 significantly reduced the overall detectable number of cell-associated bacteria to a level that prevented reliable assessment of intracellular and extracellular trends (Fig. S7). This was not the case for KL4, and in fact, complementation with pBAC-KL4 confirmed that native KL4 synthesis does not result in significant differences between the number of intracellular and extracellular bacteria per cell (Fig. 10I), similar to the KL4 ΔCPS_v_ mutant harboring the vector control (Fig. 10H). In contrast, KL4 ΔCPS_v_ harboring pBAC-KL2 shifted localization to a significant majority of extracellular bacteria compared to nearly undetectable numbers of intracellular bacteria (Fig. 10J). Sialylated KL2 CPS is therefore capable of restricting macrophage phagocytosis of both its native strain and a normally non-sialylated S. marcescens lineage.
DISCUSSION
In this study, S. marcescens strains isolated from clinical and non-clinical sources were assayed for survival characteristics during infection. These strains were selected as representatives of five sequence-defined capsule clades. While all strains demonstrated an ability to infect mice using two bacteremia models, significant differences in organ colonization and immune cell interactions were observed. Though we attempted to capture a range of diverse isolates within the species, one acknowledged limitation of the study is the use of a single KL representative in most cases. Nonetheless, the capsule is established here as a critical fitness determinant for each of the clinical clades assessed. Given the mounting genomic evidence distinguishing S. marcescens clinical lineages from environmental isolates (12, 14, 15), it is likely that other factors also contribute to the infection fitness of healthcare-associated strains, and our ongoing work aims to identify and characterize such factors encoded within these clinical accessory genomes. Strains belonging to the environmental capsule clade KL5 generally exhibited a lower infection capacity and less dependence on CPS in our models. For type strain ATCC 13880, these observations could be attributed to an inability to synthesize CPS. Repeated laboratory passage of encapsulated bacteria can result in spontaneous loss of capsule due to mutation, but whether this is the case for ATCC 13880 remains to be determined. However, KL5 strain 19F yields abundant CPS yet exhibited no significant fitness cost when the capsule was lost, supporting the conclusion that KL5 CPS plays a minor role in bacteremia.
Two approaches were used to demonstrate that sialic acid-containing CPS of KL1 and KL2 S. marcescens had the greatest protective effect against BMDM internalization. We hypothesize that KDN and Neu5Ac, which were uniquely associated with these capsule clades (17), may therefore have a specific role in influencing Serratia-macrophage interactions. While such molecular interactions, particularly for the more abundant and understudied KDN component, have yet to be demonstrated for S. marcescens, this hypothesis is indirectly supported by experimentally established roles for Neu5Ac-mediated modulation of innate immune cells in other systems (23–25). It is notable that non-sialylated clinical CPS types also have a meaningful but perhaps different role in infection as demonstrated by the significant fitness cost of acapsular KL3 and KL4 derivatives. With the exception of the aforementioned wzi gene, each S. marcescens KL was expected to be sufficient for type-specific CPS synthesis; however, the limited ability or inability to produce non-native CPS in some strains indicates that additional unknown factors are required. KL1 bacteria, in particular, failed to synthesize any of the other CPS, including the closely related CPS of KL2, and KL1 CPS was only poorly surface associated in the other lineages.
In our previous KL comparison, the KL1 and KL2 lineages had the greatest number of representatives in our genome cohort, and both were overwhelmingly composed of infection isolates (17). In the context of the comprehensive S. marcescens genomic architecture published by Ono et al. (12), the UMH9 KL1 and gn773 KL2 strains characterized here both segregate into clade 1. This is notable because clade 1 S. marcescens spp. are almost exclusively hospital-associated or clinical isolates, have a higher number of antimicrobial resistance alleles, and encode a distinct set of accessory genes compared to other lineages. Furthermore, of the 215 strains identified as either KL1 or KL2 in our study, 188 were also included in the Ono et al. study, and all of them were assigned to the clade 1 genomic lineage. This observation independently confirms our conclusion that KL1 and KL2 CPSs are a differential component of these infection-adapted S. marcescens and, together with the results reported here, indicate that sialylated CPSs contribute to the niche-specific characteristics that provide these isolates with a selective advantage during infection.
