Single-Nucleotide Polymorphisms of BRCA1 and BRCA2 and Risk of Papillary Thyroid Carcinoma
Chang Myeon Song, Yun Jin Kim, Hyun Sub Cheong, Yong Bae Ji, Kyung Tae

TL;DR
This study found that certain genetic variations in the BRCA1 gene are linked to a lower risk of developing papillary thyroid cancer in the Korean population.
Contribution
The study identifies specific BRCA1 SNPs and haplotypes associated with reduced susceptibility to papillary thyroid carcinoma in Koreans.
Findings
BRCA1 SNPs rs8176318, rs1799966, rs16940, rs799917, and rs1799949 were significantly associated with reduced papillary thyroid carcinoma risk.
Haplotypes of BRCA1, including rs8176318(C)-rs1799966(T)-rs799917(G)-rs16940(A)-rs1799949(G), were linked to decreased PTC risk.
Certain genotypes of BRCA1 SNPs showed odds ratios of 0.67–0.70, indicating reduced cancer risk.
Abstract
This study was conducted to analyze the association between genetic polymorphisms of breast cancer genes 1 (BRCA1) and 2 (BRCA2), which are well known for their association with breast cancer and the development of thyroid cancer. We analyzed the relationship between single-nucleotide polymorphisms (SNPs) in BRCA1 and BRCA2 and thyroid cancer and conducted a haplotype analysis of these polymorphisms to study the interactions between genetic polymorphisms. The results of this study show