Genome-Wide Analysis of GRETCHEN HAGEN3 Genes and Characterization of IAA-Amido Synthetase Gene CsGH3.1 in Rhizome Proliferation in Cymbidium sinense ‘Qijianbaimo’
Sisi Jia, Cuiling Liu, Chushu Liao, Ruizhen Zeng, Li Xie, Qian Wei, Zhisheng Zhang

TL;DR
This study explores GH3 genes in Cymbidium sinense, focusing on CsGH3.1's role in rhizome growth and auxin regulation.
Contribution
The study identifies and characterizes CsGH3.1 as a key regulator of rhizome proliferation in Cymbidium sinense.
Findings
Seven GH3 genes were identified and classified into two subfamilies in Cymbidium sinense.
CsGH3.1 and CsGH3.2 are upregulated by NAA, suggesting roles in auxin homeostasis during rhizome proliferation.
Overexpression of CsGH3.1 in Arabidopsis reduced growth and callus proliferation, confirming its regulatory role.
Abstract
This study systematically identified and functionally analyzed the GRETCHEN HAGEN 3 (GH3) gene family in Cymbidium sinense ‘Qijianbaimo’ (CSQ). Seven GH3 genes (CsGH3.1–CsGH3.7) were identified using genomic and transcriptomic data. Phylogenetic analysis revealed that CsGH3s are divided into two subfamilies, with CsGH3.1–CsGH3.4 belonging to subfamily II and CsGH3.5–CsGH3.7 to subfamily I. Gene structure analysis showed that CsGH3s contain one to four exons, and their promoter regions feature auxin and jasmonic acid response elements. CsGH3.1 and CsGH3.2 were significantly upregulated in response to naphthaleneacetic acid (NAA) during rhizome proliferation, indicating their pivotal role in auxin homeostasis regulation. The overexpression of CsGH3.1 in Arabidopsis led to restricted growth and a significant reduction in callus proliferation rates, further confirming its role in regulating…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
