New species of Plesioaxymyia Sinclair (Diptera, Axymyiidae) from the Palaearctic Region, including an updated molecular phylogeny of the family

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
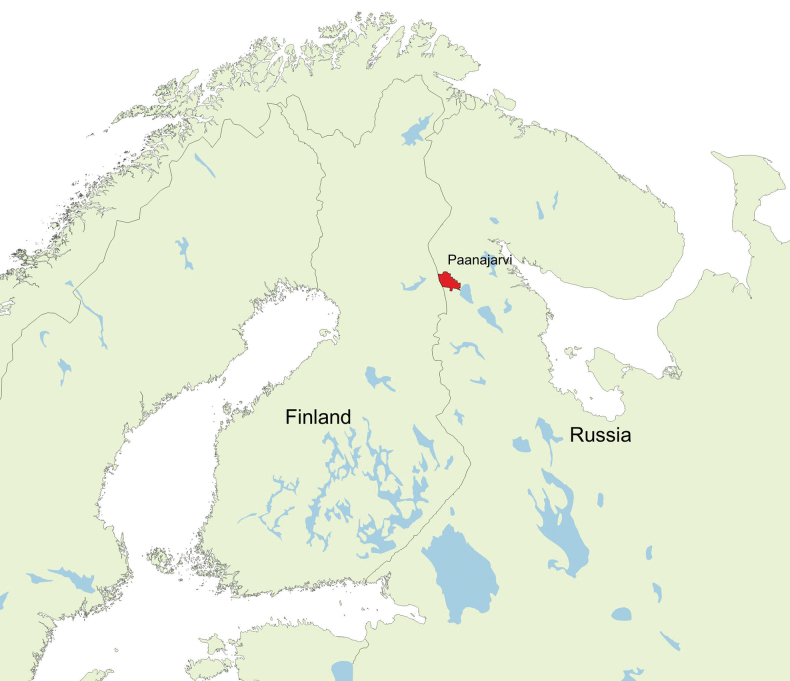 Figure 1
Figure 1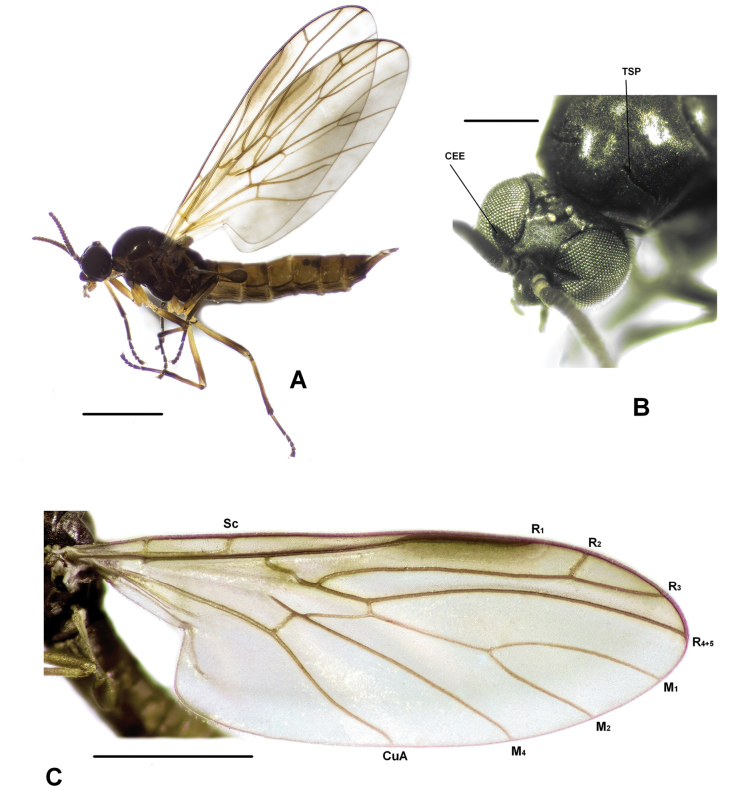 Figure 2
Figure 2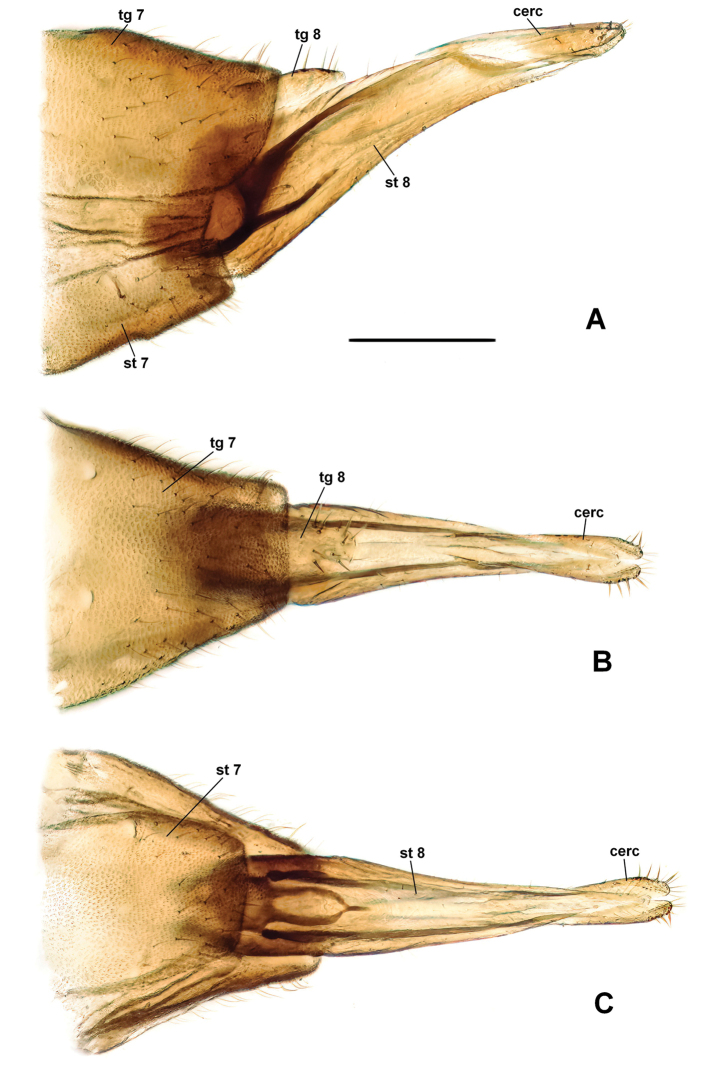 Figure 3
Figure 3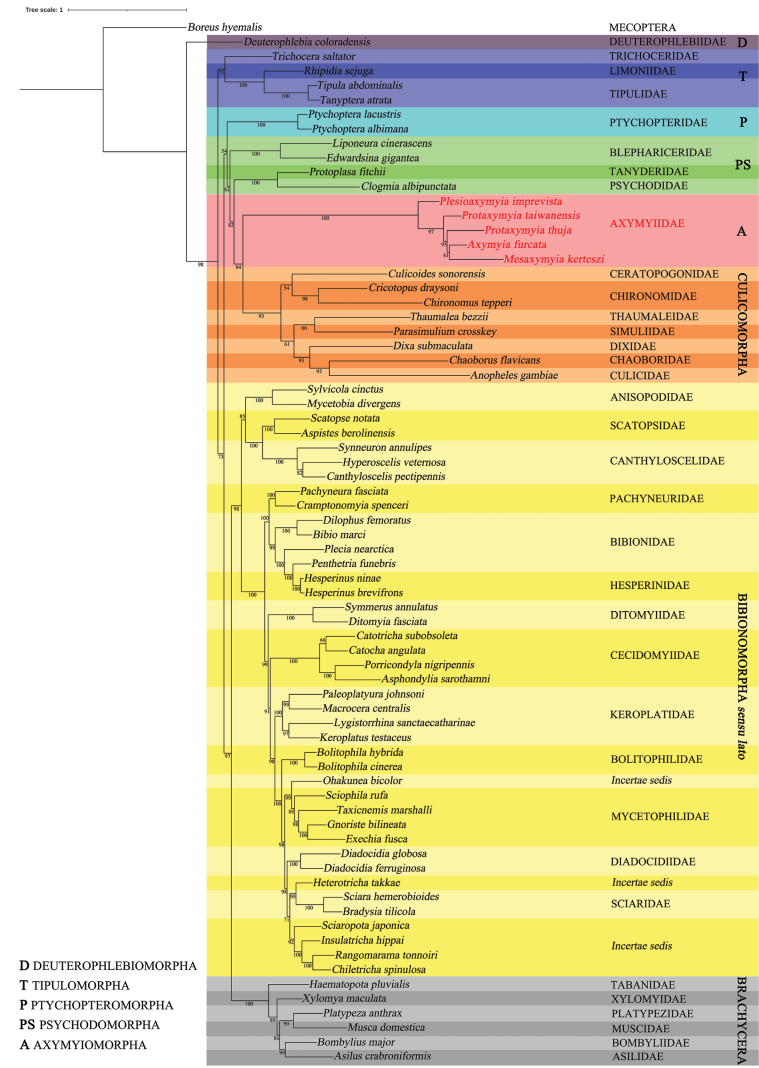 Figure 4
Figure 4 Figure 5
Figure 5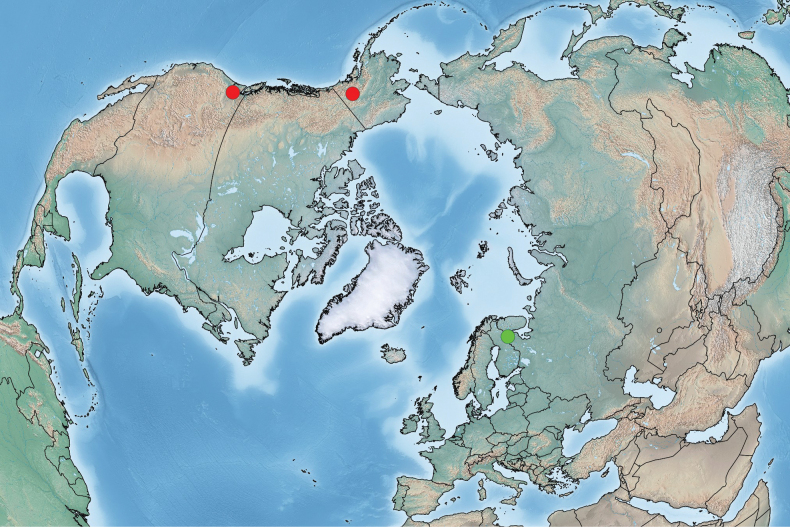 Figure 6
Figure 6| Gene fragment | Direction | Primer sequences (5´→ 3´) | Source |
|---|---|---|---|
| 12S | F | CTGGGATTAGATACCCTGTTAT |
|
| R | CAGAGAGTGACGGGCGATTTGT |
| |
| F | TACTATGTTACGACTTAT |
| |
| R | GCCAGCATTTGCGGTTATAC | M. Žurovcová lab., České Budějovice, Czech Republic | |
| 16S | F | TAATCCAACATCGAGGTC |
|
| R | CGAAGGTAGCATAATCAGTAG |
| |
