Genomic data uncover complex hybridization and evolutionary history of the bunchberry species complex (Cornus L., Cornaceae)
Yanxia Sun, Wenbin Zhou, Qiu-Yun (Jenny) Xiang

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
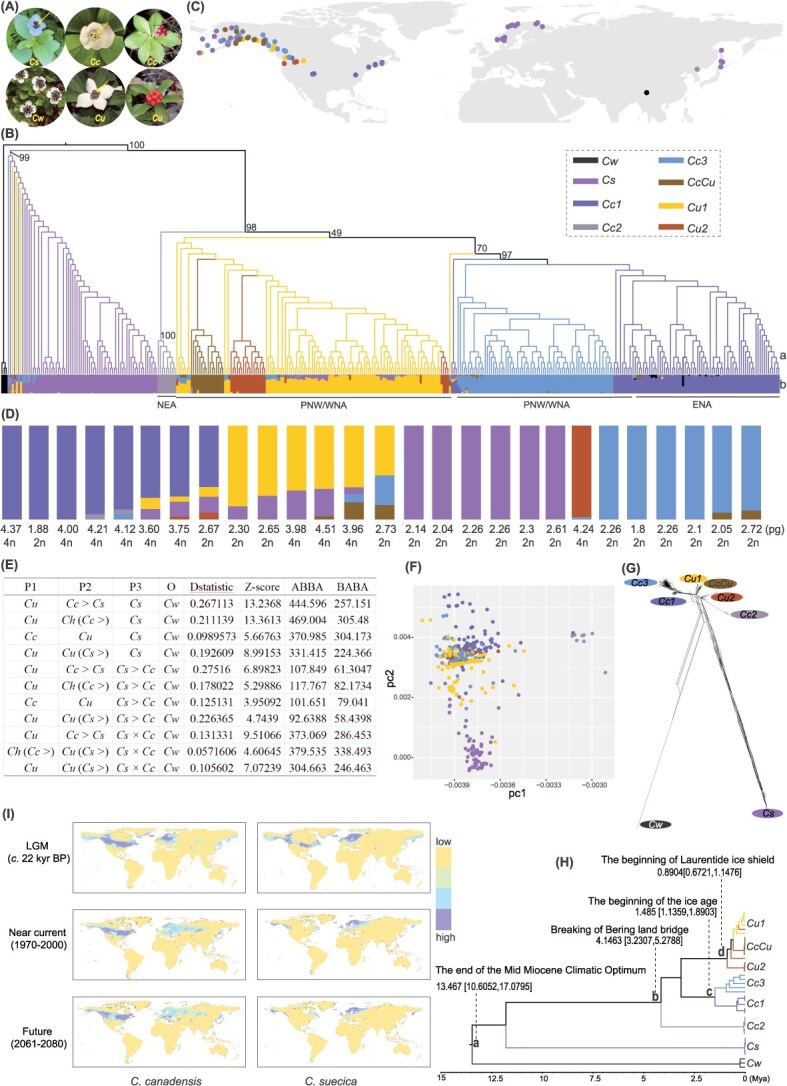 Figure 1
Figure 1- —National Natural Science Foundation of China10.13039/501100001809
- —National Institute of Food and Agriculture10.13039/100005825
- —NSF10.13039/100000001
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBerry genetics and cultivation research · Plant Pathogens and Fungal Diseases · Yeasts and Rust Fungi Studies
Dear Editor,
Hybridization is an important force in plant evolution and speciation, and a powerful traditional method for target breeding of horticultural crops [1]. Investigating the occurrence, extent, and genetic consequences of natural hybridization in a group is not only critical to understanding the processes shaping plant diversity but also important to genetic breeding of ornamental plants by guiding the selection of wild relatives for hybridization. However, our knowledge on the dynamics of natural hybridization and its evolutionary outcomes in many plants, especially those restricted to special habitats and with horticultural importance/potentials, is highly limited.