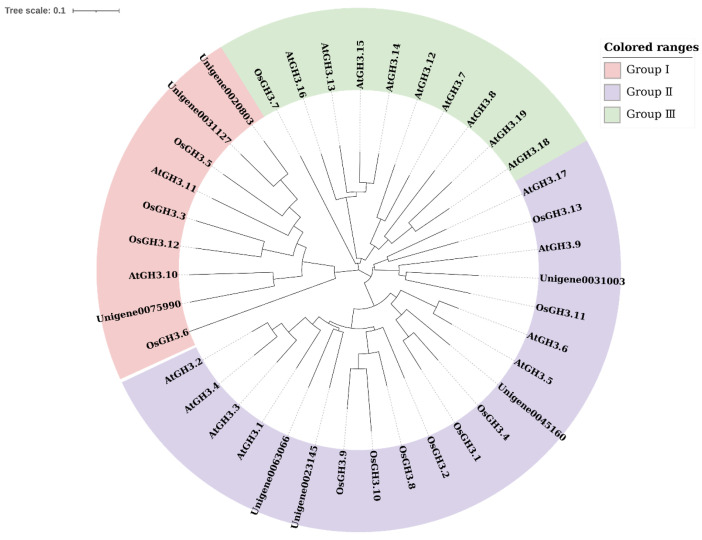 Figure 1
Figure 1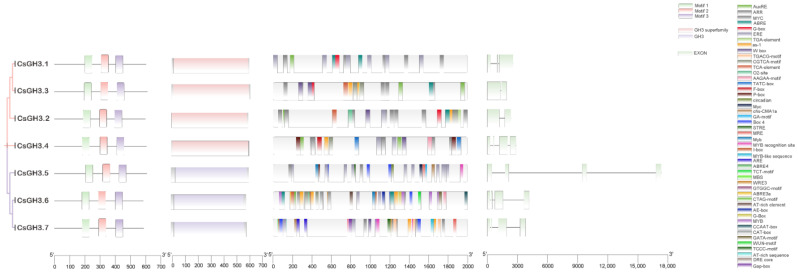 Figure 2
Figure 2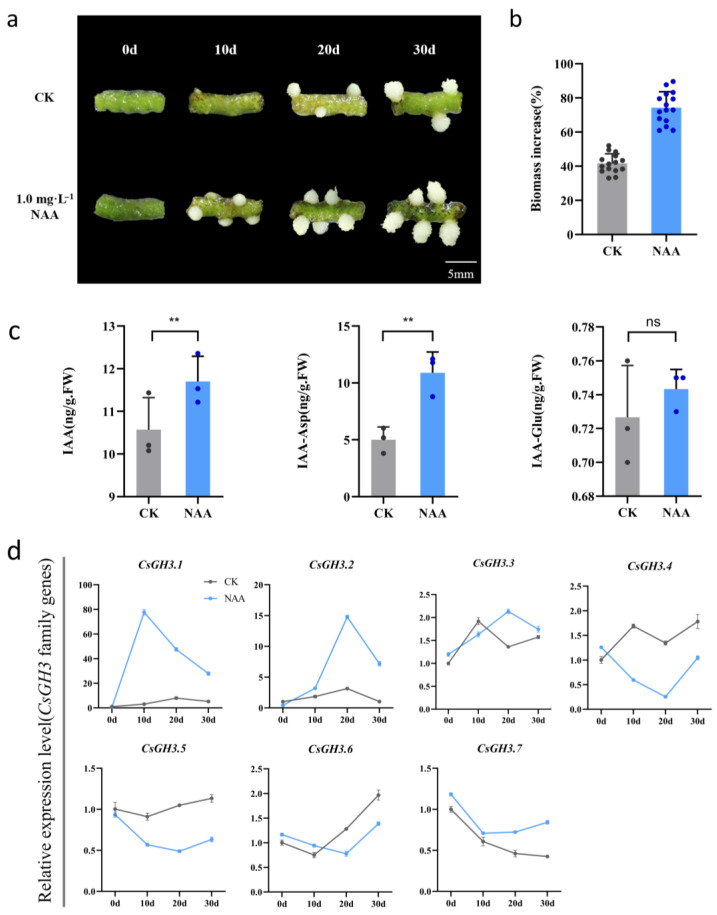 Figure 3
Figure 3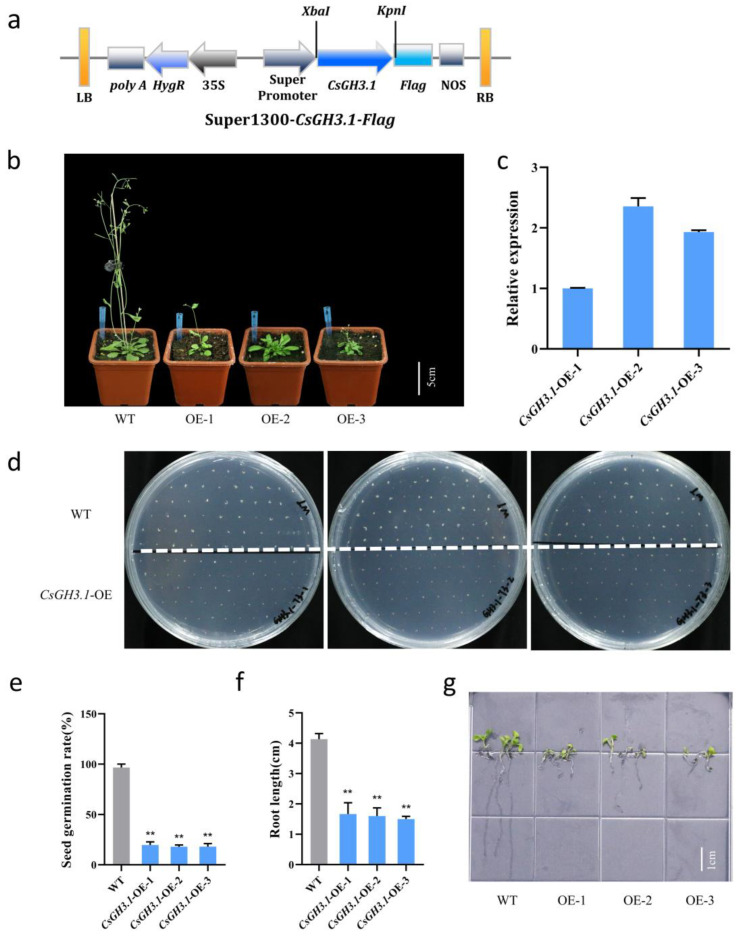 Figure 4
Figure 4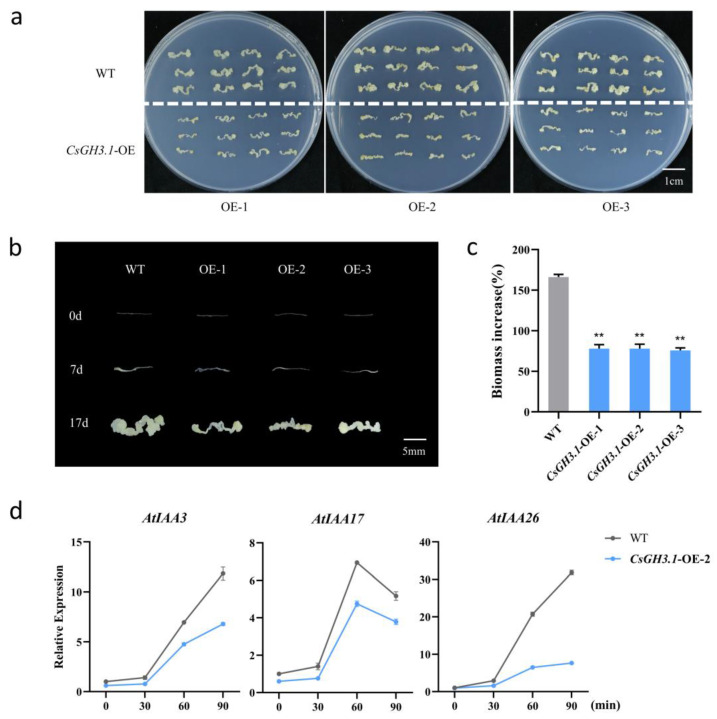 Figure 5
Figure 5- —Research and Development Plan in a Key Area of Guangdong Province, China
- —Science and Technology Program of Guangzhou
- —Science and Technology Program of Qingyuan City
- —Science and Technology Innovation Team Project of Qingyuan City
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Molecular Biology Research · Plant Reproductive Biology · Plant tissue culture and regeneration
1. Introduction
Plants regulate their growth and development in response to environmental signals, with hormones playing a key role, particularly in modulating auxin/indoleacetic acid (IAA) activity and content [1]. The IAA content in specific plant tissues is determined by various metabolic processes, including synthesis, transport, inactivation, reactivation, and degradation through oxidative pathways [2]. In addition to its free form, IAA exists in various modified forms, such as glucose conjugates and amide conjugates with amino acids and peptides [3]. The GRETCHEN HAGEN 3 (GH3) gene family, a acyl adenylate synthetase family member, is classified into three subfamilies based on similarities in protein sequences and substrate specificity [4]. Subfamily II GH3 genes are involved in auxin conjugation, catalyzing the binding of IAA to amino acids to form conjugated IAA. In contrast, subfamily I GH3 genes exhibit adenylation activity for jasmonic acid and catalyze the binding of jasmonic