that polymorphisms of BRCA1 were significantly related to the susceptibility to papillary thyroid carcinoma in the Korean population. Background/Objectives: We sought to evaluate the association between the risk of papillary thyroid carcinoma (PTC) and single-nucleotide polymorphisms (SNPs) of breast cancer genes 1 (BRCA1) and 2 (BRCA2). Methods: We prospectively recruited 515 cases with PTC and 296…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
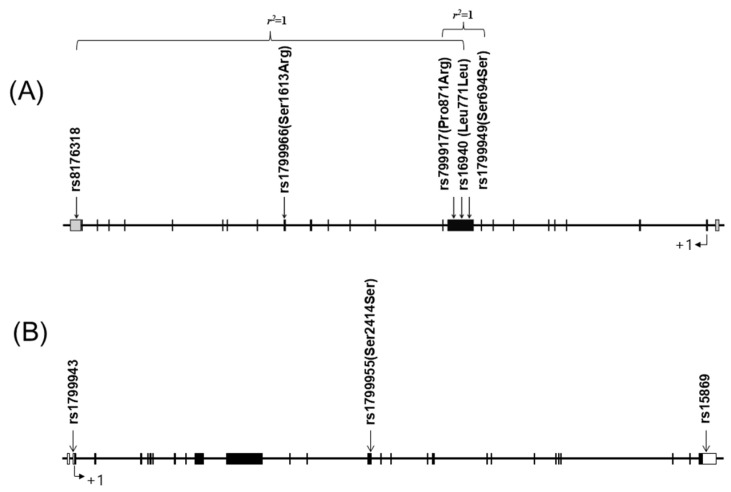 Figure 1
Figure 1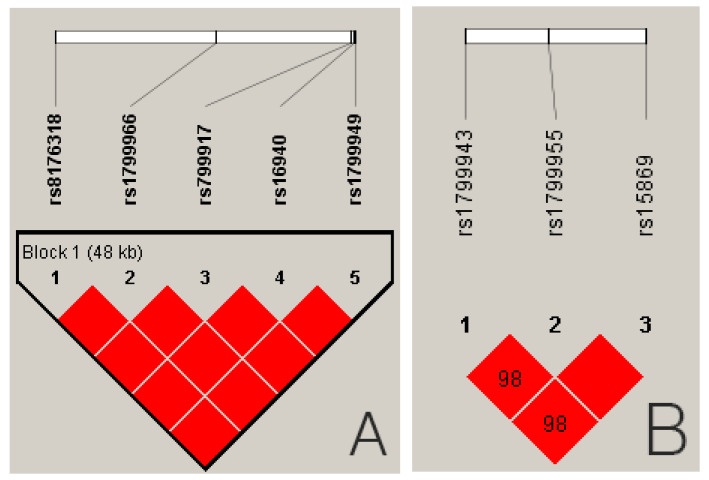 Figure 2
Figure 2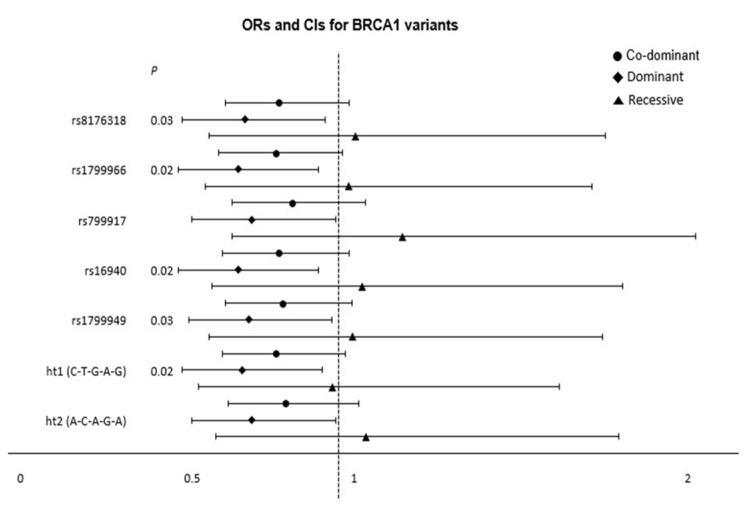 Figure 3
Figure 3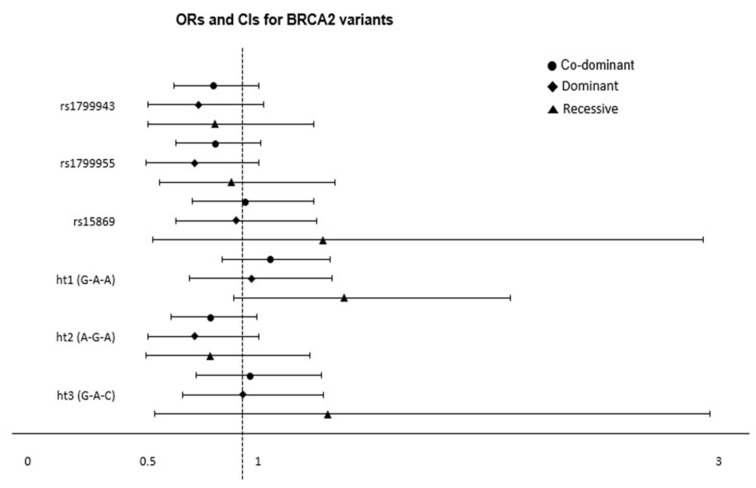 Figure 4
Figure 4- —Hanyang University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBRCA gene mutations in cancer · Genetic Associations and Epidemiology · Nutrition, Genetics, and Disease
1. Introduction
Thyroid cancer includes differentiated carcinoma, such as papillary and follicular carcinoma, as well as non-differentiated cancers, including medullary, anaplastic carcinoma, and lymphoma. Thyroid cancer accounts for approximately 2% of all types of cancer diagnosed worldwide, and 3.5–4% of those are diagnosed in the United States [1,2]. Its incidence rates are four times higher among females than males [3]. However, the cause of this gender difference is not well known.