| F | CGCCTGTTTATCAAAAACAT |
| |
| R | CCGGTCTGAACTCAGATCACGT |
| |
| 18S | F | AACCTGGTTGATCCTGCCAGT |
|
| R | TGATCCTTCTGCAGGTTCACCTACG |
| |
| F | AGATACCGCCCTAGTTCTAACC |
| |
| R | GGTTAGAACTAGGGCGGTATCT |
| |
| 28S | F | AGAGAGAGAGTTCAAGAGTACGTG |
|
| R | TAGTTCACCATCTTTCGGGTC |
| |
| F | ACCCGCTGAATTTAAGCAT |
| |
| COI | F | GGTCAACAAATCATAAAGATATTGG |
|
| R | TAAACTTCAGGGTGACCAAAAAATCA |
| |
| CAD | F | GGNGTNACNACNGCNTGYTTYGARCC |
|
| R | TTNGGNAGYTGNCCNCCCAT |
| |
| F | ACNGAYTAYGAYATGTGYGA |
| |
| R | TCRTTNTTYTTWGCRATYAAYTGCAT |
|
| 1 | Sternite 8 forming somewhat protruding dorsoapical corner in lateral view, basal segment of cerci distinct ( |
|
| – | Sternite 8 with smoothly rounded dorsoapical margin in lateral view, basal segment of cerci completely reduced (Fig. |
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsDiptera species taxonomy and behavior · Insect symbiosis and bacterial influences · Environmental DNA in Biodiversity Studies
Introduction
The family Axymyiidae is often referred to as “enigmatic” in the literature (Wihlm and Courtney 2011; Blagoderov and Lukashevich 2013; Fitzgerald and Wood 2014). This small group of flies includes four extant genera and nine species known in the Holarctic and Oriental regions (Fitzgerald and Wood 2014). Additionally, eight species in four extinct genera have been described from the Jurassic or Early Cretaceous deposits of Asia (Blagoderov and Lukashevich 2013; Shi et al. 2013). Only one species, Mesaxymyiakerteszi (Duda, 1930), is so far known from Europe. This is an extremely rare taxon, with a few recent records from eastern Slovakia (Martinovský and Roháček 1993) and Northwest Russia (Polevoi et al. 2018).