acid to isoleucine [5,6]. As one of the early auxin-responsive gene families, subfamily II GH3 genes play an important role in regulating auxin homeostasis [7]. When IAA concentrations increase, free IAA is catalyzed by GH3 amido synthetases to bind with amino acids, forming storage forms like IAA-Asp and IAA-Glu. These conjugates can either be converted back to free IAA via ILR1/ILL amido hydrolases or degraded through the GH3-ILR1-DAO pathway [6]. This interconversion between free and conjugated IAA helps maintain a dynamic balance in IAA levels.
The GH3 gene family typically includes multiple members across various plant species. In Arabidopsis, 19 GH3 genes have been identified on chromosomes 1, 2, 4, and 5 [6]. Similarly, 13 GH3 genes have been identified in rice, located on chromosomes 1, 5, 6, and 7 [8,9]. The GH3 gene family is also present in other plants such as maize, rapeseed, tomato, coffee, citrus, and potato [10,11,12,13,14]. In Arabidopsis, the expression of AtGH3.2/YDK1, AtGH3.5/AtGH3a/WES1, AtGH3.6/DFL1, and AtGH3.10/DFL2 is regulated by auxin. The overexpression of these genes results in dwarfism, reduced free auxin levels, and increased conjugated IAA-Asp levels [15,16,17,18]. The AtGH3.17/VAS2 knockout mutant (vas2) exhibits elongated hypocotyls and increased free IAA levels [2]. In rice, the overexpression of OsGH3.1 downregulates genes related to morphogenesis, cell differentiation, and metabolism [19], OsGH3.2 catalyzes auxin conjugation in response to drought stress [20,21], OsGH3.8 catalyzes the formation of IAA-Asp from IAA, Asp, and ATP, and the gain-of-function mutant tld1-D of OsGH3.13/TLD1 shows reduced free auxin levels, dwarfism, and decreased fertility [22,23,24,25].
