S100PBP interacts with nucleoporin TPR and facilitates XY crossover formation in mice
Yufan Wu, Yang Li, Huan Zhang, Jingwei Ye, Ming Li, Jianteng Zhou, Xuefeng Xie, Hao Yin, Min Chen, Gang Yang, Suixing Fan, Baolu Shi, Hanwei Jiang, Qinghua Shi, Hui Ma

TL;DR
This study identifies S100PBP as a protein that helps form crossovers between X and Y chromosomes in mice and interacts with nuclear pore components during meiosis.
Contribution
The study reveals S100PBP's dual role in promoting XY crossovers and limiting autosomal crossovers through its interaction with nucleoporin TPR.
Findings
S100PBP is essential for XY crossover formation and male fertility in mice.
S100PBP interacts with nucleoporin TPR and localizes to nuclear pores in meiocytes.
Disruption of S100PBP increases autosomal recombination intermediates and crossovers.
Abstract
During meiosis, at least one crossover is selectively generated per pair of homologous chromosomes through homologous recombination to ensure their faithful segregation. The molecular mechanisms controlling meiotic recombination, particularly in XY chromosomes that share a tiny region of homology (i.e., the pseudoautosomal region, PAR), remain poorly understood. Here, we identify S100PBP as a key modulator of both XY and autosomal recombination in mice. S100pbp-knockout mice exhibit male infertility and