The dwarf dogwoods, the Arctocrania clade of Cornus L. (Cornaceae), are four rhizomatous perennial species, Cornus canadensis L. (Cc), C. suecica L. (Cs), C. unalaschkensis Ledeb. (Cu), and C. wardiana Rushforth & Wahlsteen (Cw) (Fig. 1A), well known for their strong tolerance to low temperature and uses as ornamental and food plants. These plants are durable, attractive ground covers widely grown in hanging baskets or gardens, and make edible red fruits that are used for jams, jellies and fruit preserves. The boundaries of the four known species are blurred due to prevalent intermediate forms between Cc and Cs throughout the North American boreal–arctic distribution range of the clade, which are classified as Cu, Cc > Cs (morphologically more similar to Cc), and Cs > Cc (morphologically more similar to Cs) (Fig. S1) [2]. These three intermediate forms and the two morphological extremes represented by Cc and Cs comprise the bunchberry species complex (Table S1). The morphological variation suggested that hybridization may have been an important process driving the evolution of the group, leading to species formation and diversification and, maybe, also breakdown of species boundaries. To test the hypothesis, we conducted a study of the species complex by including 374 dwarf dogwood individuals from 88 populations across the distributional range covering morphological variation representing the known species and intermediate forms using genome-wide genetic polymorphisms at 43,137 variant sites from 1,126 ddRAD (double-digest restriction site-associated DNA) loci (Table S2). We integrated population genomics, phylogeography, and distribution modeling (Fig. S2; Tables S3 and S4) to document hybridization and gain insights into its role in shaping the evolution of the species complex and how climate changes have affected hybridization.
Our analyses revealed eight ancestries for the 374 individuals: Cw, Cs, Cc1, Cc2, Cc3, CcCu, Cu1, and Cu2 (Figs 1B, S3, S4, and S5; Table S2), each containing a portion of individuals with admixed ancestries, except Cc2 from northeastern Asia (NEA) (Figs 1B, 1C, S4, and S5). The spatial pattern of genetic clusters and their phylogenetic relationships indicates a number of points: (i) hybridization and introgression within the species complex are prevalent and complicated (Figs 1B, S4, and S5), which was also supported by the D-statistic (Fig. 1E); (ii) Cw, Cs, and Cc2 are geographically and genetically well separated (Figs. 1B, 1C); (iii) Cc and Cu are genetically and evolutionarily more diverse than previously known, showing divergence associated with geographic regions (Fig. 1B and C); (iv) Cc > Cs and Cs > Cc are not recognized as distinct genetic clusters but spread in other clusters (Fig. S4; Table S2); (v) the Pacific Northwest (PNW) region is a center of genetic diversity and heterogeneous hybridization (Figs 1B and S4); (vi) possible ancient hybridization between Cw and Cc1 based on individuals showing significant admixture of ancestries of Cw and Cc1 (Figs 1B, S4, and S5). In short, the patterns of genetic admixture combined with results from the D-statistic suggest the presence of hybrids at different generations, from F1 to later backcross generations, from both ancient and more recent events in the clade, based on levels of introgression/admixture of genetic ancestries of different species.
Although Cu has been recognized as an allopolyploid species from hybridization between Cs and Cc [2], the genetic admixture and phylogeny suggest Cu was probably derived independently of Cs, but evolving within Cc during the Pleistocene (Fig. 1B and H). This timing of Cu’s origin is consistent with the previous hypothesis [2]. Limited data for flow cytometry showed a mixture of ploidy of the individuals in Cu1 including those with admixed ancestries (Fig. 1D). The detection of the diploid Cs × Cu1 genome points to a diploid ancestry of Cu1 and suggests likely autotetraploidy in the Cu1 group (Fig. 1D). Additional flow cytometry data from plants not included in the genetic analysis show a complex variation of genome size at a given location (Fig. S6; Table S5), suggesting dynamic changes in genome sizes that likely involved hybridization, aneuploidy, and/or polyploidy within and between taxa.