MATERIALS AND METHODS
Bacterial strains and culture conditions
The S. marcescens strains used in this study are listed in Table 1. Escherichia coli DH10B and DH5α were routinely used for cloning purposes. DH5α harboring helper plasmid pRK2013 (26) or E. coli BW29427 (B. Wanner, unpublished data) cultured in 0.3 mM diaminopimelic acid was used as a donor strain for conjugation. Bacteria were cultured in either lysogeny broth (LB) medium (27) with or without 20 mM glucose or M9 (28) medium supplemented with 1 mM MgSO_4_, 36 µM FeSO_4_, 100 µM CaCl_2_, and 20 mM glucose. Antibiotics for bacterial culture were used at the following concentrations: kanamycin, 50 µg/mL; hygromycin, 200 µg/mL; spectinomycin, 100 µg/mL; gentamicin, 10 and 20 µg/mL; and ampicillin, 100 µg/mL.
Generation of mutants
The S. marcescens ATCC 13880 and UMH7 KL mutations (Table 1) were constructed by recombineering as previously described (18, 29). Briefly, the nptII gene from pKD4 (30) was PCR amplified with oligonucleotides possessing ~50 bp of the 5′ sequence homology to the targeted CPS_v_ region. Electrocompetent recipient strains harboring pSIM18 or pSIM19 (31) were transformed with DpnI-treated PCR products. Kanamycin-resistant transformants were genotyped by PCR and sequenced, then cured of pSIM plasmids prior to use in phenotypic assays. KL mutations in strains UMH11, gn773, and 19F (Table 1) were accomplished via allelic exchange with pTOX11 (32) derivatives as previously described (17). Plasmids were constructed from PCR-amplified fragments using NEB HiFi Assembly and harbored ~700 bp of homologous sequence flanking the nptII allele to facilitate recombination. The unmarked ΔKL1 mutation in strain UMH9 was generated using a similar approach but did not include the nptII insertion. Allelic exchange was performed following conjugation of pTOX plasmids from E. coli donor strain BW29427 into recipient S. marcescens strains as previously described (17, 32). The presence of mutant alleles was confirmed by PCR amplification and sequencing. Transconjugants were also assessed by PCR to ensure Mu phage was not transferred from the BW29427 donor (33). All primer sequences used for PCR amplification in the procedures described above are listed in Table S1.
Genetic complementation
KL sequences were cloned into BAC vector pGNS-BAC1 (22). Each KL region with upstream intergenic sequence was PCR amplified in two fragments using the primers listed in Table S1. Fragments were then cloned into HindIII-digested pGNS-BAC1 using HiFi DNA Assembly and transformed into electrocompetent E. coli DH10B. Recombinant pGNS-BAC1 plasmids were purified by alkaline lysis, and whole-plasmid sequences were determined by Nanopore (SNPsaurus). The resulting pBAC-KL plasmids (Table 1) were transferred to S. marcescens recipient strains via tri-parental mating using helper plasmid pRK2013 (26). Ampicillin and gentamicin were used to select for the loss of E. coli donor strains and the presence of pBAC-KL plasmids in S. marcescens, respectively.
Quantitation of uronic acids and polysaccharide analysis
Extracellular uronic acids of S. marcescens were measured using previously described methods (17, 34, 35). Measurements were based on a standard curve of glucuronic acid and normalized to culture optical density (600 nm). Results are the means from three biological replicates and are representative of at least two independent experiments. Isolation of S. marcescens extracellular polysaccharides and visualization by SDS-PAGE were also performed according to published methods (17, 36), and the results are representative of at least two independent experiments.
Human serum exposure
Bacterial viability following a 90 min incubation in 40% pooled human serum (Innovative Research) was determined as previously described (17). Results are the means from three biological replicates and are representative of three independent experiments.
