Development of a Mechanism of Action-Reflective Cell-Based Reporter Gene Assay for Measuring Bioactivities of Therapeutic Glucagon-like Peptide-2 Analogues
Xiaoming Zhang, Chunyan Li, Zhe Deng, Chenggang Liang, Jing Li

TL;DR
Researchers developed a cell-based test to measure the effectiveness of GLP-2 analogues, which are promising treatments for intestinal and bone diseases.
Contribution
A new cell-based reporter gene assay was created to accurately assess the bioactivity of GLP-2 analogues.
Findings
The assay showed good accuracy, linearity, precision, and specificity for measuring GLP-2 analogue potency.
RNA sequencing revealed that GLP-2 analogues have multi-target regulatory effects.
The assay can be used for stability testing, drug screening, and therapeutic monitoring of GLP-2 analogues.
Abstract
Glucagon-like peptide-2 (GLP-2) is a gut hormone that plays a pivotal role in regulating intestinal epithelial cell growth and function, making it a promising therapeutic agent for intestinal damage and bone-related diseases. Nonetheless, the therapeutic potential of GLP-2 is substantially diminished due to its inactivation by dipeptidyl peptidase 4 (DPP-4). In recent years, advancements have been made in developing dipeptidyl peptidase 4 (DPP-4) resistant GLP-2 analogues with an extended half-life. The murine model with extensive experimental bowel resection maintained on parenteral nutrition has been used for assessing the physiology and pharmacology of GLP-2, and for the preclinical validation of GLP-2 analogues. However, it possesses certain limitations, such as complex procedure, considerable variability, and time-consuming nature. Consequently, there is a pressing need for the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