As it is known so far, all immature records of Axymyiidae are associated with dead wood. Extremely specialized larvae live inside very wet, often submerged, logs of various tree species (Wood 1981; Krivosheina 2000). The systematic position of this family is still uncertain (see Blagoderov and Lukashevich 2013; Sinclair 2013; Fitzgerald and Wood 2014 for reviews). It is often placed in or as a sister group to Bibionomorpha, but close relationships with this infraorder are not always supported by reconstructions, based on molecular data. In the comprehensive study by Wiegmann et al. (2011), Axymyiidae were placed within Bibionomorpha sensu lato, as a sister group to Bibionomorpha sensu stricto, whereas in other studies this family was represented as a sister group to Culicomorpha (Bertone et al. 2008; Ševčík et al. 2016) or in an unresolved polytomy (Zhang et al. 2023).
During studies of the Diptera fauna in Paanajarvi National Park (Russia, Karelia), an unusual looking female specimen was collected, evidently belonging to the family Axymyiidae. Preliminary examination showed that it was not a representative of any previously known Palaearctic genus, but accords well with the genus Plesioaxymyia Sinclair, which includes one North American species, P.vespertina Sinclair, 2013. Being morphologically very similar, the Karelian specimen differs from P.vespertina by characters of its terminalia. Considering the general rarity of the family Axymyiidae and the evident importance of the new record we feel it necessary to describe the new species and not wait for additional material. Moreover, we could not miss an opportunity to re-evaluate the phylogenetic position of Plesioaxymyia and Axymyiidae as a whole, based on new molecular data for extant species.
Material and methods
The type specimen was collected in Paanajarvi National Park, located in the northwestern part of the Republic of Karelia, Russia (Fig. 1). The park was established in 1992, and presently occupies a territory of over 104,000 ha, mostly covered with natural coniferous forests (Bizhon and Systra 1996; Timofeeva and Kutenkov 2009). In 2021, we used three Malaise traps as part of an insect inventory program in the territory of the park. The Plesioaxymyia specimen was collected with a trap set in a patch of spruce- and pine-dominated forest of Vacciniummyrtillus type, in the vicinity of the abandoned village of Vartolambina. The operation period of the trap was from June 1–27, which corresponds to late spring. Willows and some early-season flowers (e.g., Tussilago) were in blossom, and remnants of snow still could be found in shady places.
Location of Paanajarvi National Park, Karelia, Russia.
The trap residue was initially kept in 70% alcohol, and one Axymyiidae specimen was recognized during a preliminary inspection in the laboratory. For a detailed study, the specimen was dried by xylol and amyl acetate baths (Achterberg 2009). Terminalia were detached and macerated in KOH for 24 hours, then neutralized in acetic acid, washed in 70% alcohol, and transferred to glycerine. Finally, terminalia were placed in glycerine vial and pinned together with the rest of the specimen. The holotype is stored in the collection of the Zoological institute, St. Petersburg, Russia (ZISP).
Images of the habitus and wing were taken with a Leica MZ 9.5 stereo microscope and those of the terminalia with a Leica DM1000 compound microscope, both supplied with a LOMO MC6.3 camera. Z-stacked image series were combined using Helicon Focus v. 8.2.0 software, and final plates prepared with GIMP. The morphological terminology follows Sinclair (2013). The distribution map was created using the online tool SimpleMappr, available at https://www.simplemappr.net/.
Molecular methods principally follow those described in Ševčík et al. (2016). A total of 72 terminal taxa are included in the dataset (Appendix 1). Most of the specimens used in this study were collected by Malaise traps during the years 2000–2022. Some sequences were taken from the GenBank database. The material used in the molecular study was stored in ethanol (70% to 96%) or pinned. For DNA extraction, we used a NucleoSpin Tissue Kit (Macherey-Nagel, Düren, Germany) following the manufacturer’s protocol. PCRs (total volume = 20 μl) were performed using specific primers for sequences of three mitochondrial (ribosomal 12S and 16S and protein-encoding COI) and three nuclear genes (ribosomal 18S and 28S, protein-encoding CAD). The primers used for PCR amplifications and sequencing are listed in Table 1.