For the attempted selection of ATCC 13880 CPS synthesis, wild type and the ΔCPS_v_ derivative were passaged every 24 h in LB medium or LB medium supplemented with increasing concentrations of pooled normal human serum (5%, 10%, and 20%) over the course of 72 h. The presence of CPS was assessed via negative stain with Maneval’s reagent (37) and visualized using a Nikon Ti2 widefield microscope with a ×100 objective (University of Michigan Microscopy Core). KL2 bacteria harvested from LB agar were used as a positive control.
Quantitation of sialic acids
Sialic acids were quantitated from S. marcescens strains by thiobarbituric assay as previously described (17, 38). Extracellular polysaccharides were subjected to acid hydrolysis with 0.1 N HCl at 80°C for 60 min, and bacteria were subsequently removed by centrifugation. Hydrolyzed solutions were subjected to periodate oxidation according to the protocol and reacted with thiobarbituric acid. Chromophore extraction was performed with an equal volume of cyclohexanone, and absorbance was measured at 549 nm. The amount of sialic acid was determined with the following formula: (volume [mL] prior to extraction × OD_549_) / 57 = μmole Neu5Ac equivalents. The amount of sialic acid detected was normalized to the bacterial culture density, and 100 µM Neu5Ac served as the positive control. Results are reported as the mean from three biological replicates and representative of two independent experiments.
Murine infections
Murine infections were performed with protocols approved by the University of Michigan Institutional Animal Care and Use Committee and in accordance with Office of Laboratory Animal Welfare guidelines. For TVI mono-infections, male and female 7- to 8-week-old C57BL/6J mice (Jackson Laboratories) were inoculated with 0.1 mL bacterial suspensions in phosphate-buffered saline (PBS) containing a target dose of 5 × 10^6^ CFU. For TVI competition infections, wild-type bacteria were mixed at a 1:1 ratio with mutant strains and delivered at a dose of 5 × 10^6^ total CFU. Mice were euthanized 24 h post-infection, unless otherwise specified, and the spleen, liver, and kidneys were harvested and homogenized. Bacterial counts of the inoculum (input) and organ homogenates (output) were determined by plating serial dilutions on LB agar with or without kanamycin. The CI was determined by the following calculation: (CFU_mutant_/CFU_wild type_)^output^/(CFU_mutant_/CFU_wild type_)^input^. A log-transformed CI of less than zero indicates a competitive disadvantage for mutant bacteria compared to the parental strain.
For the bacteremic pneumonia model, 0.05 mL bacterial suspensions were delivered to the retropharyngeal space of anesthetized 7–8 week-old mice at a target dose of 1 × 10^7^ total CFU. Mice were euthanized 24 h post-inoculation, and the spleen, liver, kidneys, and lung were harvested. CI was determined as described above.
Gentamicin protection assays
Isolation and propagation of BMDM were accomplished using established protocols (39). Monocytes from the femur and tibia bone marrow of 7- to 8-week-old C57BL/6J mice were diluted to 1 × 10^6^ cells/ml in medium containing 15% L929 cell supernatant. At 7 days post-harvest, BMDMs were dissociated from wells with ice-cold 2 mM EDTA in Dulbecco’s phosphate-buffered saline (DPBS) and collected by centrifugation. BMDMs were seeded into 96-well flat bottom plates at 1 × 10^5^ cells/well and incubated at 37°C at 5% CO_2_ for 24 h prior to inoculation with bacteria. Wild-type and kanamycin-resistant mutant strains were mixed in a 1:1 ratio and added to BMDMs at a target multiplicity of infection of 20. Plates were centrifuged briefly and incubated at 37°C for 60 min in 5% CO_2_. The medium was then aspirated, and wells were washed with DPBS. Dulbecco’s modified Eagle medium containing 10% fetal bovine serum and 100 µg/mL gentamicin was added to wells for 30 min at 37°C in 5% CO_2_ followed by removal of gentamicin-containing medium and washing. For time point zero, BMDMs were exposed to 1% saponin at 37°C for 10 min, mixed with 0.1 mL LB medium, then serially diluted and plated on an LB agar plate with and without kanamycin for CFU determination. For all the other time points, BMDMs were incubated in medium containing 10 µg/mL gentamicin until permeabilization and CFU determination. Internalization indices from two independent experiments were calculated as described for CI with internalized bacteria substituting for the infection output parameter.