Divergence time estimation of the eight genetic groups shows a good match to several past climatic events (Fig. 1H), suggesting significant impacts of historical climate changes on the diversification and hybridization of the dwarf dogwoods. The successive early divergence of Cw and Cs was concordant with the ending of the Mid-Miocene Climatic Optimum. Given the detection of putative hybrids between Cw and Cc1 in eastern North America (ENA) (Fig. S4), the divergence of Cw from Cs + Cc-Cu likely occurred in the north, where their ranges were once overlapping to facilitate hybridization, before it dispersed southward into the Sino-Himalayan region. The divergence between NEA Cc2 and North America (NA) Cc-Cu corresponds to the breaking of the Bering land bridge [3], and the diversification of the NA lineage into the ENA Cc1 and western North America (WNA) Cc3, Cu1, Cu2, and CcCu occurred during the mid-Pleistocene ice age when the grassland in central NA had well-developed isolating forests in the eastern and western USA and glaciation cycles significantly impacted the PNW region, where it is known to have provided refugia within refugia for organisms [4]. Hence, diversification of the CcCu-Cu subclade following the beginning of the Laurentide ice shield was likely promoted by combined effects of geographic isolation, remixing, hybridization, and polyploidization in refugia of the PNW during the glaciation periods.
Considering genetic differentiation, phylogenies, morphology, and distribution, we propose to recognize NEA Cc2 as a new species with the name Cornus orientalis Y.X.Sun, W.B.Zhou & Q.-Y.Xiang, and two cryptic subspecies within Cu [C. unalaschkensis subsp. unalaschkensis (Cu1 + CcCu) and C. unalaschkensis subsp. borealis (Cu2)], and two cryptic subspecies within Cc [C. canadensis subsp. canadensis (Cc1) and C. canadensis subsp. pristina (Cc3)]. Incomplete lineage sorting and gene flow are expected between species and subspecies, which explains why CcCu and Cu2 individuals nested within Cu1.
It must be noted that the previously diverged lineages can face the risk of reverting to a single gene pool due to continuous hybridization and introgression in the PNW region (Fig. 1F). This concern is coupled with the predicted reduction of suitable habitats of Cs in the region (Fig. 1I). The modern suitable range of the species has significantly narrowed and shifted southward in Beringia, Alaska, western Canada, and NE America, compared with the last glacial maximum (LGM) (Fig. 1I). The future range of Cs is predicted to be highly reduced in the PNW, with loss in some parts of ENA. Shrinkage of the coastal arctic and alpine habitats due to sea level increase under climate warming will enhance the threat to Cs from hybridization with Cc and Cu.
In summary, our study based on genome-wide polymorphisms revealed hidden genetic structures of the bunchberry species complex. The evolutionary histories may have involved complicated hybridization, introgression, and polyploidization. Results from the present study support important roles of hybridization and climate changes in the diversification of species adapted to cold and subarctic regions that are facing the most threat of climatic warming [5]. The new knowledge learned from the study that all species are capable of hybridization presents opportunities to breed varieties with desired traits from different species or hybrid forms, such as Cw-like evergreen plants with white petals via artificial hybridization.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Cui Y, Li R, Li G. et al. Hybrid breeding of rice via genomic selection. Plant Biotechnol J. 2020;18:57–6731124256 10.1111/pbi.13170 PMC 6920338 · doi ↗ · pubmed ↗
- 2Murrell Z . Dwarf dogwoods: intermediacy and the morphological landscape. Syst Bot. 1994;19:539–56
- 3Gladenkov AY, Oleinik AE, Marincovich L. et al. A refined age for the earliest opening of Bering Strait. Palaeogeogr Palaeoclimatol Palaeoecol. 2002;183:321–8
- 4Smith ML, Wallace J, Tank DC. et al. The role of multiple Pleistocene refugia in promoting diversification in the Pacific Northwest. Mol Ecol. 2022;31:4402–1635780485 10.1111/mec.16595 · doi ↗ · pubmed ↗
- 5Rantanen M, Karpechko AY, Lipponen A. et al. The Arctic has warmed nearly four times faster than the globe since 1979. Commun Earth Environ. 2022;3:168
- 6Du ZY, Jenny Xiang QY, Cheng J. et al. An updated phylogeny, biogeography, and Phylo Code-based classification of Cornaceae based on three sets of genomic data. Am J Bot. 2023;110:e 1611636480351 10.1002/ajb 2.16116 · doi ↗ · pubmed ↗
- 7Shearer K, Ranney TG. Ploidy levels and relative genome sizes of species, hybrids, and cultivars of dogwood (Cornus spp.). Hort Science. 2013;48:825–30