Shedding light on shade-avoidance: SlPIF8a plays a pivotal role in the tomato shade response
Róisín Fattorini

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
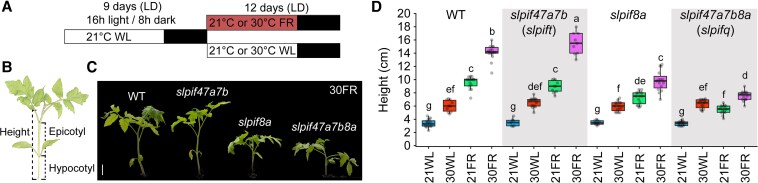 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsLight effects on plants · Greenhouse Technology and Climate Control · Postharvest Quality and Shelf Life Management
Sun-loving plants exhibit remarkable phenotypic plasticity through their shade-avoidance response. This response triggers morphological and physiological changes, such as stem and petiole elongation, that enable the plant to escape suboptimal light-limiting conditions and maximize exposure to the sunlight required for photosynthesis. In the shade, the proportion of far-red light increases and the ratio of red to far-red light (R/FR) decreases. Plant photoreceptors, primarily phytochrome B (phyB), sense shading from neighboring vegetation as a reduction in R/FR. In its active form, phyB inactivates and degrades the transcription factors known as PHYTOCHROME-INTERACTING FACTORS (PIFs) (Qiu et al. 2017). As the R/FR ratio falls below one, phyB is converted into its inactive form, releasing the PIFs from phyB-mediated repression. PIFs regulate multiple processes related to plant growth, including auxin synthesis (Hornitschek et al. 2012; Li et al. 2012; Casal 2013). While the role of PIFs has been well studied in Arabidopsis (A. thaliana), little is known about PIF functioning in crop plants despite the potential of PIFs, as growth regulators, to enhance agronomic traits.
In new work in The Plant Cell, Srinivas Kunta and colleagues (Kunta et al. 2025) found that SlPIF8a is a key regulator of the stem elongation response to low R/FR light in tomato (Solanum lycopersicum) seedlings. SlPIF candidate genes were selected based on high sequence similarity with known regulators in Arabidopsis, AtPIF4 and AtPIF7 (Burko et al. 2022). CRISPR-Cas9-generated slpif tomato mutants were grown under 1 of 2 light (white or low R/FR) and temperature (21 °C or 30 °C) conditions (Figure). Impaired stem elongation occurred under low R/FR conditions only in plants lacking the wild-type SlPIF8a gene (Figure). SlPIF8a functioning was corroborated through additional experiments, including the expression of SlPIF8a under its native promoter and a constitutive promoter in slpif8a or slpif quadruple (slpif47a7b8a) mutants, respectively, which led to a partial to nearly full rescue of stem elongation in the slpif mutants in low R/FR conditions. Comparisons between the slpif8a single mutant and higher order slpif double, triple, and quadruple mutants demonstrated that SlPIF4, SlPIF7a, and SlPIF7b also have minor roles in tomato stem elongation during the low R/FR light response.
RNA-seq data provided further insight into the dominant role of SlPIF8a. A comparison between SlPIF expression patterns in the aerial organs of tomato seedlings grown in white light revealed that SlPIF8a had the highest expression levels overall, particularly in the cotyledons and first leaf. The low R/FR response is thought to be initiated in these organs with the production of auxin, which is likely transported to the hypocotyl and epicotyl, where it promotes growth. High SlPIF8a transcript levels may lead to high protein expression, with SlPIF8a released from phyB-mediated degradation/inactivation upon exposure of the plant to low R/FR conditions. Therefore, the key role of SlPIF8a in the stem elongation response to low R/FR light may be, in part, due to its high expression in these organs.
The downstream mechanisms of the tomato low R/FR response were investigated through a series of elegant RNA-seq comparative analyses. A comparison was made between the genes differentially expressed in low R/FR and white light conditions among the wildtype, slpif8a single mutant, and slpif quadruple mutant (slpif47a7b8a) plants. This enabled the identification of genes regulated by both SlPIFs and low R/FR light, which included cell wall modifying genes and genes associated with gibberellin, auxin, and flavonoid biosynthesis. Interestingly, organ-specific expression data revealed that the hypocotyl and epicotyl elongation response mechanisms may be distinct. The response to low R/FR light was associated with the upregulation of cell cycle and cell division-associated genes exclusively in the epicotyl, whereas genes associated with cell elongation were enriched in the hypocotyl. An important next step is to characterize the developmental mechanisms underlying the epicotyl and hypocotyl elongation responses. Subsequently, determining the organ-specific roles of SlPIFs in regulating these elongation processes would be an exciting future research avenue.