Immunofluorescence
BMDMs were allowed to adhere to glass coverslips overnight and then infected as described for the gentamicin protection assays with single S. marcescens strains. After 60 min incubation, coverslips were washed with PBS and fixed with 4% paraformaldehyde. PBS with 10% goat serum was used to block coverslips prior to incubation with a 1:100 dilution of the primary antibody, an anti-E. coli polyclonal antibody (Invitrogen, AB_780488) that cross-reacts with S. marcescens. After washing, a 1:400 dilution of goat anti-rabbit secondary antibody conjugated to Alexa Fluor 647 (Invitrogen, AB_2535813) was applied. Coverslips were thoroughly washed and fixed again in paraformaldehyde, followed by permeabilization of BMDM with 0.2% saponin and re-application of the primary antibody. A 1:400 goat anti-rabbit secondary antibody conjugated to Alexa Fluor 488 (Invitrogen, AB_143165) was applied to differentiate internalized bacteria. Imaging was conducted on a Nikon N-SIM A1R confocal microscope (University of Michigan Microcopy Core) using a ×60 objective. All image analysis was performed in the Fiji distribution of ImageJ (40). Intracellular and extracellular bacteria were differentiated by single (488 nm) or dual fluorescence, respectively, and only cell-associated bacteria were counted. Each strain was assayed in at least two independent experiments, and images were collected from a minimum of 15 fields per coverslip.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Wisplinghoff H, Bischoff T, Tallent SM, Seifert H, Wenzel RP, Edmond MB. 2004. Nosocomial bloodstream infections in US hospitals: analysis of 24,179 cases from a prospective nationwide surveillance study. Clin Infect Dis 39:309–317. doi:10.1086/42194615306996 · doi ↗ · pubmed ↗
- 2Sader HS, Streit JM, Carvalhaes CG, Huband MD, Shortridge D, Mendes RE, Castanheira M. 2021. Frequency of occurrence and antimicrobial susceptibility of bacteria isolated from respiratory samples of patients hospitalized with pneumonia in Western Europe, Eastern Europe and the USA: results from the SENTRY Antimicrobial Surveillance Program (2016-19). JAC Antimicrob Resist 3:dlab 117. doi:10.1093/jacamr/dlab 11734671728 PMC 8522161 · doi ↗ · pubmed ↗
- 3Antimicrobial Resistance Collaborators. 2022. Global burden of bacterial antimicrobial resistance in 2019: a systematic analysis. Lancet 399:629–655.35065702 10.1016/S 0140-6736(21)02724-0PMC 8841637 · doi ↗ · pubmed ↗
- 4GBD Antimicrobial Resistance Collaborators. 2022. Global mortality associated with 33 bacterial pathogens in 2019: a systematic analysis for the Global Burden of Disease Study 2019. Lancet 400:2221–2248.36423648 10.1016/S 0140-6736(22)02185-7PMC 9763654 · doi ↗ · pubmed ↗
- 5Raymond J, Aujard Y, European Study Group. 2000. Nosocomial infections in pediatric patients: a European, multicenter prospective study. Infect Control Hosp Epidemiol 21:260–263. doi:10.1086/50175510782588 · doi ↗ · pubmed ↗
- 6Dessì A, Puddu M, Testa M, Marcialis MA, Pintus MC, Fanos V. 2009. Serratia marcescens infections and outbreaks in neonatal intensive care units. J Chemother 21:493–499. doi:10.1179/joc.2009.21.5.49319933039 · doi ↗ · pubmed ↗
- 7Johnson A, Watson D, Dreyfus J, Heaton P, Lampland A, Spaulding AB. 2020. Epidemiology of Serratia bloodstream infections among hospitalized children in the United States, 2009-2016. Pediatr Infect Dis J 39:e 71–e 73. doi:10.1097/INF.000000000000261832091494 · doi ↗ · pubmed ↗
- 8Mahlen SD. 2011. Serratia infections: from military experiments to current practice. Clin Microbiol Rev 24:755–791. doi:10.1128/CMR.00017-1121976608 PMC 3194826 · doi ↗ · pubmed ↗