spermatogenesis arrest at meiotic metaphase I, resulting from a drastic reduction in XY crossovers. This failure in XY crossover formation is due to a reduction in TEX11/M1AP-bound recombination intermediates at the PAR. By contrast, disruption of S100PBP significantly increases the number of recombination intermediates and crossovers on autosomes. Co-immunoprecipitation mass…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
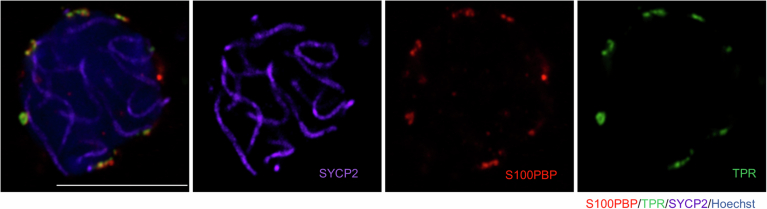 Figure 10
Figure 10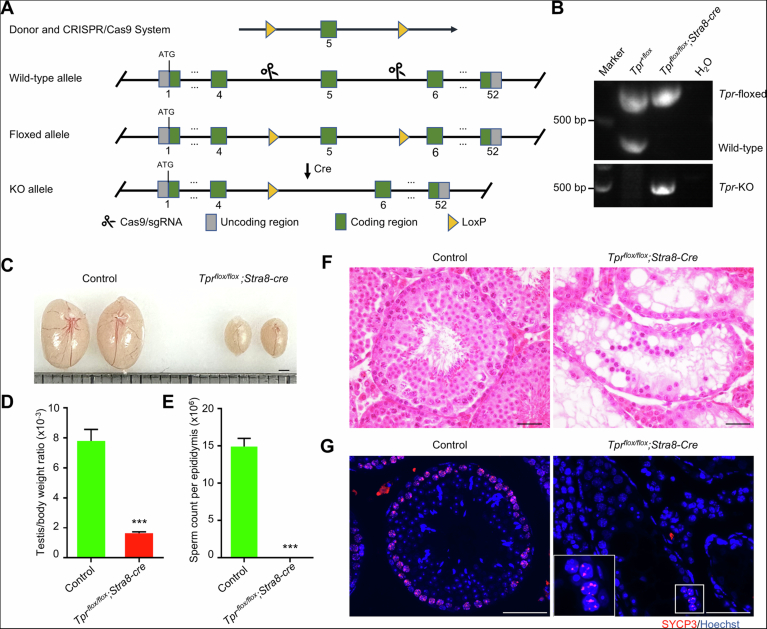 Figure 11
Figure 11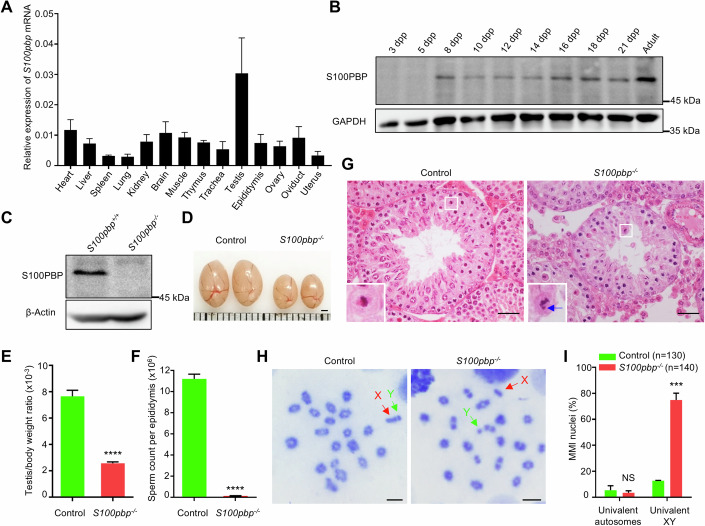 Figure 1
Figure 1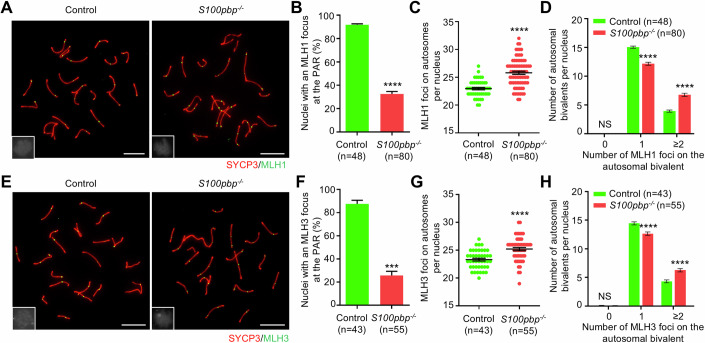 Figure 2
Figure 2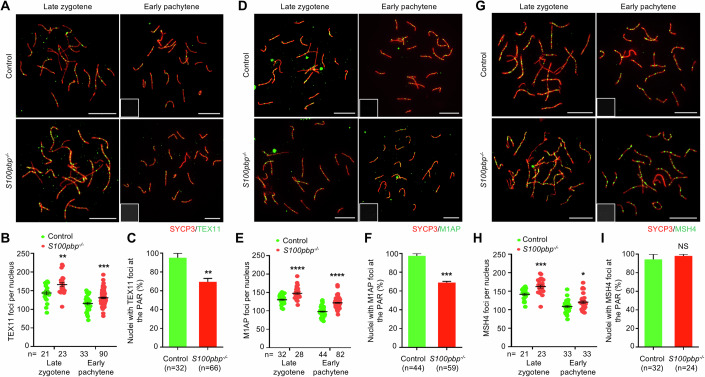 Figure 3
Figure 3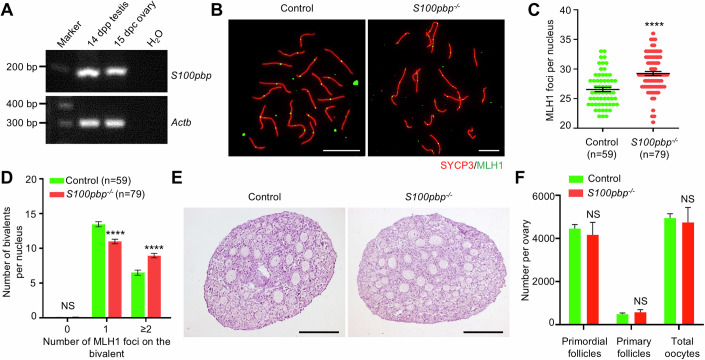 Figure 4
Figure 4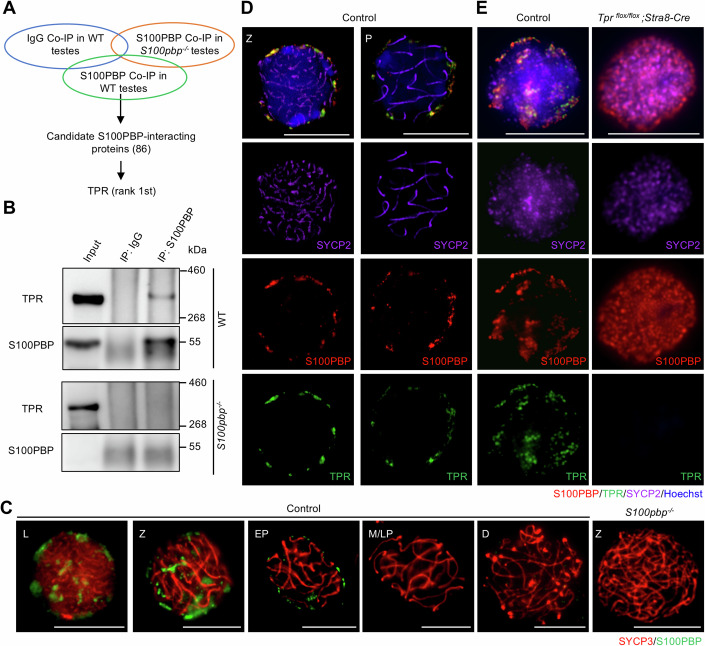 Figure 5
Figure 5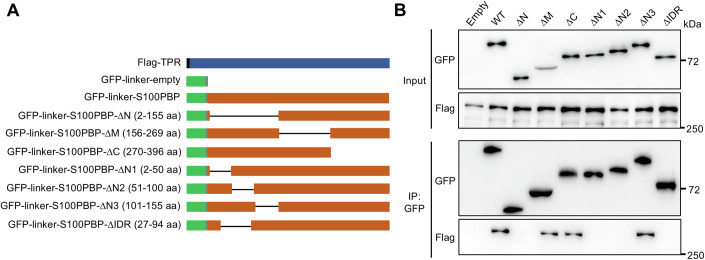 Figure 6
Figure 6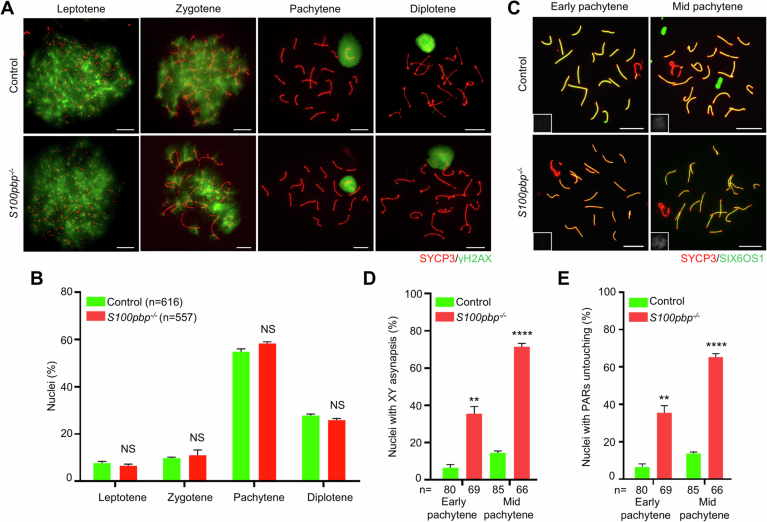 Figure 7
Figure 7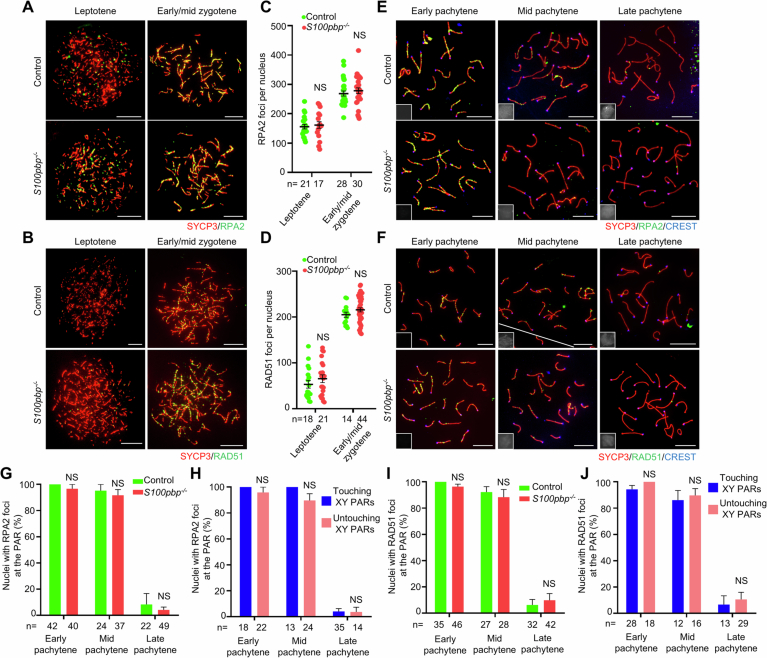 Figure 8
Figure 8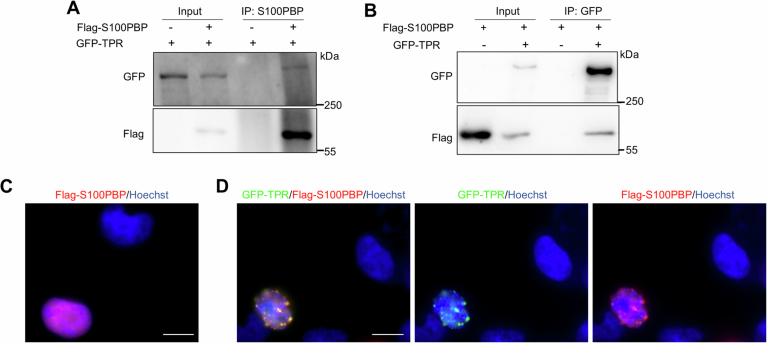 Figure 9
Figure 9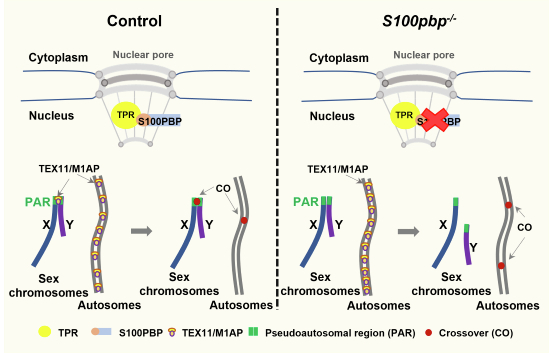 Figure 12
Figure 12 Figure 13
Figure 13- —http://dx.doi.org/10.13039/501100001809MOST | National Natural Science Foundation of China (NSFC)
- —http://dx.doi.org/10.13039/501100012166MOST | National Key Research and Development Program of China (NKPs)