Orchid tissue culture and rapid propagation are core technologies for orchid industrialization, primarily involving the induction, proliferation, and differentiation of intermediate propagules [26]. In terrestrial orchids, the rhizome is the intermediate propagule, but it proliferates slowly and is challenging to differentiate [26]. Cymbidium sinense ‘Qijianbaimo’ (CSQ), an important native flower in Guangdong Province, holds a high economic and ornamental value. Enhancing its regeneration capacity is crucial for large-scale propagation. Plant hormones play a vital role in embryogenesis, organogenesis, and plant regeneration [27]. The recently completed genome sequencing of Cymbidium sinense provides a valuable resource for investigating the molecular mechanisms underlying hormone regulation [28]. In tissue culture, exogenous plant hormones facilitate in vitro regeneration by activating specific developmental pathways [29]. Previous studies have shown that adding auxins such as naphthaleneacetic acid (NAA), indolebutyric acid (IBA), and IAA to the medium significantly increases rhizome proliferation rates. A preliminary transcriptomic analysis of CSQ during proliferation indicated that GH3 genes are highly expressed after exogenous NAA application, leading to a deceleration in the rate of endogenous auxin accumulation and limiting rhizome proliferation [26]. The GH3 genes, known for their role in conjugating endogenous auxin, likely play a significant inhibitory function during the proliferation of CSQ rhizomes, contributing to the observed slow proliferation phenotype.
To further investigate the role of GH3 genes in orchid tissue culture and rapid propagation, we conducted a comprehensive analysis of the CsGH3 gene family using the transcriptomic and genomic databases of CSQ, which identified seven CsGH3 genes. This study examines their phylogenetic relationships, conserved motifs, gene and protein structures, promoter cis-elements, subcellular localization, and expression profiles in response to exogenous auxin during rhizome proliferation. Furthermore, the CsGH3.1 gene was cloned, and transgenic Arabidopsis materials were generated through genetic transformation, confirming the auxin conjugation function of CsGH3.1 in explant proliferation. These findings provide a theoretical foundation for further research on the role of GH3 genes in orchid tissue culture, rapid propagation, and auxin-regulated propagation characteristics.
2. Results
2.1. The Genome of CSQ Contains Seven GH3 Genes
To obtain the GH3 protein sequences of CSQ, sequence alignment and protein domain analysis were performed using GH3 protein sequences from Arabidopsis and rice. Seven GH3 protein sequences were identified from the transcriptomic and genomic data of CSQ, located on chromosomes 2, 8, 9, 13, 14, and 15, and named CsGH3.1–CsGH3.7 (Table 1). The chemical properties of the predicted CsGH3 proteins, including molecular weight (MW), isoelectric point (IP), and chemical formula, were calculated using ProtParam (https://web.expasy.org/protparam// (accessed on 21 October 2023)). The molecular weights of all the CsGH3 proteins ranged from 65,500 to 69,000 Da, and their isoelectric points ranged from 5.0 to 7.0.
2.2. Phylogenetic Relationship of CsGH3s in CSQ
To study the evolution of the CsGH3 gene family, a Neighbor-Joining tree was constructed using the protein sequences of the 7 CsGH3s from CSQ, 19 AtGH3s from Arabidopsis, and 13 OsGH3s from rice (Figure 1). The GH3 gene family was divided into three clades in Arabidopsis, rice, and other plants [29]. The CsGH3s were found in subfamily I and subfamily II, with CsGH3.1-CsGH3.4 belonging to subfamily II (related to auxin conjugation) and CsGH3.5-CsGH3.7 belonging to subfamily I (related to jasmonic acid conjugation). The isoelectric points of the CsGH3 proteins also varied significantly between subfamilies, with subfamily I proteins showing isoelectric points between 5.5 and 5.8, and subfamily II proteins ranging from 6.2 to 6.3.
Clustal W was used for multiple sequence alignment. MEGA 7.0 was adopted for phylogenetic reconstruction using the Neighbor-Joining (NJ) clustering method. Circles of different colors represent different subfamilies.
2.3. Analysis of CsGH3 Gene Structures and Cis Elements in the CsGH3 Gene Promoters
To further study the diversity of CsGH3 protein structures and predict their functions, conserved motifs and domains were analyzed using the MEME 5.5.7 online software and the NCBI CDD website. The results revealed that all CsGH3 proteins contained three motifs. Specifically, CsGH3.1–CsGH3.4 from subfamily II exhibited complete GH3 superfamily conserved domains, while CsGH3.5–CsGH3.7 from subfamily I contained only the GH3 conserved domain (Figure 2). To predict the potential roles of CsGH3 family genes at different developmental stages, cis-elements in their promoter regions were identified using the PlantCARE database. The promoters of the CsGH3 genes contained many plant hormone response elements, including those related to auxin, jasmonic acid, gibberellin, cytokinin, salicylic acid, and abscisic acid (Figure 2). Gene structure analysis showed that the CsGH3 genes contained between one and four exons. CsGH3.1 contained three exons, CsGH3.2 and CsGH3.3 contained two exons, and CsGH3.4–CsGH3.7 had four exons (Figure 2).