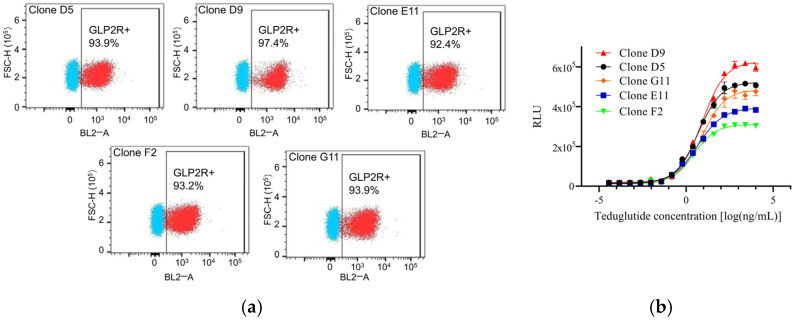 Figure 1
Figure 1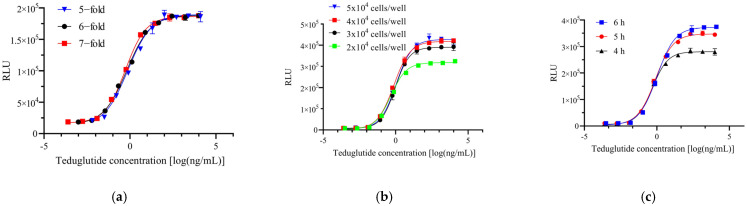 Figure 2
Figure 2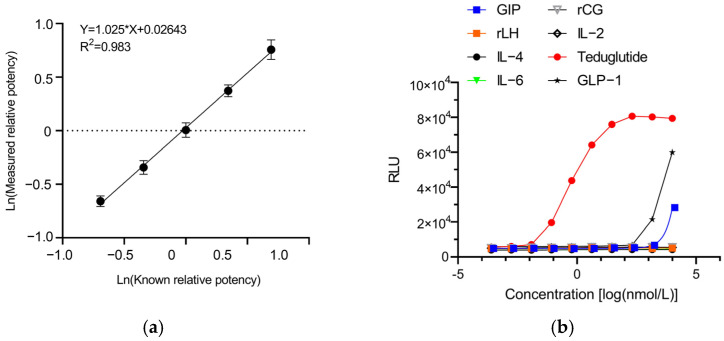 Figure 3
Figure 3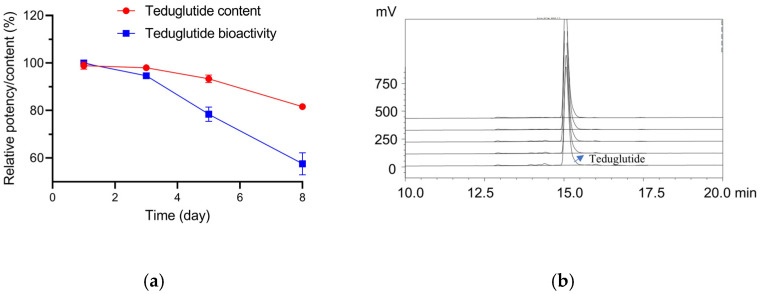 Figure 4
Figure 4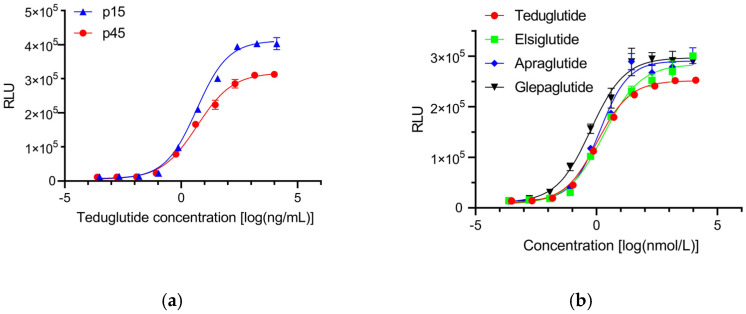 Figure 5
Figure 5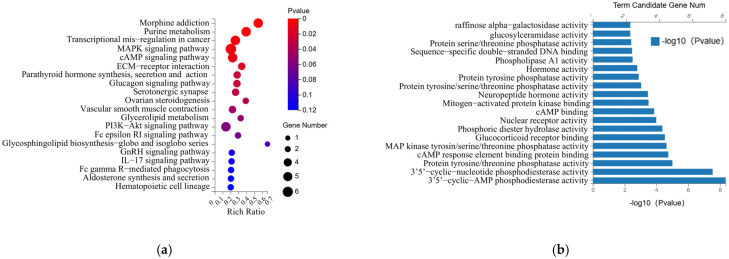 Figure 6
Figure 6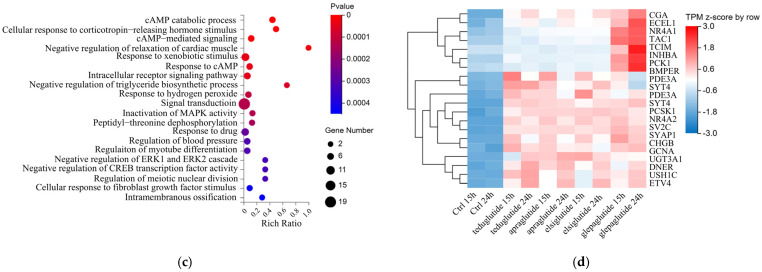 Figure 7
Figure 7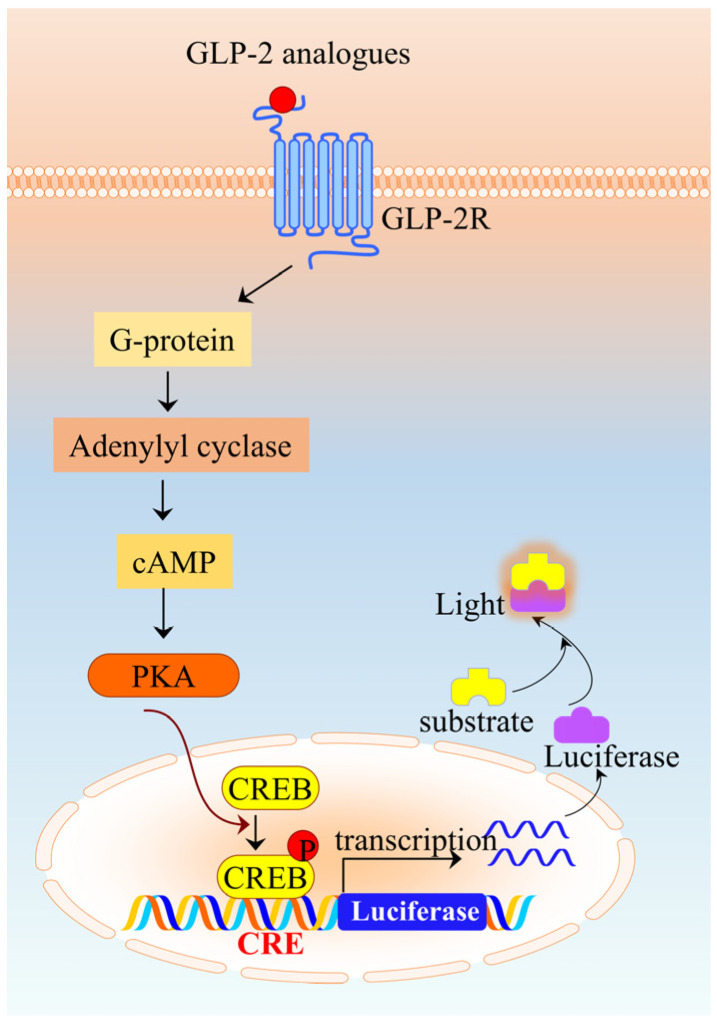 Figure 8
Figure 8- —State Key Laboratory of Drug Regulatory Science
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsDiabetes Treatment and Management · Peptidase Inhibition and Analysis · Pharmaceutical studies and practices
1. Introduction
Glucagon-like peptide-2 (GLP-2) is a 33 amino acid-long intestinal hormone that is encoded together with glucagon and glucagon-like peptide-1 (GLP-1), by the proglucagon gene. As a gastrointestinal hormone, GLP-2 is secreted postprandially by enteroendocrine L cells located throughout the intestinal tract [1]. Additionally, albeit in significantly smaller quantities, GLP-2 is also secreted from preproglucagonergic neurons situated in the brainstem and the hypothalamus [2]. GLP-2 is a pleiotropic hormone affecting multiple facets of intestinal physiology. Primarily, it regulates intestinal epithelial cell growth and functions that are crucial for digestion and absorption of nutrients, including enhancement of nutrient absorption, stimulation of blood flow, anti-inflammatory activity, improvement of barrier function, and modulation of motility [3,4]. Thus, GLP-2 emerges as a promising therapeutic target in intestinal damage. It was found that GLP-2 also plays an important role in bone turnover, thereby possessing potential for the treatment of bone-related diseases.