Ionizing radiation is an established risk factor for differentiated thyroid cancer, and its association with genetic changes is well known [4]. The incidence of thyroid cancer increases with exposure to environmental radiation, including that caused by the Chernobyl nuclear power plant accident and the explosion of atomic bombs [5,6,7]. The latency period is at least 5 to 10 years, and molecular alterations, including gene rearrangements, are frequently implicated [8]. Experimental radiation exposure results in genetic changes and tumorigenicity in thyroid epithelial cell lines, supporting the evidence for the effect of radiation on genetic changes related to thyroid cancer [9]. Furthermore, the risk of thyroid cancer is elevated 8-fold in first-degree relatives of thyroid cancer patients [10]. Therefore, an investigation of the basis of genetic predisposition of thyroid cancer is needed to fully understand the pathogenesis of differentiated thyroid carcinoma. However, familial inheritance can be related to both genetic predisposition and a shared common environment. A nationwide cohort study of Asian patients with non-medullary thyroid cancer reported that genetic and common environmental contributions to first-degree relative thyroid cancer were 28% and 14%, respectively [11]. Relative risks for non-medullary thyroid cancer for siblings, offspring, and parents were 6.4, 5.2, and 5.1, respectively.
Studies of the association of genetic variation and thyroid cancer have included several genome-wide association studies [11,12,13,14,15]. Various single-nucleotide polymorphisms (SNPs) have been shown to be associated with the risk of non-syndromic differentiated thyroid carcinoma [16,17,18,19]. Genes related to thyroid cancer development are involved in DNA repair, ATM (ataxia–telangiectasia mutated) signaling, RET (receptor tyrosine kinase) signaling, and the regulation of thyroid activity (FOXE1, NKX2-1, TSHR) [7]. The DNA repair systems associated with thyroid cancer development are the direct repair, excision repair, recombinant repair, and mismatch repair pathways [20]. SNPs that are associated with radiation-related thyroid cancer and ionizing radiation exposure in thyroid cancer patients include FOXE1, MGMT, ALKBH3, LIG1, XRCC2, NKX2-1, ATM, and TP53 [21,22,23,24,25].
BRCA1 and BRCA2 are tumor suppressor genes involved in DNA damage repair before cell division is initiated [26]. The BRCA1 DNA repair-associated protein (BRCA1) is implicated in DNA damage repair and transcription repression and acts as a cell cycle checkpoint [27]. It also interacts with tumor suppressors, DNA damage repair proteins, oncogenes, transcriptional activators/repressors, and cell cycle regulators [27]. The BRCA1- and BRCA1-interacting protein C-terminal helicase 1 (BRIP1) complex plays an important role in double-strand break repair. Mutation of BRCA1 results in abnormal apoptosis, cell cycle checkpoint defects, chromosome instability, aneuploidy, and impaired DNA damage repair [28]. In contrast, the BRCA2 DNA repair-associated protein (BRCA2) regulates the intracellular shuttling and activity of RAD51, a recombinase enzyme active in DNA repair via homologous recombination [29].
Mutations in BRCA1 and BRCA2 are associated with a risk of breast, ovary, prostate, bladder, and pancreatic cancer, as well as melanoma and other skin cancers [30]. BRCA1 mutations have also been identified in patients with double primary cancers of the breast and thyroid [31,32]. However, the association of BRCA1/BRCA2 genetic polymorphism with the risk of papillary thyroid carcinoma (PTC) has not been thoroughly investigated. We aimed to prioritize the investigation of SNPs to facilitate the subsequent selection of additional genetic targets. In this study, we analyzed the single-nucleotide polymorphisms of BRCA1/BRCA2 in patients with PTC and controls to assess the association of BRCA1/BRCA2 polymorphism and the risk of PTC. To the best of our knowledge, this study is the first to examine the association between BRCA1/BRCA2 polymorphism and the risk of PTC in the Korean population.
2. Materials and Methods
2.1. Subjects
We prospectively enrolled patients with pathologically confirmed PTC, who had undergone surgery at a tertiary hospital. Although both PTC and follicular carcinoma are classified as differentiated carcinomas, we limited our study to PTC in order to maintain homogeneity. The controls consisted of healthy volunteers who visited the hospital for health check-ups or patients who were admitted to the hospital for treatment of upper respiratory infections or for operations on benign lesions of the ear, nose, and throat. We included subjects over 18 years of age of Korean nationality who resided in South Korea. We excluded subjects with previous or simultaneous histories of malignancy other than thyroid cancer or histories of genetic disease. Patients who received radiation therapy were excluded. Finally, 811 subjects (515 patients with PTC and 296 controls without cancer) were included in this study. This study was approved by the Institutional Review Board of our tertiary hospital. Informed consent was obtained from all the participants. Given the potential genetic variations between papillary thyroid carcinoma and other thyroid cancer types, this study was limited to patients with papillary thyroid carcinoma to maintain cohort homogeneity.