2.4. Function of CsGH3 Genes During Rhizome Proliferation in CSQ
To study the function of the CsGH3 genes during rhizome proliferation in CSQ, the proliferation rate, auxin level, and gene expression level of CsGH3 were analyzed in rhizomes cultured on an MS medium with and without 1.0 mg·L^−1^ NAA (Figure 3). The results showed a significant increase in the proliferation rate of rhizomes after the exogenous application of 1.0 mg·L^−1^ NAA compared to the hormone-free control (Figure 3a,b). Hormone level measurements revealed that exogenous auxin did not significantly increase the free IAA levels during rhizome proliferation, but the conjugated IAA-Asp levels significantly increased (Figure 3c). These findings suggest that CSQ may maintain auxin homeostasis by converting free IAA into conjugated IAA-Asp during rhizome proliferation. Further analysis of CsGH3 gene expression in rhizomes under hormone-free and 1.0 mg·L^−1^ NAA conditions showed that CsGH3.1 and CsGH3.2 were significantly upregulated at 10, 20, and 30 days in response to NAA, with CsGH3.1 showing a faster and more pronounced increase in expression (Figure 3d). These results indicate that CsGH3.1 is crucial in auxin homeostasis during rhizome proliferation.
2.5. Overexpression of CsGH3.1 Leads to Decrease in Callus Proliferation Rate in Arabidopsis
To further investigate the functional role of CsGH3.1 in the proliferation culture, a Super1300-CsGH3.1-GFP expression vector was constructed and used to transform Arabidopsis via the floral dip method. Three independent Arabidopsis overexpression lines (OE-1, OE-2, and OE-3) were obtained (Figure 4). Compared to wild-type Arabidopsis, the three overexpression lines exhibited delayed flowering and a dwarf phenotype (Figure 4b).
The seed germination and early seedling development of CsGH3.1 overexpression lines were also significantly impaired. On 1/2 MS solid medium, wild-type seeds achieved a nearly 100% germination rate at 3 days post-sowing, while all three CsGH3.1 overexpression lines showed significantly reduced germination rates of approximately 20% (Figure 4d,e). Furthermore, at 7 days post-sowing, the wild-type seedlings developed primary roots measuring 4.13 ± 0.07 cm in length. In contrast, the overexpression lines displayed significantly shortened primary roots, with lengths of 1.67 ± 0.15 cm, 1.60 ± 0.11 cm, and 1.50 ± 0.04 cm for the three independent transgenic lines, respectively (Figure 4f,g). These results collectively demonstrate that CsGH3.1 overexpression significantly inhibits the early growth phases in Arabidopsis.
The roots from 7-day-old seedlings of wild-type Arabidopsis and the three T3 generation CsGH3.1 overexpression lines were used as explants to induce callus formation on the CIM medium (Figure 5). After 7 days, the callus was transferred to a 1/2 MS medium for 10 days of proliferation. All three CsGH3.1 overexpression lines showed 80–90% reduced callus proliferation compared to the wild-type controls (Figure 5a–c). These results suggest that the overexpression of CsGH3.1 in Arabidopsis negatively impacts plant growth and development, significantly reducing the proliferation efficiency of root explant-derived calluses.
Additionally, CsGH3.1 overexpression exhibited distinct effects on the expression of AtIAA3 in Arabidopsis (Figure 5d). While both the wild-type and CsGH3.1-OE-2 lines exhibited a rapid induction of AtIAA3, AtIAA17, and AtIAA26 during culture initiation, the transcript levels of these genes were consistently attenuated in the CsGH3.1-OE-2 plants relative to their wild-type counterparts (Figure 5d). These findings suggest that CsGH3.1 overexpression substantially impacts auxin signaling transduction in Arabidopsis, thereby reducing the plant’s sensitivity to auxin responses.
3. Discussion
As early auxin-responsive genes, the acyl-adenylate synthetase family GH3 plays a crucial role in maintaining auxin homeostasis in plants [20,30,31,32,33]. Although GH3 genes have been identified in many species, their functional characterization in orchids remains limited. Previous studies have identified 19 and 13 GH3 family genes in Arabidopsis and rice, respectively [31]. In this study, seven CsGH3 genes were identified from the transcriptome and genome of CSQ using bioinformatics methods. All CsGH3 proteins contained conserved GH3 domains and three conserved motifs. Based on their functional characteristics and sequence similarities, the GH3 proteins in Arabidopsis, rice, maize, wheat, and potato are classified into three subfamilies [9,34,35,36]. In Arabidopsis, subfamily II members such as AtGH3.1, AtGH3.2 (YDK), AtGH3.3, AtGH3.4, AtGH3.5 (AtGH3a), and AtGH3.6 (DFL1) catalyze the adenylation of IAA with amino acids [33,37]. Similarly, in rice, subfamily II members such as OsGH3-1, OsGH3-2, OsGH3-8, and OsGH3-13 are involved in auxin signaling. Phylogenetic analysis revealed that the CsGH3 genes in CSQ are clustered into subfamilies I and II (Figure 1), suggesting that subfamily II CsGH3 genes may also participate in auxin signaling. The promoters of soybean GH3 genes contain at least three auxin response elements (AuxREs) with a conserved TGTCTC sequence, which is essential for auxin responsiveness [38]. In this study, the promoters of four CsGH3 genes in subfamily II contained core AuxREs, while those in subfamily I lacked these elements (Figure 2). These results suggest that subfamily II CsGH3 genes are primarily involved in IAA signaling pathways.