GLP-2 exerts its effects largely through interaction with the GLP-2 receptor (GLP-2R), which belongs to the class B1 of the G protein-coupled receptors. However, it is noteworthy that GLP-2R has been localized to central nervous system and several intestinal cell types that do not include the proliferating crypt cells [5]. This led to the hypothesis that GLP-2 acts indirectly through a complex network of indirect mediators that induce diverse signaling pathways, which remains poorly understood. For instance, GLP-2 has been shown to enhance the weight of small and large intestinal weight by stimulating epithelial cell proliferation and inhibition of apoptosis. It has been demonstrated that GLP-2 acts through an IGF-I-dependent pathway to activate intestinal crypt β-catenin, which integrates signals from multiple pathways to regulate the balance between crypt cell proliferation and differentiation [6]. The antiapoptotic response action of GLP-2 has been linked to the cAMP pathway involving both protein kinase A (PKA)-dependent and -independent mechanisms [7]. BHK fibroblasts stably transfected with the rat GLP-2R exhibit dose-dependent cAMP accumulation in response to GLP-2 administration [8,9].
The native form of GLP-2 undergoes rapid inactivation by the enzyme dipeptidyl peptidase 4 (DPP-4), which excises the two N-terminal amino acids, resulting in the truncated and inactive GLP-2 (3–33). This truncated form is subsequently degraded by endopeptidases and presumably cleared via renal excretion [10]. The inactivation of GLP-2 by DPP-4 significantly reduces its potential as a therapeutic agent. Consequently, DPP-4 resistant GLP-2 analogues with an extended half-life were developed [11,12]. Teduglutide (Gattex, Revestive) is the first and the only GLP-2 analogue that has been approved for medical treatment of short bowel syndrome (SBS). Teduglutide has an Ala-to-Gly substitution at position 2 to eliminate the DPP4 cleavage site ([Gly2] hGLP-2 (1–33)), and exhibits a terminal half-life of 2 to 3 h when administered subcutaneously to humans [13]. SBS is a severe and chronic disease characterized by malabsorption due to physical or functional loss of significant portions of the small intestine [14]. Managing SBS is challenging and requires careful monitoring to ensure nutritional and fluid needs are met through parenteral nutrition (PN). However, long-term PN can lead to serious complications. It has been found that native GLP-2 and teduglutide increase intestinal absorption by promoting intestinal growth and delays accelerated gastric emptying, gastrointestinal transit, and hypersecretions in SBS [15,16]. In patients with SBS, daily treatment with teduglutide led to a significant reduction in PN requirements and an improvement in intestinal function [14,17].
In addition to teduglutide, various additional analogues of GLP-2 with reduced clearance relative to GLP-2 are under development for SBS, including apraglutide, glepaglutide, and elsiglutide [18]. In a head-to-head preclinical study, apraglutide demonstrated a longer half-life and greater intestinotrophic effect in rat, compared to teduglutide and glepaglutide [2]. In Phase II clinical trials for patients with short bowel syndrome (SBS), apraglutide demonstrated a significant reduction in patients’ dependence on PN support, indicating its broad application prospects as a potential new therapy for rare gastrointestinal diseases [19,20]. Glepaglutide incorporates a number of design strategies presumably to optimize pharmacokinetic and potency properties, and is primarily used in research for inflammatory bowel disease and Crohn’s disease. Glepaglutide is also being developed as a potential treatment option for SBS [21]. Elsiglutide can increase cell proliferation and reduce intestinal cell apoptosis, showing great potential in the treatment of gastrointestinal mucositis, including chemotherapy-induced diarrhea [22].
While great excitement has been engendered by the clinical success of GLP-2 analogues, the development of appropriate in vitro bioassays for measuring potency and characterizing the biological properties of GLP-2 analogues has not been reported. The murine SBS model with extensive experimental bowel resection maintained on PN has been used for assessing the physiology and pharmacology of GLP-2, and for the preclinical validation of GLP-2 analogues [23,24,25]. However, the SBS model is not suitable for bioassay development, since it has certain limitations, such as complicated procedures, a time-consuming nature, high variability, and the sacrifice of the experimental animals after treatment. The reporter gene assay (RGA)—which is mechanism-of-action (MOA)-related, less variable, accurate, and precise—has been adopted as alternative assay for the bioactivity determination of biopharmaceuticals in quality control (QC) in recent years [26,27,28]. Here, we develop a novel RGA that engages GLP-2R and the subsequent signaling pathway. This cell-based bioassay is suitable for lot release and stability testing, as well as for biological property characterization of the GLP-2 analogues.