- —Global Select Project of the Institute of Health and Medicine, Hefei Comprehensive National Science Center
- —Joint Fund for New Medicine of USTC
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsDNA Repair Mechanisms · RNA Research and Splicing · Genomics and Chromatin Dynamics
Introduction
In meiosis, the generation of crossovers between homologous chromosomes is one of the key events as it is essential for ensuring the proper alignment and segregation of homologous chromosomes at meiotic metaphase I (MMI) (Baudat et al, 2013; Hunter, 2015). Crossovers are initiated by the generation of programmed double-strand breaks (DSBs) during the leptotene and zygotene stages. The DSB ends are then resected to produce 3’ single-stranded DNA (ssDNA) overhangs protected by replication protein A (RPA) proteins, which are later replaced by the recombinase RecA homologs, RAD51 and DMC1 recombinases (Brown and Bishop, 2014; Hinch et al, 2020; Ribeiro et al, 2016). The recombinase-coated ssDNA filaments can recognize and invade the homologous chromosomes to form recombination intermediates which promote the alignment and synapsis of the homologous chromosomes. These recombination intermediates can be stabilized by a cluster of conserved proteins known as the ZMM complex (comprising Zip1-4, Msh4/Msh5, Mer3, and Spo16, initially identified in budding yeast), form double-Holliday junctions, and finally are resolved into crossovers by the endonuclease activity of the MutL homolog heterodimer MLH1-MLH3 (MutLγ) (Cannavo et al, 2020; Dai et al, 2021; Hunter, 2015). If the intermediates fail to be stabilized, they will be processed to generate non-crossover products.
The process of meiotic crossovers is strictly controlled such that crossovers are widely spaced (crossover interference), each homolog pair receives at least one crossover (crossover assurance), and the average number of crossovers per cell is well-controlled (at around 24 per spermatocyte in wild-type mice) (Otto and Payseur, 2019; Wang et al, 2015). Failure in crossover formation would result in infertility due to meiotic arrest or aneuploidy (Handel and Schimenti, 2010). For example, disruption of components in the mouse ZMM subcomplexes, including SHOC1-TEX11-SPO16 (also known as ZZS complex named after the yeast homologs Zip2-Zip4-Spo16) and MSH4-MSH5 (MutSγ), resulted in reduced or no crossover formation, along with incomplete synapsis of varying degrees (Adelman and Petrini, 2008; Santucci-Darmanin et al, 2002; Shinohara et al, 2008; Snowden et al, 2004; Yang et al, 2008). Recently, we found that loss of M1AP, a ZZS-interacting protein, also reduces MSH4 and TEX11 foci, leading to decreased crossovers (Li et al, 2023).