2.2. Selection and Genotyping of Single-Nucleotide Polymorphisms
Genotypes with respect to five BRCA1 SNPs (rs8176318, rs1799966, rs799917, rs16940, rs1799949) and three BRCA2 SNPs (rs15869, rs1799943, rs1799955) were determined. These 8 SNPs have been associated with the risk of malignancies of multiple organs in the published literature, and their minor allele frequencies exceeded 10% based on the data provided by National Cancer for Biotechnology Information (http://www.ncbi.nlm.nih.gov/snp, accessed on 7 August 2018) [33,34,35,36,37].
The genotypes were determined using the TaqMan assay method on an ABI prism 7900HT sequence detection system (Applied Biosystems, Foster City, CA, USA), and the used polymerase chain reaction sequences and reaction conditions have been summarized in a previous report [38]. A summary of the SNPs and the probes used in this study is given in Table 1. We also evaluated the potential association of haplotypes with the risk of PTC based on the findings of linkage disequilibrium (LD). Originally, linkage disequilibrium (LD) analysis was conducted for different genes. However, in this study, we analyzed LD based on the assumption that the eight SNPs were located in different positions.
2.3. Statistical Analysis
Continuous variables and categorical variables were analyzed using Student’s t test and Chi-square test, respectively. We evaluated Lewontin’s D′ (|D′|) and the LD coefficient r2 between all the pairs of bi-allelic loci using Haploview v4.2 software, provided by the Broad Institute (Cambridge, MA, USA) (http://www.broadinstitute.org/mpg/haploview, accessed on 7 August 2018) [39]. The haplotypes were inferred using PHASE software (https://stephenslab.uchicago.edu/phase/download.html, accessed on 7 August 2018), and statistical analysis was performed using Statistical Analysis System (SAS) version 9.4 (SAS Inc., Cary, CA, USA) [40]. Logistic regression analyses were performed to compare genotype distributions between the PTC cases and controls, calculating the odds ratio, 95% confidence intervals (CIs), and the corresponding p-values. An association analysis was performed using a referent model and 3 alternatives. The referent model compared heterozygotes vs. major homozygotes and minor homozygotes vs. major homozygotes. The co-dominant model compared minor allele homozygotes vs. heterozygotes vs. major allele homozygotes. The dominant model for minor alleles compared minor allele homozygotes plus heterozygotes vs. major allele homozygotes. The recessive model compared minor allele homozygotes vs. heterozygotes plus major allele homozygotes. We considered p < 0.05 as statistically significant.
3. Results
3.1. Genotyping
The demographic characteristics of the participants are summarized in Table 2. There was no significant difference in age between the PTC group and controls. However, the proportion of female patients was higher in the PTC group (75.9% vs. 66.2%, p = 0.003). To avoid bias, all the logistic regression analyses were adjusted for age as a continuous covariate and sex as a categorical covariate.
The five SNPs of BRCA1 were all located on chromosome 17q21.31, and the three SNPs of BRCA2 were located on chromosome 13q13.1 (Figure 1A,B). These eight SNPs were genotyped in 811 study subjects. For the BRCA1 genes, genotyping for rs8176318, rs1799966, rs799917, rs16940, and rs1799949 was successful in 97.3%, 97.2%, 98.8%, 96.7%, and 99.1% of the subjects, respectively. For the BRCA2 genes, genotyping for rs1799943, rs1799955, and rs15869 was successful in 98.6%, 98.2%, and 96.1% of the subjects. The quality of genotyping was assessed by duplicate verification in 10% of the samples (rate of concordance in duplicates = 100%).
The alleles, the distribution of genotypes, the frequencies of homozygosity, heterozygosity, and minor alleles, and the tests for deviation from the Hardy–Weinberg equilibrium are listed in Table 3. The distributions of genotypes for all eight genes were not significantly different from the Hardy–Weinberg equilibrium. Minor allele frequency ranged from 0.196 (for rs15869) to 0.406 (rs1799955).