2. Results
2.1. Generation of the HEK293-GLP2R-Luc Cell Line
The intracellular effects of GLP-2 are initiated by binding to its cell membrane-spanning receptor, the GLP-2R. Following GLP-2 binding, the intracellular cAMP biosynthesis is elevated, leading to activation of PKA, followed by phosphorylation of the cAMP-response element binding protein (CREB), which then induces the transcription of several GLP-2 target genes. In order to develop a novel RGA for the determination of GLP-2 bioactivity, we constructed a HEK293-GLP-2R-Luc cell line based on the GLP-2R/PKA/CREB signaling pathway (Figure A1). This was achieved by stably expressing a luciferase reporter gene and GLP-2R in HEK293 cells. The transcription of the luciferase reporter gene is controlled by a cis-regulatory element (CRE) that can be recognized by CREB and activated by GLP-2. Specifically, we sequentially infected HEK293 cells with lentivirus particles carrying the luciferase gene under the control of CRE and lentivirus particles expressing GLP-2R. Resistant clones were selected in the presence of puromycin and zeocin, and then subcloned by limited dilution. The expression of GLP-2R was then confirmed with flow cytometry analysis. As shown in Figure 1a, five clones (D5, D9, E11, F2, G11) were subjected to cytometry analysis, and all clones showed ectopic expression of GLP-2R. The positive rates of GLP-2R for the monoclonal clones D5/D9/E11/F2/G11 were 93.9%, 97.4%, 92.4%, 93.2%, and 93.9%, respectively. Furthermore, all five clones showed high response to teduglutide treatment, and clone D9 was selected for subsequent experiments since it was more sensitive to GLP-2 (10 µg/mL for 6 h, EC50 of 4.6 ng/mL) and had the highest lower/upper asymptotes (~31.4) compared to the other 4 clones (Figure 1b). This clone was then designated as HEK293-GLP-2R-Luc cell line and employed to establish a RGA for GLP-2 analogues.
2.2. Reporter Gene Assay Optimization
To determine the optimal RGA conditions, we optimized the RGA via the following parameters: dilution ratio, incubation time, and cell density. When one condition was optimized, the other conditions were held constant. To establish the most reliable sigmoidal dose–response curve, we systematically evaluated 5-, 6-, and 7-fold serial dilutions of teduglutide, starting from an initial concentration of 10 µg/mL. To determine the proper cell density, we investigated 4 different cell densities: 2 × 10^4^, 3 × 10^4^, 4 × 10^4^, and 5 × 10^4^ cells per well of 96-well white plate. Finally, 3 different incubation times (4, 5, and 6 h) were investigated. As shown in Figure 2a, the point distribution of the four-parameter logistic (4PL) curves obtained were all complete, but the 4PL curve obtained from a 7-fold dilution is the most uniform. An increased drug response was seen when the cell number increased from 2 × 10^4^ cells per well to 4 × 10^4^ cells per well, but no further increase in drug response was seen when the number of target cells was at 4 × 10^4^ cells/well (Figure 2b). A concentration-dependent drug response was seen after 4 h of incubation. A stronger response (higher upper asymptote) was observed when the incubation time was increased to 5 h and 6 h (Figure 2c). Thus, the optimized conditions for the subsequent experiments were as follows: (1) 7-fold dilution of teduglutide for 10 points; (2) 4 × 10^4^ cells per well for the optimal cell density; (3) the optimal incubation time was 5 to 6 h. The optimized experimental parameters were systematically applied to all GLP-2 analogs evaluated in this study, ensuring uniformity across all experimental cohorts.
2.3. Validation of the Reporter Gene Assay
In order to prove the relative accuracy, linearity, precision, and specificity of the RGA, we comprehensively validated the RGA according to the ICH Q2 (R2) guidelines and the Chinese Pharmacopoeia (ChP), as well as the United States Pharmacopoeia (USP).
Table 1 showed the average and 90% confidence interval (CI) of validation results in the Ln scale, as well as corresponding potency, relative bias, and the geometric coefficient of variation (%GCV). The RGA has acceptable relative bias at levels from 50% to 200%, yielding mean relative bias region of −0.034 to 6.60, which fall within the acceptance region of ±12% relative bias. The slope of the linear curve was 1.025, which falls within the acceptance region of 0.8 to 1.25 (Figure 3a). The relative bias and slope of the linear curve indicated good relative accuracy. The mean %GCV of five relative potency levels were between 4.97% and 9.46%, which were below 20%, indicating high intermediate precision of the novel RGA. Furthermore, the linear curve of Ln measured potency versus Ln target potency also showed excellent linearity with R^2^ of 0.983. Taken together, these results demonstrated the novel RGA had acceptable relative bias, intermediate precision, and linearity within the range of 50% to 200%.
Specificity describes the ability of this RGA to assess teduglutide exclusively. GLP-2R, glucose-dependent insulinotropic peptide receptor (GIPR), and GLP-1R all belong to the class B GPCR subfamily, which is mainly expressed in the gut, pancreas, and brain [29]. We thus investigated the responsiveness of HEK293-GLP-2R-Luc cells to GIP and GLP-1. As shown in Figure 3b, the concentration–response curve was unsuccessfully induced by GIP and GLP-1 that belongs to the same family as GLP-2. Furthermore, we randomly examined the response of HEK293-GLP-2R-Luc cells to two more hormones, rCG and rLH, the receptor of which is also a G-protein-coupled receptor [30]. Several cytokines, including IL-2, IL-4, and IL-6, were also used to validate the specificity of the RGA. As expected, no dose–dependent response curve was induced by rCG, rLH, IL-2, IL-4, and IL-6. Collectively, these results indicated that the established RGA is highly specific to teduglutide.