Compared with autosomes, recombination on XY chromosomes is more dedicatedly controlled to assure the formation of a crossover within a tiny homologous segment known as the pseudoautosomal region (PAR). It has been reported that the timing of programmed DSB formation and enzymes catalyzing meiotic DSBs at the PAR are different from those of autosomes in mice (Acquaviva et al, 2020; Boekhout et al, 2019; Brick et al, 2012; Kauppi et al, 2011; Kauppi et al, 2012; Lange et al, 2016; Papanikos et al, 2019). Recent findings on mutant mouse models have also shown that disruption of RAD51AP2, a RAD51-interacting protein, or ATF7IP2, a meiosis-specific SETDB1-interacting protein, specifically impairs DSB repair towards crossovers at the PAR, but not on autosomes (Alavattam et al, 2024; Ma et al, 2022; Shao et al, 2023), suggesting that the DSB repair mechanisms leading to crossover formation on XY chromosomes are also differently regulated from those on autosomes.
To unravel new functional genes in meiosis, we conducted in-depth mining of the published single-cell sequencing data of male germ cells and identified a substantial number of genes highly expressed in spermatocytes but without known functions in spermatogenesis. Among these genes, we noted that S100pbp, encoding an interacting partner of Ca^2+^-binding S100P protein (Dowen et al, 2005), is highly expressed in the early spermatocytes, implying a potential role in meiosis (Chen et al, 2018). S100PBP has been reported to inhibit the adhesion of pancreatic cancer cells through the S100PBP/CTSZ/RGD αvβ5 pathway (Lines et al, 2012; Lu et al, 2021) and plays a tumor suppressor role (Srivastava et al, 2023). However, whether it is required for spermatogenesis is unknown. In this study, we identify S100PBP as a novel and intriguing regulator of meiotic recombination, which promotes spermatogenesis by facilitating XY crossover formation while constraining autosomal crossover formation. We find that S100PBP localizes to the nuclear pores of meiocytes likely via the interaction with the nucleoporin TPR. Our study elucidates the physiological role of S100PBP and reveals potential regulatory mechanisms involving nuclear pores in mammalian recombination.
Results
S100pbp is essential for spermatogenesis
To investigate the potential role of S100pbp, we first examined its expression using reverse transcription-polymerase chain reaction (RT-PCR) and western blotting. The quantitative polymerase chain reaction (qPCR) assays revealed that S100pbp mRNA was detected in various organs examined of adult mice with a predominantly highest expression in testes (Fig. 1A). To confirm the expression of S100PBP protein in testis, we generated an anti-S100PBP antibody that recognizes an epitope consisting of amino acids 19–33 of the mouse protein. S100PBP protein was detected in mouse testes starting from 8 days postpartum (dpp) (Fig. 1B), suggesting a potential role in meiosis.Figure 1. Meiotic metaphase I arrest with a high prevalence of univalent XY chromosomes in S100pbp^−/−^ mice.(A) Quantitative real-time PCR showing the expression pattern of S100pbp in various tissues from adult wild-type mice. Data represent the mean ± SEM of three biological replicates obtained from three different mice for each tissue. (B) Western blot analysis of S100PBP expression in testes from mice at different days postpartum (dpp). GAPDH was used as the loading control. (C) Western blotting with testis lysates from control and S100pbp^−/−^ mice using the anti-S100PBP antibody. β-Actin was used as the loading control. (D) Representative images of testes from 8-week-old control and S100pbp^−/−^ mice. Each grid represents 1 mm. (E, F) The ratio of testis/body weight (E) and sperm count per epididymis (F) of 8-week-old control and S100pbp^−/−^ mice. The data are from at least three biological replicates and represent the mean ± SEM. ****P < 0.0001, two-tailed Student’s t test. (G) Testicular histology from 8-week-old control and S100pbp^−/−^ mice. The magnified view of the boxed area is shown in the lower-left corner of the image. The blue arrow indicates the unaligned chromosome in the representative metaphase cell. Scale bars, 50 μm. (H) Representative meiotic metaphase I (MMI) spermatocytes stained with Giemsa. Chromosomes X and Y are indicated. Scale bars, 10 μm. (I) Quantification of the frequencies of nuclei with univalent XY or autosomes in MMI spermatocytes shown in (H). Data represent the mean ± SEM from at least three biological replicates. n, the number of MMI cells scored. NS, not significant; ***P = 0.0002; two-tailed Student’s t test. Source data are available online for this figure.