All amplified products were purified using a Gel/PCR DNA Fragments Extraction Kit (Geneaid, New Taipei City, Taiwan) following the manufacturer’s protocol. Purified products were sequenced by Macrogen Europe (Netherlands) or Eurofins Genomics (Germany). The sequences were assembled and edited in Sequencher v. 5.0 (Gene Codes Corporation, Ann Arbor, MI, USA) or SeqTrace v. 0.9.0 (Stucky 2012). GenBank accession numbers for all the sequences are listed in the Appendix 1. All sequences were checked with the NCBI database using the BLAST and in single-gene trees to avoid possible contamination or other inappropriate results.
Alignments of all genes were created using MAFFT v. 7 on the MAFFT server (http://mafft.cbrc.jp/alignment/server/). The resulting alignments were visually inspected and manually refined in BioEdit v. 7.2.5 (Hall 1999) when necessary. All unreliably aligned regions of rRNA genes were removed in the program GBLOCKS v. 0.91b (Castresana 2000); with conditions set as follows: allow smaller blocks, allow gap positions within the final blocks, allow less strict flanking positions and do not allow many contiguous non-conserved positions, or in ClipKIT v. 2.2.2 (Steenwyk et al. 2020) using the -gappy option with a threshold of 0.7. The third positions of COI gene were excluded from the subsequent analyses using software DAMBE (Xia and Xie 2001). The alignments were concatenated using FASconCAT v. 1.0 (Kück and Longo 2014). The final data matrix consisted of 5419 characters: 12S – 301 bp, 16S – 287 bp, 18S – 1985 bp, 28S – 1005 bp, COI – 436 bp (third positions removed) and CAD – 1405 bp.
The final concatenated dataset was partitioned by gene and codon position and subsequently analysed using the maximum likelihood (ML) method. Analyses were conducted using IQ-TREE v. 1 (Nguyen et al. 2015) on the IQ-TREE web server (Trifinopoulos et al. 2016). Best-fitting substitution models were chosen automatically by the IQ-TREE software: 12S – TVM+F+I+G4; 16S – TVM+F+I+G4; 18S – GTR+F+I+G4; 28S – TVM+F+I+G4; CAD_1 – TN+F+I+G4; CAD_2 – SYM+I+G4; CAD_3 – GTR+F+I+G4; COI_1 – TIM2+F+I+G4; COI_2 – TIM3+F+I+G4; without free-rate heterogeneity. Branch supports were evaluated using 1000 ultrafast bootstrap (Hoang et al. 2018). All other settings were left as default. The node support values are given in the form of ultrafast bootstrap (= ufboot). The resulting phylogenetic tree (consensus trees) was visualized using the Interactive Tree of Life (iTOL v. 7.0; Letunic and Bork 2024). A species of Mecoptera, Boreushyemalis (Linnaeus, 1767), was used as a root.
Results
Family Axymyiidae
Plesioaxymyia
imprevista
Taxon classificationAnimaliaDipteraAxymyiidae
Polevoi sp. nov.
FA16C5BE-26EC-510C-BC4A-857827D0D5E8
https://zoobank.org/F101EC9C-CA50-40E2-ABA1-B631CAD5F331
Specimens examined.
Holotype. Russia • ♀; Karelia, Paanajarvi National Park, Vartolambina; 66.246°N, 30.555°E; 80 m a.s.l.; 1–27 Jun. 2021; A. Protasova leg.; Malaise trap; GenBank: accession numbers PV036313, PV036316, PV036319, PV036317, PV035246, PV037681; ZISP, INS_DIP_0001011.
Differential diagnosis.
Medium-sized, dark brown species (Fig. 2A); wings hyaline with dark elongated pterostigma, covering apical half of vein R_1_; legs yellowish-brown with darkened tarsi, femora and tibiae darkened apically. Similar to Plesioaxymyiavespertina, from which it is distinguished by details of female terminalia: sternite 8 with smoothly rounded dorsoapical margin (forming somewhat protruding dorsoapical corner in P.vespertina) and completely reduced basal segment of cerci (distinct in P.vespertina).