An analysis of the linkage disequilibrium revealed that the five BRCA1 SNPs (rs8176318, rs1799966, rs799917, rs16940, rs1799949) were in complete linkage disequilibrium (all D’= 1.000, Figure 2A). The r^2^ values for the BRCA1 SNPs ranged from 0.985 to 1.000, and the three BRCA2 SNPs (rs1799943, rs1799955, rs15869) were in strong LD (all D’ ≥ 0.98, Figure 2B). However, the r^2^ values were 0.169 between rs1799955 and rs15869, 0.152 between rs1799943 and rs15869, and 0.910 between rs1799943 and rs1799955. We also used the five BRCA1 SNPs and three BRCA2 SNPs consecutively to construct haplotypes. This yielded four haplotypes for the BRCA1 SNPs, of which the two major ones with a frequency exceeding 1% were selected for further statistical analysis. For the SNPs, we constructed six haplotypes, four of which were used for further haplotype analysis. The data for haplotype construction are shown in Appendix A Table A1.
3.2. Logistic Regression Analysis of the Association of BRCA1 and BRCA2 Polymorphisms with Thyroid Cancer
We performed a logistic regression analysis to evaluate the associations between BRCA1 and BRCA2 SNPs and the risk of PTC, adjusting for age and sex (Figure 3 and Figure 4, Table A2, Table A3 and Table A4). Five BRCA1 SNP genotypes were significantly associated with the risk of PTC. The AC genotype of rs8176318 was associated with a decreased risk of PTC in the referent model (OR = 0.69, 95% CI = 0.51–0.94, p = 0.02) and in the dominant model (OR = 0.72, 95% CI = 0.53–0.96, p = 0.03). Similarly, the CT genotype of rs1799966 was associated with a decreased risk of PTC in the dominant model (OR = 0.70, 95% CI = 0.52–0.94, p = 0.02), and the CC genotype was associated with a decreased risk of PTC in the referent model (OR = 0.67, 95% CI = 0.49–0.91, p = 0.01). The AG genotype of rs799917 was significantly associated with the risk of PTC in the referent model (OR = 0.70, 95% CI = 0.52–0.95, p = 0.02), and the AG genotype of rs16940 was associated with a decreased risk of PTC both in the referent (OR = 0.67, 95% CI = 0.49–0.91, p = 0.01) and dominant (OR = 0.70, 95% = 0.52–0.94, p = 0.02) models. Finally, the AG genotype of rs1799949 was associated with a decreased risk of PTC in both the referent (OR = 0.70, 95% CI = 0.52–0.95, p = 0.02) and dominant (OR = 0.73, 95% CI = 0.55–0.98, p = 0.03) models. Contrarily, there were no significant associations between the genotypes of the three BRCA2 SNPs and PTC.
Logistic regression with subgroup analyses was performed under the co-dominant, dominant, and recessive models to calculate the ORs (95% CI) and p-values. Each plot represents the point estimate of ORs on the x-axis, with 95% CIs represented by error bars. Significant associations are indicated by a p-value < 0.05.
3.3. Association of BRCA1 and BRCA2 Haplotypes with Thyroid Cancer
The distributions of the haplotype genotypes of BRCA1 and BRCA2 and the association of each haplotype with the risk of PTC are summarized in Table A4 and Figure 3 and Figure 4. Haplotype 1 [rs8176318(C)-rs1799966(T)-rs799917(G)-rs16940(A)-rs1799949(G)] ± of BRCA1 was associated with a significantly decreased risk of PTC in the referent model (OR = 0.69, 95% CI = 0.51–0.93, p = 0.02) and the dominant model (OR = 0.71, 95% CI = 0.53–0.95, p = 0.02). Haplotype 2 [rs8176318(A)-rs1799966(C)-rs799917(A)-rs16940(G)-rs1799949(A)] ± of BRCA1 was associated with a decreased risk of PTC in the referent model (OR = 0.71, 95% CI = 0.52–0.96, p = 0.03) and the dominant model (OR = 0.74, 95% CI = 0.56–0.99, p = 0.04). Haplotypes of BRCA2 genes were not significantly associated with the risk of thyroid cancer.