Next, to explore the physiological function of S100PBP in vivo, we generated S100pbp-knockout mice (S100pbp^−/−^) using CRISPR/Cas9 technology. S100pbp^−/−^ mice carry a 2-base-pair deletion in exon 1 (c.79_80 del) of the S100pbp gene, which introduces a premature stop codon (Appendix Fig. S1). Western blotting detected the presence of full-length S100PBP in the wild-type testes, but not in S100pbp^−/−^ testes (Fig. 1C). It should be noted that the frameshift allele may produce a truncated protein (p.G27Ffs*4) lacking the C-terminal region or a truncated protein starting at an internal ATG, which, if present, may not be recognized by the anti-S100PBP antibody.
Both S100pbp^+/−^ and S100pbp^−/−^ mice show no obvious developmental abnormalities and appear healthy. S100pbp^+/−^ mice are fertile and capable of breeding normally. Detailed examination of S100pbp^+/−^ mice revealed no discernible defects in spermatogenesis. Specifically, there are no significant differences in testis size, testis/body weight ratio, or sperm number when compared to their wild-type littermates (Appendix Fig. S2). However, when we crossed the adult S100pbp^−/−^ male mice with wild-type females for a minimum of 3 months, no litters were produced, indicating that the S100pbp^−/−^ male mice are infertile (Appendix Table S1). S100pbp^−/−^ mice have smaller testes and a significantly reduced testes-to-body weight ratio when compared to those of the control littermates (7.66 ± 0.46 × 10^−3^ in controls versus 2.59 ± 0.10 × 10^−3^ in S100pbp knockouts; n = 4; P < 0.0001; unpaired t test; Fig. 1D,E). While over 10 million spermatozoa per epididymis were detected in the control mice, only a few spermatozoa were seen in the epididymis of S100pbp^−/−^ mice (Fig. 1F). Histological analyses of testes from the control mice revealed the presence of numerous spermatogenic cells of all developmental stages. In contrast, the seminiferous tubules of S100pbp^−/−^ mice contain many MMI spermatocytes with unaligned chromosomes, and the postmeiotic germ cells are markedly fewer than in the controls. (Fig. 1G). These results suggest that the homozygous frameshift mutation of S100pbp in mice causes male infertility and impairs spermatogenesis.
An increased prevalence of XY univalents in S100pbp−/− MMI spermatocytes
At meiotic metaphase I, chiasmata tether each pair of homologous chromosomes, which are referred to as the bivalents, ensuring their alignment at the metaphase plate and the subsequent accurate separation of homologous chromosomes into daughter cells (Baudat et al, 2013; Hunter, 2015). Achiasmatic chromosomes, which are referred to as univalents, would fail to align at the metaphase plate (Carpenter, 1994). Thus, we infer that the unaligned chromosomes observed in S100pbp^−/−^ MMI spermatocytes suggested the presence of univalents. To test this possibility, we prepared metaphase chromosome spreads from control and S100pbp^−/−^ testes. In control mice, the majority of MMI cells contain 20 bivalents, among which the XY bivalent has a typical appearance of “!” in morphology. However, XY chromosomes were found frequently separated in the mutant MMI cells (Fig. 1H). Analysis of metaphase I chromosomes revealed that, in contrast to 12.86% of MMI spermatocytes containing univalent XY in the control mice, 75.54% of MMI spermatocytes in S100pbp^−/−^ mice have univalent XY, while the frequency of univalent autosomes is similar between these two groups (5.42% in controls versus 3.52% in S100pbp knockouts) (Fig. 1I). Thus, based on these results, we believe that the unaligned chromosomes observed in S100pbp^−/−^ MMI spermatocytes are the univalent XY chromosomes.