Plesioaxymyiaimprevista sp. nov., female holotype A habitus, lateral view B head and thorax, oblique anterodorsal view C wing. Abbreviations: CEE – triangular excision of compound eye; CuA – anterior branch of cubital vein; M1,2,4 – medial veins; R2,3,4+5 – radial vein; Sc – subcostal vein; TSP – transverse shiny patch of mesonotum. Scale bars: 1 mm (A, С); 0.3 mm (B).
Description.
Head. Head dark brown. Face sunken, clypeus convex, mouthparts brown. Palpus brown, 5-segmented, with short (almost hidden) first segment and broadened third segment. Compound eye densely covered with short ommatrichia; divided by fine groove into upper and lower hemispheres of unequal size and with deep triangular excision opposite antennal base (Fig. 2B). Three ocelli arranged in equilateral triangle, placed on elevated tubercle. Frons with a few brownish hairs above antennal bases and between ocellar triangle and compound eye. Posterior part of head with numerous brownish hairs. Antenna 16-segmented, brown; pedicel yellowish apically; middle flagellomeres about twice as wide as long.
Thorax. Mesonotum dark brown, thinly dusted, lacking any larger setae but covered with tiny yellowish hairs; a pair of narrow transverse shiny patches present along prescutal suture (Fig. 2B); prescutum yellowish laterally. Scutellum yellowish-brown, strongly convex, with short hairs along posterior margin. Pleura brown.
Wing. Wing length 4.08 mm. Wing hyaline with light brownish tinge (Fig. 2C); brown elongated pterostigma occupies apical half of R_1_. Costa hardly produced beyond R_4+5_. Sc curved into costa proximally to Rs; Sc-r reduced. Rs with kink; R_2+3_ forked well beyond apex of R_1_, R_2_ deviates in slightly obtuse angle. Crossvein r-m perpendicular to R_4+5_. M_1+2_ branching slightly before apex of R_1_; section of M-stem distal to r-m about as long as M_1_, longer than M_2_ and about twice as long as its section proximal to r-m. M_4_ straight; CuA distinctly sinuous. CuP short, scarcely reaching beyond posteromedial angle of wing. Anal lobe well-developed. Macrotrichia on wing veins not visible. Halter brown.
Legs. Coxa, trochanters, femora and tibiae yellowish-brown; all femora and tibiae darkened apically; tarsi brownish. Hind tibia slightly curved in middle, with brush of bristly hairs posteroapically. Ratio of basitarsus to tibia: bt1:t1 – 0.45, bt2:t2 – 0.43, bt3:t3 – 0.30. Tibial spurs not developed.
Abdomen. Abdomen brown. Tergites 1–7 with sparse hairs posteriorly. Terminalia (Fig. 3) brown. Tergite 8 approximately one-third length of sternite 8, rounded and setose apically; sternite 8 lengthened, tapered apically, with sparse short hairs. Cercus one-segmented; basal segment not developed; apical segment about 4 times longer than wide, bearing ca. 10 setae.
Plesioaxymyiaimprevista sp. nov., female terminalia A lateral view B dorsal view C ventral view. Abbreviations: cerc—cercus; st 7, 8—sternites; tg 7, 8—tergites. Scale bars: 0.2 mm (A–C).
Etymology.
The species epithet is from the Latin imprevistus (unexpected, unforeseen), stressing that the finding of this species in Northwest Russia was a real surprise.
Distribution.
The species is currently known only from the type locality in Russian Karelia (Northwest Russia).
Biology.
The adult was collected with a Malaise trap set in Vacciniummyrtillus type pine- and spruce-dominated forest. The larval biology is unknown.
Key to the Holarctic species of Plesioaxymyia, females
**: **
Molecular data
The family Axymyiidae, represented in this dataset by five species from all four extant genera, is recovered as monophyletic with maximum support (ufboot = 100) (see Fig. 4). The new species described in this paper, Plesioaxymyiaimprevista sp. nov., represents the sister taxon to all other species of Axymyiidae included in the dataset. Surprisingly, two species of Protaxymyia are not recovered as sister taxa. Instead, Protaxymyiathuja Fitzgerald & Wood, 2014 is sister species (ufboot = 95) to the branch including Axymyiafurcata McAtee, 1921 and Mesaxymyiakerteszi. The sister relationship of the latter two species is, however, weakly supported (ufboot = 61).
Maximum likelihood hypothesis (IQ-TREE) for relationships among selected taxa of lower Diptera based on DNA sequence data /12S, 16S, 28S, 18S, COI (third positions removed) and CAD/, 5419 characters. Support numbers refer to ultrafast bootstrap values (ufboot) over 50.