GH3 genes respond rapidly to auxin signals and catalyze the conjugation of free IAA with amino acids to form auxin conjugates, thereby reducing endogenous auxin levels and maintaining auxin homeostasis [5,16,39,40]. In Arabidopsis thaliana, subfamily II AtGH3 genes, including AtGH3.1, AtGH3.2, AtGH3.3, and AtGH3.4 [30,41], which cluster with CsGH3.1 on the same evolutionary branch, are induced by auxin and are involved in synthesizing IAA-amido conjugates, thereby regulating auxin metabolic homeostasis. The expression of AtGH3.9 is suppressed by low concentrations of exogenous IAA, and its overexpression results in shorter primary roots, indicating its primary role in regulating root growth and development [35]. In contrast, AtGH3.17 is upregulated by auxin, and its enhanced expression is accompanied by increased levels of IAA-Glu and IAA-Asp. A loss of AtGH3.17 function leads to an accumulation of free IAA, suggesting its critical role in maintaining auxin homeostasis by modulating conjugated auxin levels [2]. These results suggest that different GH3 genes may exhibit specialized functional roles in regulating auxin metabolism. In this study, the CsGH3 subfamily II genes in Cymbidium sinense responded rapidly to auxin during rhizome proliferation, significantly increasing the conjugated auxin IAA-Asp (Figure 3c,d). Both CsGH3.1 and CsGH3.2 were rapidly induced by auxin, but CsGH3.1 showed a faster and more pronounced increase in expression (Figure 3d). These findings suggest that CsGH3.1 may play a more critical role in regulating auxin homeostasis during rhizome proliferation in Cymbidium sinense. It is proposed that CsGH3.1 promotes the conjugation of IAA with amino acids, reducing free IAA levels and thereby regulating rhizome proliferation and development.
Maintaining active IAA levels is essential for normal plant growth and development. Previous studies have shown that overexpressing GH3 genes results in auxin-deficient phenotypes and developmental abnormalities [42]. For example, in Arabidopsis, the auxin-responsive gene AtGH3.6/dfl1-D negatively regulates hypocotyl and lateral root elongation, resulting in a dwarf phenotype [35]. In rice, the overexpression of OsGH3.8 and OsGH3.13 causes slow growth and dwarfism, accompanied by increased levels of IAA-Asp and IAA-Glu conjugates [23,43]. In this study, three CsGH3.1 overexpression Arabidopsis lines displayed dwarfism, delayed flowering, and reduced growth rates (Figure 4a), which is similar to the phenotypes observed in most GH3 gene overexpression mutants. Similarly, the heterologous overexpression of OsGH3.2 and OsGH3.8 in Arabidopsis resulted in slow growth, dwarfism, and smaller leaves. During callus culture, callus proliferation was significantly reduced in all three CsGH3.1 overexpression lines (Figure 4c–e). These results indicate that the overexpression of CsGH3.1 causes growth abnormalities resembling auxin-deficient phenotypes, highlighting its critical role in regulating auxin levels and homeostasis. We propose that CsGH3.1 modulates rhizome proliferation in Cymbidium sinense by conjugating auxin and reducing free IAA levels.
4. Materials and Methods
4.1. Identification and Bioinformatics Analysis of the GH3 Gene Family in CSQ
GH3 gene family protein sequences from Arabidopsis and rice were retrieved from the NCBI database and were used for a BLAST 1.30 search against the transcriptomic and genomic databases of CSQ to preliminarily identify GH3 genes. The presence of GH3 protein domains in the candidate genes was confirmed by predicting conserved domains using the protein domain prediction tool (https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi (accessed on 22 October 2023)). A phylogenetic tree was constructed using MEGA7 to assess the homology of candidate genes with known GH3 genes [44]. MEME was utilized to predict conserved motif elements in the identified CsGH3s (https://meme-suite.org/meme/index.html (accessed on 25 October 2023)). The 2 kb upstream region of the first exon of the CsGH3s was selected as the promoter region, and cis-elements were predicted using the PlantCARE website and visualized using TBtools v0.665 [45].