S100pbp−/− spermatocytes have increased autosomal crossovers but decreased XY crossovers
The increased frequency of XY separation in S100pbp^−/−^ MMI spermatocytes indicates that crossover formation at the PAR is likely abrogated. To test this, we quantified the number of MLH1 foci, which mark sites of crossovers, in spread mid-pachytene spermatocytes. The results showed that the frequency of nuclei with an MLH1 focus at the PAR is significantly lower in S100pbp^−/−^ mice than in control mice (91.88% in controls and 32.65% in S100pbp knockouts; Fig. 2A,B), indicating decreased XY crossover formation. Intriguingly, we observed a significant increase of MLH1 foci on autosomes in S100pbp^−/−^ spermatocytes (25.81 ± 0.28) than in control mice (22.96 ± 0.21) (Fig. 2C) with more autosomal bivalents harboring 2 or 3 MLH1 foci per nucleus (6.79 in S100pbp knockouts versus 3.94 in controls, Fig. 2D). To confirm the intriguingly opposite alterations in crossover numbers between the XY and autosomes in S100pbp knockouts, we analyzed the staining of MLH3, which interacts with MLH1 to form MutLγ nuclease for crossover processing, at the mid-pachytene stage. Similar to the observations for MLH1, the frequency of nuclei with an MLH3 focus on the sex chromosomes in S100pbp^−/−^ spermatocytes is decreased (87.60% in controls and 25.78% in S100pbp knockouts; Fig. 2E,F), with more MLH3 foci on autosomes per nucleus (23.33 ± 0.25 in controls versus 25.22 ± 0.29 in S100pbp knockouts; Fig. 2G) and autosomal bivalents containing 2 or 3 MLH3 foci per nucleus (Fig. 2H).Figure 2S100pbp^−/−^ spermatocytes show a decrease in XY crossovers while exhibiting an increase in autosomal crossovers.(A, E) Representative mid-pachytene spread spermatocytes stained for SYCP3 (red) and MLH1 (green, A) or MLH3 (green, E). Miniaturized H1t staining (gray), shown in the lower-left corner of the overlay images, was used to identify mid-pachytene spermatocytes (H1t-moderate). Scale bars, 10 μm. (B, F) Frequencies of nuclei with an MLH1 focus (B) or an MLH3 focus (F) at the pseudoautosomal region (PAR) of XY chromosomes at the mid-pachytene stage. (C, G) The number of MLH1 foci (C) or MLH3 foci (G) on autosomes per nucleus. (D, H) Number of autosomal bivalents with 0, 1, and ≥2 MLH1 foci (D) or MLH3 foci (H) per nucleus. (B–D, F–H) Data represent the mean ± SEM from at least three biological replicates. n, the number of cells scored. NS, not significant; ***P = 0.0002; ****P < 0.0001; two-tailed Student’s t test. Source data are available online for this figure.
In addition, in S100pbp^−/−^ mice, typical spermatocytes of all four substages of meiotic prophase I (leptotene, zygotene, pachytene, and diplotene) were identified, and their frequencies are all comparable to those in control mice, indicating that the S100pbp^−/−^ spermatocytes progress normally through meiotic prophase I (Fig. EV1A,B). Similar to control cells, autosomes are fully synapsed and γH2AX staining is restricted to the XY body in the S100pbp^−/−^ pachytene cells (Fig. EV1A). Noticeably, there are significantly higher proportions of pachytene cells showing unsynapsed XY and untouching XY PARs in S100pbp mutant mice than those in control mice (Fig. EV1C–E).
These observations together indicate that S100PBP facilitates crossover formation on XY chromosomes, constrains excess crossovers formed on autosomes, and is dispensable for meiotic prophase progression.
S100pbp-disruption increases the number of genome-wide ZMM-bound recombination intermediates
Previous reports suggest that the elevated number of autosomal MLH1 foci, particularly the higher fractions of bivalents with ≥2 MLH1/MLH3 foci, might be associated with a reduction in crossover interference (Barchi et al, 2008; Girard et al, 2023; Zhang et al, 2014). To explore this, we measured the distances between adjacent MLH1 foci on autosomes with two or more foci, and found that the inter-focus distances in S100pbp^−/−^ mice are similar to those of the control mice. This indicates that crossover interference remains unchanged on autosomes of the S100pbp^−/−^ spermatocytes (Appendix Fig. S3A).