4. Discussion
The results of this study suggest that BRCA1 is significantly associated with the pathogenesis of thyroid cancer, while BRCA2 seems to have little or no association with the risk of PTC. Notably, all five of the evaluated BRCA1 SNP genotypes were significantly associated with the risk of PTC, and there was no significant association between any of the three BRCA2 SNPs and PTC. The five BRCA1 SNPs (rs8176318, rs1799966, rs799917, rs16940, rs1799949) were in complete linkage disequilibrium (all D’= 1.000, r^2^ = 0.9851.000), and the three BRCA2 SNPs (rs1799943, rs1799955, rs15869) were in strong LD (all D’ ≥ 0.98, r^2^ = 0.1520.910). Two haplotypes of the five BRCA1 SNP genes and four haplotypes of the three BRCA2 SNP genes were constructed with frequencies greater than 1%. BRCA1 haplotype 1 (CTGAG in rs8176318, rs1799966, rs799917, rs16940, and rs1799949) and BRCA1 haplotype 2 (ACAGA in rs8176318, rs1799966, rs799917, rs16940, and rs1799949) were significantly associated with a decreased risk of PTC. The haplotypes of the BRCA2 genes were not associated with the risk of PTC. Although the LD analysis was performed on the basis that the SNPs were not in identical locations, caution is needed in interpretation because three SNPs, rs799917, rs16940, and rs1799949, are located in proximity.
BRCA1 is located at chromosome 17q21 and contains 24 exons that are translated into 1863 amino acids [41]. BRCA1 is involved in signaling DNA damage by forming macromolecular complexes that trigger the repair of double-stranded DNA (dsDNA) breaks with proteins that repair chromosomal damage [42,43]. It is also involved in multiple phases of the cell cycle by interacting with proteins such as p53, p21, BARD1, ATR, and ATM. The deficiency of BRCA1 may result in alterations in the S-phase, G2/M, and cell cycle spindle checkpoints [27]. BRCA2 rs1799943 is located in an untranslated site of chromosome 13. BRCA2 directly controls the production of RAD51 and repairs DNA breaks through homologous recombination [28]. In BRCA2-deficient cells, the sensitivity to radiation is increased, leading to defective dsDNA break repair [44].
Mutations in pathogenic variants of BRCA1 and BRCA2 are reported to lead to lifetime risks of developing breast and ovarian cancer, as high as 84 and 45%, respectively [26]. In another study, carriers of impaired BRCA1 genes had a cumulative risk of breast cancer of 72% by the age of 80, and for BRCA2, the corresponding risk was 69% [45]. In addition, the frequency of pathological genetic mutations in BRCA1 and BRCA2 in sporadic breast cancers in the South Korean population was found to be 3.1% [46]. It is unclear why breast and ovarian cancers are especially associated with BRCA defects since the BRCA proteins are active in all cell types. The association of BRCA1 and BRCA2 mutations with thyroid carcinoma has not been well evaluated. Xu et al. examined eight BRCA1 SNPs in 303 cases of differentiated thyroid carcinoma in the United States [47]; haplotypes that carried the rs799917 variant allele were significantly associated with a decreased risk of thyroid cancer, and the AG/GG genotypes of rs1799950 were associated with a reduced cancer risk, with an adjusted odds ratio of 0.31. These observations agree with our finding that the AG genotype of rs799917 is associated with a reduced risk of PTC. The SNP of rs1799966 was not significantly associated with the risk of differentiated thyroid cancer in that study, unlike in this study. This may be due to the difference in ethnicity of the two populations. A study of a Chinese ethnic population evaluated the association of SNPs in the 3′-UTR double-strand break repair genes RAD51, RAD51B, BRCA1 (rs12516, rs8176318), BRCA2 (rs15869), XRCC4, and XRCC5 with the risk of thyroid cancer in 206 patients with PTC [48]. The CC genotype of BRCA2 rs15869 was associated with a greater risk of PTC than the AA genotype (OR = 2.595, p = 0.031), unlike in our study. Another study evaluating the genetic polymorphisms of the ATM–CHEK2–BRCA1 axis in 1781 Polish patients with PTC [10] found that CHEK2 rs17879961 and BRCA1 rs16941 (OR = 1.6, p = 0.005) were risk alleles for PTC. BRCA1 SNP rs16940 was also associated with the risk of PTC in our study, and we did not evaluate rs16941.