2.4. Stability Testing of Teduglutide with the RGA
We expected that the new RGA could be applied to the biological stability study of teduglutide. According to ICH Q5C “Stability Testing of Biotechnological/Biological Products” and ChP General Rule 9402 “Stability Testing of Biological Products”, the stability test should explore and optimize the test conditions based on the product’s own characteristics. The temperature in the influence factor test should be high enough to observe the inactivation, denaturation, or degradation of the sample and exceed the quality standard limit. To this end, teduglutide drug substance (DS) was subjected to a temperature of 37 °C for durations of 1, 3, 5, 8, 12, and 14 days, respectively. To our surprise, the DS solution develops a clear colloidal precipitate after incubation for 12 or 14 days at 37 °C, rendering it unsuitable for bioactivity determination. As illustrated in Figure 4a, The relative potency of teduglutide, when compared to non-treated disease state (DS), progressively diminishes with increasing incubation time at 37 °C, as evidenced by a decline in EC50 values from 100.5% to 57.5%. The teduglutide contents were then determined using high-performance liquid chromatography (HPLC). As expected, the content of teduglutide decreased from 99.9% to 81.6% (Figure 4b), which suggested that the reduction of teduglutide bioactivity was successfully monitored via our new RGA.
2.5. Stability and Application of the New RGA
The cell passage stability serves as an indicator of system stability, and plays a pivotal role in assessing the consistency of bioactivity determination. As thus, two different passages (p15 and p45) of HEK293-GLP-2R-Luc cells after cryopreservation and recovery were applied to measure the relative bioactivities of teduglutide reference sample. As shown in Figure 5a, two parallel teduglutide dose-dependent curves were obtained, with the lower/upper asymptotes decreased slightly from 32 in p15 cells to 28 in p45 cells, while the EC50 of both cells were similar (4.6 ng/mL in p15 cells, 4.8 ng/mL in p45 cells). We then measured the bioactivity of 3 batches of teduglutide DS and 3 batches of drug products (DP) using the two passage cells simultaneously. There were no statistical differences in the relative bioactivity of teduglutide between the two passages. Collectively, these results suggest that our HEK293-GLP-2R-Luc cells exhibited sufficient stability.
In order to explore the application of the new RGA in other GLP-2 analogues, apraglutide, glepaglutide, and elsiglutide were synthesized and applied to bioactivity investigation using the HEK293-GLP-2R-Luc cells (Table 2). All three GLP-2 analogues were diluted to 2.665 µmol/L, and further diluted 7-fold for 10 concentration points. As expected, all three analogues induced sigmoid curves similar to those induced by teduglutide (Figure 5b). These findings indicated that the bioactivity of GLP-2 analogues was successfully monitored via our new RGA.
2.6. The Multi-Target Regulatory Effect of GLP-2 Analogues Was Revealed by RNA Sequencing
GLP-2 regulates proliferative and cytoprotective pathways in the intestine. Administration of GLP-2 to rodents enhances the growth of the intestinal epithelium by stimulating the proliferation of crypt cells and inhibiting the apoptosis of enterocytes [8,31]. A study on GLP-2 biology indicates that the activation of GLP-2R triggers a cAMP- and PKA-dependent pathway, which is associated with the inhibition of apoptosis. Additionally, it couples with the activation of the Ras/MAPK pathway and stimulates DNA synthesis [32]. GLP-2 has also been reported to stimulate neuronal differentiation of enteric neurons and glial cells in culture via the PI3K/Akt/mTOR pathway [33].
RNA sequencing was applied to elucidate whether GLP-2 analogues also activate proliferative and cytoprotective pathways in HEK293-GLP-2R-Luc cells. A total of 83 differentially expressed genes (DEGs) comprised of 76 upregulated genes and 7 downregulated genes with more than 2-fold change were obtained from teduglutide, apraglutide, glepaglutide, and elsiglutide-treated HEK293-GLP-2R-Luc cells in comparison to DMSO control-treated cells (Appendix A, Table A1). Notably, 370 DEGs, comprising 283 upregulated genes and 87 downregulated genes with a fold change greater than 2 are differentially expressed exclusively in HEK293-GLP-2R-Luc cells treated with glepaglutide (Table A2). The KEGG enrichment analysis of the DEGs listed in Table A1 revealed that these DEGs were predominantly enriched in pathways such as the MAPK signaling pathway, cAMP signaling pathway, glucagon signaling pathway, and PI3K-Akt signaling pathway (Figure 6a). GO enrichment of molecular function (MF) revealed enrichment in cAMP phosphodiesterase activity, protein tyrosine/threonine phosphatase activity, MAP kinase tyrosine/serine/threonine phosphatase activity, mitogen-activated protein kinase binding (Figure 6b). GO enrichment of biological process (BP) involved a wide range of physiological activities, including cAMP catabolic process, response to xenobiotic stimulus, response to hydrogen peroxide, regulation of blood pressure, negative regulation of ERK1 and ERK2 cascade, and cellular response to fibroblast growth factor stimulus (Figure 6c). The top 20 significantly upregulated genes in teduglutide, apraglutide, glepaglutide, and elsiglutide-treated HEK293-GLP-2R-Luc cells were listed in the heatmap (Figure 6d).
3. Discussion
The diverse functions of GLP-2 highlight its significance in maintaining intestinal health and physiology. As a naturally occurring gut hormone, GLP-2 not only promotes nutrient absorption by decreasing intestinal motility and permeability, but also plays a pivotal role in stimulating intestinal epithelial cell growth and function [34]. This multifaceted action of GLP-2 is crucial for the maintenance of the intestinal barrier, which is essential for preventing the translocation of harmful bacteria and toxins into the bloodstream [35]. The development of GLP-2 analogues, such as teduglutide, apraglutide, glepaglutide, and elsiglutide, has further expanded the therapeutic potential of GLP-2. These analogues exhibit extended half-lives and enhanced bioactivities, making them suitable for the treatment of various intestinal disorders [36].