The closest relative of the family Axymyiidae appears to be the infraorder Culicomorpha, although their sister relationship is only moderately supported (ufboot = 84). The well-supported clade (Tanyderidae + Psychodidae), represented by Protoplasafitchii Osten Sacken, 1859 and Clogmiaalbipunctata (Williston, 1893), forms a sister group to the clade (Axymyiidae + Culicomorpha), also with moderate support (ufboot = 82).
The infraorder Bibionomorpha sensu stricto is shown to be monophyletic with high support (ufboot = 100), as well as Bibionomorpha sensu lato (ufboot = 98), including also Anisopodidae, Canthyloscelidae and Scatopsidae. The sister clade to Bibionomorpha is the infraorder Brachycera, altogether forming the well-supported group Neodiptera (ufboot = 97), which is sister clade to the large group of aquatic or semi-aquatic lower Diptera formed by the families Ptychopteridae, Blephariceridae, Tanyderidae, Psychodidae, and Axymyiidae and the families of the infraorder Culicomorpha.
Discussion
Biology and distribution
Some species of the family Axymyiidae can be rather abundant in suitable places, at least in respect of the density of larvae (Krivosheina 2000; Wihlm and Courtney 2011). However, representatives of the genus Plesioaxymyia apparently are not among them. The Nearctic P.vespertina was searched for repeatedly in the location where it was first found (Sinclair 2013). It was encountered again only 50 years later, and far to the south of the earlier known locality. The Paanajarvi area in Russian Karelia has been known for its relatively well-studied entomofauna since the middle of 20^th^ century (Viramo 1998; Yakovlev et al. 2000), but during this time no Axymyiidae came into any entomologist’s view. All known records of Plesioaxymyia, both in North America and Russian Karelia, look to be accidental, which may be the result of extremely small populations or a cryptic lifestyle.
Adults of Axymyiidae usually can be found in shady habitats, near rivers, rivulets or other water bodies, close to woody substrates where the larvae develop (Krivosheina 2000; Wihlm and Courtney 2011). The collecting localities of P.vespertina in North America well meet these conditions (Sinclair 2013); however, P.imprevista sp. nov. was found in a patch of relatively dry coniferous forest (Fig. 5). The patch is bordered from the north by Cladonia type pine stands, and the only available water body around is the Olanga River, about 200 m to the south. Considering unsuccessful attempts to collect immature stages at the Alaskan and Mt. Rainier localities, Sinclair (2013) suggested that the larva of P.vespertina may reside in wet litter or decomposing fungi. In Paanajarvi, potentially suitable dead wood sources were totally absent close to the trap location. The distance from the river, however, does not look unreachable (even if Axymyiidae are considered weak fliers), so the wet dead wood, potentially available along the riverbank, cannot be excluded as a larval substrate. In general, the biology of Plesioaxymyia remains almost unknown. For now, we only can outline the flight activity period and preferred habitats, while other details of the life history are yet to be discovered.
Collecting biotope of Plesioaxymyiaimprevista sp. nov. A position of Malaise trap (yellow circle), displayed on a satellite image (https://www.bing.com/maps) B general view of the biotope.
The current range of Plesioaxymyia comprises the western Nearctic and western Palaearctic (Fig. 6). It is highly likely that the current disjunct pattern is just a result of insufficient knowledge. Considering the occurrence of the genus at relatively high latitudes (or high altitudes in more southern regions), a circumpolar, possibly boreo-alpine, distribution can be suggested, and further findings from at least the northern regions of East Russia can be expected.
Distribution of the genus Plesioaxymyia in the Holarctic Region. Plesioaxymyiavespertina is indicated by red circles and P.imprevista sp. nov. by a green circle.
Molecular phylogeny
The position of Axymyiidae as a sister group of Culicomorpha was initially revealed by Bertone et al. (2008), whose reconstruction was based on four nuclear gene markers and one sequenced taxon, namely Axymyiafurcata. It was rendered together with Nymphomyiadolichopeza Courtney, 1994 (Nymphomyiidae) with relatively low branch support (56%). The authors themselves considered this placement to be ambiguous, suggesting that it may have been influenced by the long-branch attraction effect. The subsequent study by Ševčík et al. (2016), which was based on six gene markers, focused on Bibionomorpha. However, it also included two species of Axymyiidae (Axymyiafurcata and Protaxymyiathuja) among numerous taxa in the broad outgroup. In this study, the sister group relation to Culicomorpha was identified again, though with similarly low support. The recent reconstruction, based on the complete mitochondrial genome (Zhang et al. 2023), included one taxon (Protaxymyia sp.) and did not demonstrate any clear relations for Axymyiidae, whose position appeared in an unresolved polytomy.