BRCA1 mutations may also be associated with responses to chemotherapy and cancer survival. One study reported that the polymorphisms of BRCA1 rs799917 were associated with the response to chemotherapy and the overall survival of Korean lung cancer patients [49]. The TC/CC genotypes of BRCA1 rs1799966 were associated with a lower risk of death (hazard ratio = 0.617, p = 0.028), and the C allele (TC + CC vs. TT) of rs1799966 was associated with a good response to chemotherapy (OR = 0.402, p = 0.008) in patients with lung cancer in the Chinese population [50]. This result is interesting because the CT and CC genotypes of rs1799966 were associated with a lower risk of PTC incidence in our study. In another study, the CC genotype of rs1799966 was associated with significantly longer survival in non-small-cell lung cancer patients (Bonferroni-adjusted p = 0.012) [36].
The effect of BRCA1 mutations in thyroid cancer patients is partially understood. The SNPs of BRCA1 rs1799949 are nonsense mutations and are associated with breast cancer in Taiwan [51]. Another study confirmed a significant association between the rs799917 CT SNP and the risk of gastric cancer (OR = 1.81, p = 0.001) and suggested that rs799917C interferes with the interaction between BRCA1 mRNA and miR-638, resulting in reduced BRCA1 expression [52]. A genome-wide association study (GWAS) has evaluated the association between microRNA (miRNA) expression levels and 327 miRNA-related SNPs and between SNPs and colon cancer risk [53]; the AA genotype of BRCA1 rs8176318 SNP was found to be associated with reduced miRNA expression and also with colon cancer risk (OR 1.31), suggesting that SNP rs8176318 induces thyroid cancer by altering the miRNA–mRNA target interactions. The association of rs8176318 with the risk of PTC may vary between ethnicities. Further investigations are needed to clarify the roles of these genes. BRCA1 may be associated with thyroid autoimmune disease and affect cancer development through this route. The expression of thyroid hormone receptor beta (TRβ) was significantly higher in breast cancer associated with BRCA1 compared to sporadic breast cancer, the latter not being linked to BRCA1 (p = 0.001) [54]. Furthermore, the expression of TRα alpha in cells with BRCA1 mutations was linked to a reduction in the 5-year survival rate.
This study has some limitations. First, it was not a randomized study with case-control matching. We adjusted for age and sex so that randomization would reduce any bias. Second, it was not based on whole-genome sequencing and was limited to the polymorphism of the BRCA genes, which ideally should not be considered independent of other genes due to the fact that BRCA1 and BRCA2 form stable complexes with partner proteins [43]. As techniques such as whole-genome sequencing improve and their cost drops, genetic analysis using next-generation sequencing should enable the identification of all genetic differences between patients with PTC and controls. Third, the relatively small sample size of both the experimental and control groups is a limitation of this study. Additionally, the number of controls is larger than the case group. This study involved blood collection and genetic testing, and obtaining consent from a large number of control group participants was limited due to cultural considerations in the Korean population. A large-scale study is necessary through multicenter research in the future. Fourth, additional SNPs besides BRCA1 and BRCA2 should also be analyzed to evaluate the non-random association of each SNP due to the proximity of BRCA1 SNPs analyzed in this study [55].