In this study, we have established a robust and reproducible cell-based bioassay to measure the potency and characterize the biological activities of GLP-2 analogues. The RGA utilizes HEK293 cells stably expressing the GLP-2 receptor and a luciferase reporter gene, creating a sensitive and specific system for detecting GLP-2-mediated signaling. The RGA has been meticulously validated according to international guidelines, demonstrating its accuracy, linearity, precision, and specificity. These validation studies have confirmed that the new RGA is capable of measuring the bioactivity of teduglutide, as well as other GLP-2 analogues. Moreover, the new RGA can detect heat-stressed teduglutide sensitively, indicating its potential application in stability testing of GLP-2 analogues. The passage stability of the HEK293-GLP-2R-Luc cells was also assessed, ensuring the consistency and reliability of the bioactivity determinations over time.
RNA sequencing analysis identified a total of 83 differentially expressed genes in GLP-2 analogue-treated cells, providing valuable insights into the potential mechanisms of action of GLP-2 analogues. The upregulated genes are involved in a wide range of physiological activities, including cAMP catabolic process, response to xenobiotic stimulus, response to hydrogen peroxide, gluconeogenesis and lipogenesis regulation, and muscle growth and differentiation, which is consistent with previous reports [6,31,37]. This suggested that the HEK293-GLP-2R-Luc cells may potentially be useful as a cellular model for analysis of GLP-2R signaling and the screening of potential therapeutic drugs targeting to GLP-2R.
The GO enrichment analysis of the biological processes associated with these DEGs further supports the pleiotropic effects of GLP-2 analogues. For instance, the upregulation of genes involved in cAMP catabolic process and response to hydrogen peroxide indicates that GLP-2 analogues may have antioxidant and cytoprotective effects [38]. Similarly, the modulation of genes related to gluconeogenesis and lipogenesis regulation suggests that GLP-2 analogues may influence metabolic pathways and energy homeostasis [5,37]. It is worth noting that this study also found that GLP-2 analogues significantly regulated the expression of multiple genes, including NDRG1, NR4A2, etc. NDRG1 has been involved in cell growth and differentiation [39]. NR4A2 is important for the differentiation and maintenance of meso-diencephalic dopaminergic neurons during development [40]. Transcriptome data have revealed a rich GLP-2-related regulatory network, which provides a molecular-level explanation for the clinical observation that GLP-2 analogues improve intestinal barrier function. However, future research still needs to integrate multi-omics methods and establish intestinal-specific cell GLP-2 receptor knockout models, intestinal organoids, etc., to precisely dissect the molecular mechanisms of these effects. In addition, considering the potential application value of GLP-2 analogues in metabolic diseases such as obesity and diabetes, in-depth exploration of their role in energy metabolism will have significant translational medical significance.
It is of note that HEK293-GLP2R-Luc cells treated with glepaglutide exhibited a significantly higher number compared to the other three GLP-2 analogues-treated cells. This suggested that glepaglutide may have a distinct effect on gene expression within these cells. Additionally, the unique gene expression profile induced by glepaglutide may provide insights into its potential for treating specific gastrointestinal disorders or conditions where modulation of these pathways could be beneficial. Additional studies are needed to further elucidate the roles of these DEGs and their interactions in the context of glepaglutide-mediated cellular responses.
In conclusion, the establishment of this MOA-reflective cell-based bioassay represents a significant advancement in the evaluation of GLP-2 analogues. The new RGA is sensitive, specific, and reproducible, making it suitable for quality control, lot release, and stability testing, as well as for biological property characterization of the GLP-2 analogues. Furthermore, the RNA sequencing analysis has provided valuable insights into the potential mechanisms of action of GLP-2 analogues, paving the way for future research in this area. Overall, this study contributes to the ongoing efforts to develop novel therapies for intestinal damage and other related disorders.
4. Materials and Methods
4.1. Materials
The human embryonic kidney 293 (HEK293) cell line was purchased from ATCC (Manassas, VA, USA). HEK293-GLP-2R-Luc cells were cultured in Dulbecco’s modified eagle medium (DMEM) supplemented with 10% fetal bovine serum (FBS), 200 µg/mL zeocin, and 3 µg/mL puromycin. Both antibiotics, recombinant human Interleukin-2 (IL-2, PHC0027), IL-4 (200-04), IL-6 (200-06), and mouse IgG isotype control were purchased from Thermo Fisher Scientific (Waltham, MA, USA). Recombinant human chorionic gonadotrophin (rCG, 410021) was supplied by NIFDC (Beijing, China). Recombinant human luteinizing hormone (rLH) and teduglutide were provided by different manufacturers and stored at 4 °C or −80 °C in our laboratory. DMEM and FBS were purchased from Gibco (Grand Island, NY, USA). Britelite plus reporter Gene Assay System (6066769) was purchased from Revvity (Waltham, MA, USA). The pLU.cmv.Zeo and pLU.CMV.Luc.PGK.PURO were obtained from dlbiotech (Suzhou, China). The mouse anti-human GLP-2R monoclonal antibody (Clone # 413801) was purchased from R&D (Minneapolis, MN, USA).