4.2. Rhizome Proliferation Culture of CSQ
The rhizomes of CSQ, which were originally induced from shoot apices and subsequently maintained through successive subcultures on a hormone-free MS medium under continuous darkness, served as the experimental material. The vigorous rhizomes of CSQ were excised, and the apical growth points were removed. The rhizomes were cut into approximately 1 cm segments and inoculated evenly onto the MS medium with or without 1.0 mg·L^−1^ NAA [26]. Each treatment was conducted with three biological replicates, with five bottles per replicate and ten rhizomes per bottle. After inoculation, the bottles were sealed with two layers of opaque black plastic bags and cultured in a 26 °C incubator for 30 days. The weight of the rhizomes was measured at the beginning and after 30 days of culture, and the morphology of the rhizomes was photographed for analysis.
4.3. Endogenous Hormone Measurement
Samples of the CSQ rhizomes grown under hormone-free and 1.0 mg·L^−1^ NAA conditions were collected at 0, 10, 20, and 30 days for endogenous hormone analysis. Ten rhizomes were randomly selected from each treatment, with three biological replicates. The samples were rapidly frozen in liquid nitrogen and stored at −80 °C. These samples were ground to a fine powder in liquid nitrogen using a mortar and pestle, then transferred to 10 mL centrifuge tubes with pre-chilled spatulas for fresh weight measurement. The quantification of endogenous auxins (IAA, IAA-Asp, and IAA-Glu) was conducted via ESI-HPLC-MS/MS using an Agilent 1290 UHPLC system coupled with electrospray ionization (ESI) and multiple reaction monitoring (MRM) (Table 2). Mass spectrometry data were processed using Analyst software 1.7 (Nanjing Ruiyuan Biotechnology Co., Ltd., Nanjing, China).
4.4. Expression Analysis of CsGH3 Genes
The CSQ rhizomes grown under hormone-free and 1.0 mg·L^−1^ NAA conditions were collected at 0, 10, 20, and 30 days. Six to seven rhizomes were randomly selected from each treatment, with three biological replicates. The samples were quickly frozen in liquid nitrogen and stored at −80 °C for a subsequent qPCR analysis of the gene expression patterns of their CsGH3 genes.
Total RNA was extracted from the samples using the RNA Aprep Pure Plant Kit (Vazyme, Nanjing, China), following the manufacturer’s method. cDNAs were reverse transcribed using 5 HiScript III qRT SuperMix (Vazyme, Nanjing, China) and used for a subsequent quantitative analysis of candidate genes. The transcript levels of the genes were detected by qPCR using AceQ qPCR SYBR Green Master Mix (Vazyme) and the CFX96 Real-Time PCR Detection System (Bio-Rad, Hercules, CA, USA) in standard mode. The gene-specific primers are listed in Table 3, and the petunia RPS3 gene was used as an internal control. The reaction conditions were 30 s at 95 °C and 40 cycles of 5 s at 95 °C and 30 s at 60 °C. Three biological replicates were performed in this study, and the expression data were quantified via the 2^−△△CT^ method [46].
4.5. Generation of Overexpressed-CsGH3.1 Arabidopsis Transgenic Materials
The CDS sequence of CsGH3.1 was amplified and cloned into the Super1300-GFP vector to construct the overexpression vector. After Arabidopsis bolting, the opened flowers on the inflorescence were removed, leaving only the unopened flower buds. The inflorescence tip was immersed in the infection solution containing the plasmid for 3–5 s, after which the plants were kept in the dark overnight before being transferred to normal growth conditions. Approximately one month after transformation, dried and cracked siliques were collected, dried, and sown. Antibiotic selection was used to identify successful transformants. The genomic DNA of the selected transgenic lines was analyzed by PCR using hygromycin-specific primers (Hyg-F: 5′-GACATTGGGGAGTTTAGCGAGAGC-3′; Hyg-R: 5′-GGTGTCGTCCATCACAGTTTGCC-3′) to identify hygromycin-positive plants (Figure S1). The DNA of each sample was extracted using the CTAB method, and 2 μL was used as a template for PCR. PCR amplification was performed in a 50 µL reaction containing 2 µL primers, 25 µL 2 × Phanta Flash Master Mix (Dye Plus, Vazyme, Nanjing, China), and 19 µL ddH_2_O. The reaction conditions were 30 s at 98 °C, 30 cycles of 10 s at 98 °C, 5 s at 58 °C, 5 s at 72 °C, and finally, 60 s at 72 °C. RNA was extracted from the leaves of the T1 transgenic plants, and qPCR was performed to confirm the overexpression of CsGH3.1 [47].