In conclusion, Kunta et al. provide key insights into the genetic regulation of the stem elongation response to low R/FR light in tomato. The transcription factor SlPIF8a was identified as a key regulator of this process. Future studies could provide an evolutionary perspective by investigating whether SlPIF8a gene function is conserved within other Solanum species—a genus containing several important crops.
Recent related articles in the plant cell
Han et al. (2023) show that SALT OVERLY SENSITIVE2 (SOS2), a protein kinase essential for salt tolerance, positively regulates the shade avoidance response in Arabidopsis, suggesting a coordinated response of plants to salt stress and shade. Zhou et al. (2024) show that maize plants readjust their leaf azimuthal orientations in response to high planting density, maximizing canopy light interception. Kim et al. (2024) show that differences in the role of promoters, and the molecular activities of proteins, have contributed to functional diversification between PHYTOCHROME INTERACTING FACTOR 1 (PIF1) and PIF4 in Arabidopsis. The importance of each contributing factor varied depending on the light conditions.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Burko Y, Willige BC, Seluzicki A, Novak O, Ljung K, Chory J. PIF 7 is a master regulator of thermomorphogenesis in shade. Nat Commun. 2022:13(1):4942. 10.1038/s 41467-022-32585-636038577 PMC 9424238 · doi ↗ · pubmed ↗
- 2Casal JJ . Photoreceptor signaling networks in plant responses to shade. Annual Rev Plant Bio. 2013:64(1):403–427. 10.1146/annurev-arplant-050312-12022123373700 · doi ↗ · pubmed ↗
- 3Han R, Ma L, Lv Y, Qi L, Peng J, Li H, Zhou Y, Song P, Duan J, Li J, et al SALT OVERLY SENSITIVE 2 stabilizes phytochrome-interacting factors PIF 4 and PIF 5 to promote Arabidopsis shade avoidance. Plant Cell. 2023:35(8):2972–2996. 10.1093/plcell/koad 11937119311 PMC 10396385 · doi ↗ · pubmed ↗
- 4Hornitschek P, Kohnen MV, Lorrain S, Rougemont J, Ljung K, López-Vidriero I, Franco-Zorrilla JM, Solano R, Trevisan M, Pradervand S, et al Phytochrome interacting factors 4 and 5 control seedling growth in changing light conditions by directly controlling auxin signaling. Plant J. 2012:71(5):699–711. 10.1111/j.1365-313X.2012.05033.x 22536829 · doi ↗ · pubmed ↗
- 5Kim H, Lee N, Kim Y, Choi G. The phytochrome-interacting factor genes PIF 1 and PIF 4 are functionally diversified due to divergence of promoters and proteins. Plant Cell. 2024:36(8):2778–2797. 10.1093/plcell/koae 11038593049 PMC 11289632 · doi ↗ · pubmed ↗
- 6Kunta S, Dahan Y, Torgeman S, Chory J, Burko Y. Species-specific PIF utilization in the plant morphogenetic response to environmental stimuli. Plant Cell. 2025. 10.1093/plcell/koaf 048PMC 1207039640085779 · doi ↗ · pubmed ↗
- 7Li L, Ljung K, Breton G, Schmitz RJ, Pruneda-Paz J, Cowing-Zitron C, Cole BJ, Ivans LJ, Pedmale UV, Jung H-S, et al Linking photoreceptor excitation to changes in plant architecture. Genes Dev. 2012:26(8):785–790. 10.1101/gad.187849.11222508725 PMC 3337452 · doi ↗ · pubmed ↗
- 8Qiu Y, Pasoreck EK, Reddy AK, Nagatani A, Ma W, Chory J, Chen M. Mechanism of early light signaling by the carboxy-terminal output module of Arabidopsis phytochrome B. Nat Commun. 2017:8(1):1905. 10.1038/s 41467-017-02062-629199270 PMC 5712524 · doi ↗ · pubmed ↗