4.2. Peptide Synthesis
GIP, GLP-1, glepaglutide, elsiglutide, and apraglutide peptides used in this study were synthesized by GenScript (Nanjing, China). All these peptides were prepared as trifluoroacetic acid salts by solid-phase peptide synthesis, subsequently purified by reverse-phase HPLC, and definitively verified via mass spectrometry. All the peptides had purity no less than 99% as measured by analytical HPLC. Amino acid sequences of the GLP-2 analogue peptides used in this study were identified from a published report [2].
4.3. Plasmid Constructions and Virus Production
To generate constructs for viral packaging, the CRE-miniP (TGACGTCAGCTGCCAGATCCCATGGCCGTCATACTGTGACGTCTTTCAGACACCCCATTGACGTCAATGGGAGAACAGATCTGGCCTCGGCGGCCAAGCTTAGACACTAGAGGGTATATAATGGAAGCTCGACTTCCAG) was synthesized and cloned into the pLU.CMV.Luc.PGK.PURO vector via the Cla I and Xba I sites. The full-length cDNA of human GLP-2R was synthesized, cloned, and inserted into the pLU.cmv.Zeo vector using BamH I and Sal I sites. Virus production was conducted as described previously [41]. Briefly, HEK293LT cells cultured in 60 mm dishes were co-transfected with 4 mg of lentiviral transfer constructs (pLU.CRE-miniP.Luc.PGK.PURO or pLU.cmv-GLP2R.Zeo), 2 mg of the packaging construct pCMV-DR8.91 (encoding HIV-1 gag-pol), and 0.4 mg of PMD2. G (encoding the VSV-G envelope) using the calcium phosphate method. Lentivirus-containing supernatants were harvested on two consecutive days and concentrated.
4.4. Generation of the HEK293-GLP-2R-Luc Cell Line
To generate a HEK293 cell line stably expressing CRE-driven luciferase and GLP2R, the CRE-miniP.Luc virus-containing cell supernatants were diluted in DMEM in the presence of 8 µg/mL polybrene before being added to HEK293 cells. To obtain a stable cell pool, the medium was replaced 48 h after incubation, with fresh culture medium containing 3 µg/mL puromycin. After about 3 weeks selection, the cell pool was then further infected with pLU.cmv-GLP2R.Zeo virus and selected with 200 µg/mL Zeocin and 3 µg/mL puromycin. After another 3 weeks selection, stable HEK293-GLP-2R-Luc monoclonal cell lines were obtained by limited dilution. The expression of GLP-2R was examined using flow cytometry. After that, the positive clones were routinely maintained in selective medium.
4.5. Flow Cytometry Analysis
HEK293-GLP-2R-Luc cells were rinsed once with PBS and incubated in assay medium at 37 °C with 5% CO_2_ overnight (1624 h). The cells were then harvested, rinsed twice with cold staining buffer, and incubated with 1.0 μg/mL mouse anti-human GLP-2R monoclonal antibody or mouse IgG isotype control on ice for 3045 min in the dark. After staining, the cells were washed twice with staining buffer and then resuspended in the same buffer for analysis. All of the data were acquired using an Attune NxT Flow Cytometer (Thermo Fisher Scientific, Waltham, MA, USA).
4.6. Reporter Gene Assay Procedure
Cell-based RGA has been described previously [41]. Briefly, 50 microliters of HEK293-GLP-2R-Luc cells were seeded into a 96-well white plate at a density of 4 × 10^4^ cells/well. The cells were then incubated at 37 °C with 5% CO_2_ overnight (16 to 24 h). The next day, the teduglutide was serially diluted 7-fold in culture medium at a starting concentration of 10 µg/mL, corresponding to 2.665 µmol/L. Fifty microliters of diluted teduglutide were then added to each well. Notably, the final concentrations of teduglutide ranged from 5000 ng/mL to 0.000124 ng/mL. After 5 h of incubation, 100 µL of britelite plus reporter Gene Assay reagent were added to each well and mixed thoroughly under subdued light conditions for 5 min. The RLU signal was then determined with a SpectraMax M5e multimode plate reader (Molecular Devices, San Jose, CA, USA).
4.7. Validation of the RGA
The RGA was validated as described previously [41]. Briefly, the teduglutide standard reference was diluted in the culture medium to five starting concentrations, which were 5, 7.1, 10, 14.1, and 20 µg/mL. Another independently prepared 10 µg/mL teduglutide standard reference sample was used as the in-house reference. The expected relative potencies of the teduglutide dilutions were 50%, 71%, 100%, 141%, and 200%. Samples of each potency level were tested by two analysts 8 times in total. To investigate whether HEK293-GLP-2R-Luc cells also respond to GIP, GLP-1, rCG, rLH, IL-2, IL-4, and IL-6, all those samples were diluted to 2.665 µmol/L and further diluted 7-fold for 10 concentration points. The response of HEK293-GLP2R-Luc cells to these samples were determined as illustrated in 4.6.
4.8. Stability Testing of Teduglutide with the RGA
Teduglutide DS was kept at 37 °C for 1, 3, 5, 8,12, and 14 days, while the non-treated DS was preserved at −80 °C. The bioactivities of the treated and non-treated DS were then measured by the RGA. The teduglutide contents were analyzed by HPLC. An XBridge Peptide BEH C18 column (300 A, 5 μm, 4.6 mm × 250 mm) purchased from Waters (Milford, MA, USA) was used with a flow rate of 1.0 mL/min and was detected at 214 nm. The mobile phase was aqueous solution containing 0.1% trifluoroacetic acid and acetonitrile containing 0.1% trifluoroacetic acid. A total of 50 μL DS samples were injected into the column. The relative content of teduglutide was determined by teduglutide peak area ratio of DS subjected to a temperature of 37 °C to the non-treated DS.