Sinclair (2013) proposed that Plesioaxymyia is sister group to the remaining Axymyiidae based on morphological characters. This hypothesis is now corroborated by genetic data. However, the grouping of other species deviates from expectations. Notably, Protaxymyiathuja is not rendered as sister taxon to P.taiwanensis Papp, 2007, being placed closer to species described in different genera (Axymyiafurcata and Mesaxymyiakerteszi).
The distinction between the genera of Axymyiidae (except the very peculiar Plesioaxymyia) is not clearly defined. Separation of adults largely relies on wing characters (Krivosheina 2000; Zhang 2010; Shi et al. 2013); however, with the accumulation of new materials, it has become evident that wing venation is not stable and its significance in the generic classification of Axymyiidae, including extinct genera, should be re-evaluated (Martinovský and Roháček 1993; Blagoderov and Lukashevich 2013; Fitzgerald and Wood 2014). The importance of larval and pupal characters (Krivosheina 2000; Fitzgerald and Wood 2014) is similarly unclear, given that preimaginal stages of several species have not yet been discovered.
Conclusion
Our reconstruction of the molecular phylogeny is the first to incorporate representatives of all extant genera of Axymyiidae, although this still represents only approximately half of the known species. Axymyiids constitute a relatively isolated group, with the Culicomorpha families representing its closest evolutionary relatives. Both groups share an aquatic or semi-aquatic larval habitat. The sister group relation of Axymyiidae to Culicomorpha is in agreement with the findings of Bertone et al. (2008) and Ševčík et al. (2016) and is now evidenced with much better (ufboot = 84) support. While this may not yet be considered as strongly reliable, it is assumed that it could be even better supported if more taxa were included.
The current, unexpected placement of the species in relation to the Axymyiidae genera may be an artefact caused by the incomplete DNA data for Mesaxymyiakerteszi and Axymyiafurcata. However, new molecular data provide additional background for the necessity of a revised generic classification of Axymyiidae. It is likely that such a revision will not be possible until details of the morphology of preimaginal stages and, preferably, also genetic data on all extant species are available.
Supplementary Material
XML Treatment for Plesioaxymyia imprevista
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Achterberg K van (2009) Can Townes type Malaise traps be improved? Some recent developments.Entomologische Berichten 69: 129–135.
- 2Belshaw R Quicke DLJ (1997) A molecular phylogeny of the Aphidiinae (Hymenoptera: Braconidae).Molecular Phylogenetics and Evolution 7: 281–293. 10.1006/mpev.1996.04009187088 · doi ↗ · pubmed ↗
- 3Bertone MA Courtney GW Wiegmann BM (2008) Phylogenetics and temporal diversification of the earliest true flies (Insecta: Diptera) based on multiple nuclear genes.Systematic Entomology 33: 668–687. 10.1111/j.1365-3113.2008.00437.x · doi ↗
- 4Bizhon AV Systra YJ (1996) The Paanajarvi National Park – a remarkable piece of the world natural heritage.Oulanka Reports 16: 11–16.
- 5Blagoderov VA Lukashevich ED (2013) New Axymyiidae (Insecta: Diptera) from the Mesozoic of East Siberia.Polish Journal of Entomology / Polskie Pismo Entomologiczne 82: 257–271. 10.2478/v 10200-012-0040-9 · doi ↗
- 6Campbell BC Steffen‐Campbell JD Sorensen JT Gill RJ (1995) Paraphyly of Homoptera and Auchenorrhyncha inferred from 18S r DNA nucleotide sequences.Systematic Entomology 20: 175–194. 10.1111/j.1365-3113.1995.tb 00090.x · doi ↗
- 7Castresana J (2000) Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis.Molecular Biology and Evolution 17: 540–552. 10.1093/oxfordjournals.molbev.a 02633410742046 · doi ↗ · pubmed ↗
- 8Dayrat B Tillier A Lecointre G Tillier S (2001) New clades of Euthyneuran Gastropods (Mollusca) from 28S r RNA Sequences.Molecular Phylogenetics and Evolution 19: 225–235.10.1006/mpev.2001.092611341805 · doi ↗ · pubmed ↗