Since there were few cases with recurrence, we could not analyze the genetic features related to recurrence. Future research is needed to identify genes associated with thyroid cancer recurrence. Compared to well-established mutations, the interpretation of the direct causal relationship between polymorphisms and cancer requires caution. Further investigations should be performed to distinguish between association and direct causation. Nevertheless, many recent studies have demonstrated that SNPs are associated with cancer prognosis and can predict treatment response [56,57,58,59]. Although pathogenic variants of BRCA1 may have potentially caused thyroid cancer, it is possible to interpret that thyroid cancer was found in individuals with pathogenic BRCA1 mutation. Since SNPs include regions that are not translated into proteins, additional protein analysis is necessary for meaningful interpretation. Potential limitations of study evaluating the association of SNPs and cancer risk include false positive associations, difficulty of pinpointing the casual variant within a haplotype, and the possibility of overlooking rare variants, which may have more significant effects.
Further studies including miRNA, proteomic, and genomic biomarker analyses will enhance the comprehension of PTC etiopathogenesis [60].
5. Conclusions
Our findings suggest that the five BRCA1 SNPs (rs16940, rs799917, rs1799949, rs1799966, rs8176318) and haplotypes of BRCA1 contribute to the susceptibility of developing thyroid cancer. There was no association between BRCA2 and the risk of PTC. However, it is necessary to be cautious in interpreting the association between SNPs and the risk of thyroid cancer as evidence that SNP variations directly cause cancer. Further functional studies with a larger number of individuals, including diverse ethnicities and other genes related to the genetic pathway beyond BRCA, are needed to establish the role of BRCA1 in PTC.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ferlay J. Soerjomataram I. Dikshit R. Eser S. Mathers C. Rebelo M. Parkin D.M. Forman D. Bray F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in GLOBOCAN 2012 Int. J. Cancer 2015136 E 359E 38610.1002/ijc.2921025220842 · doi ↗ · pubmed ↗
- 2Roman B.R. Morris L.G. Davies L. The thyroid cancer epidemic, 2017 perspective Curr. Opin. Endocrinol. Diabetes Obes.20172433233610.1097/MED.000000000000035928692457 PMC 5864110 · doi ↗ · pubmed ↗
- 3Lortet-Tieulent J. Franceschi S. Dal Maso L. Vaccarella S. Thyroid cancer “epidemic” also occurs in low- and middle-income countries Int. J. Cancer 20191442082208710.1002/ijc.3188430242835 PMC 6587710 · doi ↗ · pubmed ↗
- 4Davies L. Morris L.G. Haymart M. Chen A.Y. Goldenberg D. Morris J. Ogilvie J.B. Terris D.J. Netterville J. Wong R.J. American Association of Clinical Endocrinologists and American College of Endocrinology Disease State Clinical Review: The Increasing Incidence of Thyroid Cancer Endocr. Pract.20152168669610.4158/EP 14466.DSCR 26135963 PMC 4923940 · doi ↗ · pubmed ↗
- 5Schonfeld S.J. Lee C. Berrington de Gonzalez A. Medical exposure to radiation and thyroid cancer Clin. Oncol.20112324425010.1016/j.clon.2011.01.15921296564 · doi ↗ · pubmed ↗
- 6Li Volsi V.A. Abrosimov A.A. Bogdanova T. Fadda G. Hunt J.L. Ito M. Rosai J. Thomas G.A. Williams E.D. The Chernobyl thyroid cancer experience: Pathology Clin. Oncol.20112326126710.1016/j.clon.2011.01.16021333507 · doi ↗ · pubmed ↗
- 7Zidane M. Cazier J.B. Chevillard S. Ory C. Schlumberger M. Dupuy C. Deleuze J.F. Boland A. Haddy N. Lesueur F. Genetic susceptibility to radiation-related differentiated thyroid cancers: A systematic review of literature Endocr. Relat. Cancer 201926 R 583R 59610.1530/ERC-19-032131476737 · doi ↗ · pubmed ↗
- 8Iglesias M.L. Schmidt A. Ghuzlan A.A. Lacroix L. Vathaire F. Chevillard S. Schlumberger M. Radiation exposure and thyroid cancer: A review Arch. Endocrinol. Metab.20176118018710.1590/2359-399700000025728225863 PMC 10118869 · doi ↗ · pubmed ↗