4.9. RNA Sequencing (RNA-seq)
For the preparation of total RNA for RNA-seq, HEK293-GLP-2R-Luc cells cultured in 6-well plate were treated with 10 µg/mL teduglutide, apraglutide, glepaglutide, elsiglutide, or DMSO for 15 h or 24 h. Total RNA was extracted with RNeasy mini kit (Qiagen, 74104, Hilden, Germany). A minimum of 2 μg of total RNA obtained from a mixture of 3-wells of cells were used for library construction. Library sequencing was performed on the Illumina HiSeq-PE150 platform (San Diego, CA, USA). RNA-seq raw data has been submitted to NCBI, and the accession numbers will be provided during review.
4.10. Statistical Analysis
For in vitro RGA, a 4PL model was used to fit the dose-response curve, which expresses the relationship between the RLU and log10 of the 10 points of teduglutide concentration. The lower/upper asymptote was determined by the ratio of the top asymptote to the bottom asymptote when RGA best fit the 4PL model. The correlation coefficient (R^2^), deviations from parallelism, and the regression in the 4PL model were estimated using SoftMax^®^ Pro 7.1 software (Molecular Devices, San Jose, CA, USA). Specifically, a linear regression with an R^2^ value no less than 0.98 was considered to have a very good fit. Additionally, a p-value below 0.01 was regarded as statistically significant. The relative potency of a sample was indicated by the EC50 ratio of the teduglutide standard reference to the teduglutide testing sample, resulting from the constraints of the 4PL model. All the dose-response curves were plotted using GraphPad Prism^®^ software 8.0.2 (GraphPad, Boston, MA, USA).
5. Conclusions
In this study, we successfully developed a robust and reproducible RGA utilizing a stable HEK293 cell line expressing the GLP-2 receptor and a luciferase reporter gene. The RGA demonstrated acceptable relative bias, intermediate precision, and linearity within the tested range. Through validation, the specificity and stability of the RGA were confirmed. Additionally, the RGA was capable of monitoring the reduction of teduglutide bioactivity. Our findings provide a sensitive and specific system for detecting GLP-2-mediated biological activities, offering valuable insights into the potential mechanisms of action of GLP-2 analogues.
6. Patents
A Chinese patent titled “A cell for determining the biological activity of GLP-2 analogues” (ZL202411308318.6) has been granted by the China National Intellectual Property Administration based on this research.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Gabe M.B.N. von Voss L. Hunt J.E. Gadgaard S. Gasbjerg L.S. Holst J.J. Biased GLP-2 agonist with strong G protein-coupling but impaired arrestin recruitment and receptor desensitization enhances intestinal growth in mice Br. J. Pharmacol.20231801674168910.1111/bph.1604036683195 · doi ↗ · pubmed ↗
- 2Suzuki R. Brown G.A. Christopher J.A. Scully C.C.G. Congreve M. Recent Developments in Therapeutic Peptides for the Glucagon-like Peptide 1 and 2 Receptors J. Med. Chem.20206390592710.1021/acs.jmedchem.9b 0083531577440 · doi ↗ · pubmed ↗
- 3Hargrove D.M. Alagarsamy S. Croston G. Laporte R. Qi S. Srinivasan K. Pharmacological Characterization of Apraglutide, a Novel Long-Acting Peptidic Glucagon-Like Peptide-2 Agonist, for the Treatment of Short Bowel Syndrome J. Pharmacol. Exp. Ther.202037319320310.1124/jpet.119.26223832075870 · doi ↗ · pubmed ↗
- 4Zhang J. Ning J. Hao X. Han X. Fu W. Gong Y. Glucagon-like peptide-2 protects the gastric mucosa via regulating blood flow and metabolites Front. Endocrinol.202213103655910.3389/fendo.2022.1036559 PMC 980141036589839 · doi ↗ · pubmed ↗
- 5Pálsson T.G. Gilliam-Vigh H. Jensen B.A.H. Jeppesen P.B. Lund A.B. Knop F.K. Targeting the GLP-2 receptor in the management of obesity Peptides 202417717121010.1016/j.peptides.2024.17121038579917 · doi ↗ · pubmed ↗
- 6DubéP.E. Brubaker P.L. Frontiers in glucagon-like peptide-2: Multiple actions, multiple mediators Am. J. Physiol. Endocrinol. Metab.2007293 E 460E 46510.1152/ajpendo.00149.200717652153 · doi ↗ · pubmed ↗
- 7Yusta B. Estall J. Drucker D.J. Glucagon-like peptide-2 receptor activation engages bad and glycogen synthase kinase-3 in a protein kinase A-dependent manner and prevents apoptosis following inhibition of phosphatidylinositol 3-kinase J. Biol. Chem.2002277248962490610.1074/jbc.M 20135820011978789 · doi ↗ · pubmed ↗
- 8Yusta B. Boushey R.P. Drucker D.J. The glucagon-like peptide-2 receptor mediates direct inhibition of cellular apoptosis via a c AMP-dependent protein kinase-independent pathway J. Biol. Chem.2000275353453535210.1074/jbc.M 00551020010940305 · doi ↗ · pubmed ↗