4.6. Phenotypic Analysis of CsGH3.1 Overexpression Arabidopsis Mutants
Three CsGH3.1 overexpression Arabidopsis lines and a wild-type Arabidopsis were sown simultaneously, and their phenotypes were recorded after 50 days. T1 mutant seeds were surface-sterilized and cold-treated for 24 h before being germinated on 1/2 MS medium. After 7 days, the roots were cut into 1 cm segments and cultured on CIM medium (Gamborg’s B5 basal salts supplemented with 20.0 g·L^−1^ sucrose, 0.5 g·L^−1^ MES, 0.5 mg·L^−1^ 2,4-D, 0.05 mg·L^−1^ kinetin, and 5 g·L^−1^ agar, pH adjusted to 5.8) for 7 days to induce callus formation. The callus was then transferred to a hormone-free B5 medium for proliferation, and the biomass increment was measured at 0 and 10 days [48].
4.7. Statistical Analysis
Biomass increment of rhizomes, endogenous hormone levels, and relative gene expression levels were analyzed from three biological replicates. Data were processed using GraphPad to calculate least significant difference at p = 0.05, and results were expressed as mean ± standard error of mean (SEM) (n = 3). Statistical significance was determined at p = 0.05 [47].
5. Conclusions
This study represents the first systematic identification of the GH3 gene family in Cymbidium sinense and explores its potential roles in auxin and jasmonic acid metabolism. Evolutionary analysis revealed that CsGH3 genes are divided into two subfamilies. Gene structure and cis-element analyses highlighted the significant role of CsGH3 genes in hormone responses. CsGH3.1 was significantly upregulated in response to auxin during rhizome proliferation in CSQ, suggesting its key role in regulating auxin homeostasis. The overexpression of CsGH3.1 in Arabidopsis significantly inhibited callus proliferation, confirming its involvement in cell proliferation and development, as well as in auxin homeostasis. These findings enhance our understanding of the functional roles of the GH3 gene family and provide a theoretical foundation for molecular breeding and hormone regulation research for Cymbidium sinense. Future studies should focus on the functional validation of CsGH3.1 and its regulatory networks in rhizome development, offering insights for orchid breeding and cultivation.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Woodward A.W. Bartel B. Auxin: Regulation, action, and interaction Ann. Bot.20059570773510.1093/aob/mci 08315749753 PMC 4246732 · doi ↗ · pubmed ↗
- 2Zheng Z. Guo Y. Novák O. Chen W. Ljung K. Noel J.P. Chory J. Local auxin metabolism regulates environment-induced hypocotyl elongation Nat. Plants 201621602510.1038/nplants.2016.2527249562 PMC 4849989 · doi ↗ · pubmed ↗
- 3Mateo-BonmatíE. Casanova-Sáez R. Šimura J. Ljung K. Broadening the roles of UDP-glycosyltransferases in auxin homeostasis and plant development New Phytol.202123264265410.1111/nph.1763334289137 · doi ↗ · pubmed ↗
- 4Westfall C.S. Zubieta C. Herrmann J. Kapp U. Nanao M.H. Jez J.M. Structural basis for prereceptor modulation of plant hormones by GH 3 proteins Science 20123361708171110.1126/science.122186322628555 · doi ↗ · pubmed ↗
- 5Luo P. Li T.-T. Shi W.-M. Ma Q. Di D.-W. The roles of GRETCHEN HAGEN 3 (GH 3)-dependent auxin conjugation in the regulation of plant development and stress adaptation Plants 202312411110.3390/plants 1224411138140438 PMC 10747189 · doi ↗ · pubmed ↗
- 6Staswick P.E. Serban B. Rowe M. Tiryaki I. Maldonado M.T. Maldonado M.C. Suza W. Characterization of an Arabidopsis enzyme family that conjugates amino acids to indole-3-acetic acid Plant Cell 20051761662710.1105/tpc.104.02669015659623 PMC 548830 · doi ↗ · pubmed ↗
- 7Hagen G. Guilfoyle T. Auxin-responsive gene expression: Genes, promoters and regulatory factors Plant Mol. Biol.20024937338510.1023/A:101520711411712036261 · doi ↗ · pubmed ↗
- 8Fu J. Yu H. Li X. Xiao J. Wang S. Rice GH 3 gene family: Regulators of growth and development Plant Signal. Behav.2011657057410.4161/psb.6.4.1494721447996 PMC 3142395 · doi ↗ · pubmed ↗