In addition, there is a well-established correlation between chromosome axis length and the number of crossovers (Wang et al, 2021). We compared the average axis length per nucleus in mid-pachytene S100pbp^−/−^ and control spermatocytes, which revealed no significant difference (Appendix Fig. S3B). These results suggest that the increased autosomal crossovers in S100pbp^−/−^ spermatocytes are not attributed to the chromosome axis length either.
Given that changes in crossover numbers could stem from alterations in DSB formation and/or repair processes, we next examined DSB formation by quantifying the numbers of RPA2 and RAD51, which bind the single-stranded DNA overhangs at the DSB sites, in spread spermatocytes. Our analysis revealed that the numbers of RPA2 and RAD51 foci are comparable between S100pbp^−/−^ spermatocytes and controls during the leptotene and zygotene stages (Fig. EV2A–D). This indicates that the generation of genomic DSBs remains unaffected by the disruption of S100PBP.
Since DSB formation is not affected, we inferred that the increased crossovers could arise during the formation and processing of recombination intermediates, at the expense of other types of repair. It has been reported that the two ZMM subcomplexes, SHOC1-SPO16-TEX11 (ZZS) and MSH4-MSH5 (MutSγ), bind to and stabilize the recombination intermediates, including the displacement loops (D-loops), single-end invasions (SEIs), and double-Holliday junctions (dHJs), to promote crossover formation (Pyatnitskaya et al, 2019). We thus quantified the dynamics of TEX11, M1AP (an interactor of the ZZS complex), and MSH4 foci on the surface spread of control and S100pbp^−/−^ spermatocytes (Fig. 3). In control mice, 143.95 ± 5.02 and 115.48 ± 3.48 TEX11 foci were observed in the late zygotene and early pachytene stages, respectively. We observed a similar trend for TEX11 foci in S100pbp^−/−^ spermatocytes, with significantly higher numbers at both the late zygotene (166.35 ± 5.44) and the early pachytene stages (130.87 ± 2.06) compared to controls (Fig. 3A,B). In addition, we noted an increase in M1AP foci, which facilitates TEX11 recruitment (Li et al, 2023), as well as MSH4 foci in S100pbp^−/−^ spermatocytes (Fig. 3D,E,G,H). This indicates that the S100pbp frameshift mutation leads to an increased number of crossover-designated recombination intermediates on the autosomes.Figure 3S100PBP deficiency results in decreased ZZS-bound recombination intermediates at the PAR but increased ZMM-bound recombination intermediates on the autosomes.(A, D, G) Immunofluorescence staining with antibodies against SYCP3 (red) and TEX11 (green, A), M1AP (green, D), or MSH4 (green, G), on spermatocyte spreads. Miniaturized H1t staining (gray), shown in the lower-left corner of the overlay images, was used to identify the early pachytene spermatocytes (H1t-negative). Scale bars, 10 μm. (B, E, H) Scatter plots showing the numbers of TEX11 foci (B), M1AP foci (E), or MSH4 foci (H), per nucleus in control and S100pbp^−/−^ spermatocytes at the indicated stages. (C, F, I) Frequencies of nuclei with TEX11 foci (C), M1AP foci (F), or MSH4 foci (I) detected at the PAR in spread early pachytene spermatocytes with touching XY PARs. (B–C, E, F, H, I) Data represent the mean ± SEM from at least three biological replicates. n, the number of cells scored. NS, not significant; **P = 0.0044 (B), ***P = 0.0002 (B), **P = 0.0070 (C), ****P < 0.0001 (E), ***P = 0.0006 (F), *P = 0.0104 (H), ***P = 0.0005 (H); two-tailed Student’s t test. Source data are available online for this figure.
Decreased number of ZZS-bound recombination intermediates at the PAR of S100pbp−/− spermatocytes
We next examined the generation and repair of DSBs at the PAR, the range of which is defined according to the measured axial length ratios as we previously reported (Ma et al, 2022). In S100pbp^−/−^ mice, the frequencies of nuclei with RPA2 or RAD51 foci at the PAR in early, mid, and late pachytene spermatocytes are all comparable to those in control mice (Fig. EV2E–J). Specifically, no significant differences were observed between nuclei with touching XY PARs and those with untouching XY PARs (Fig. EV2H,J). These results suggest that there are no detectable defects in the generation of PAR DSBs or delays in their repair, in S100pbp^−/−^ mice, regardless of whether the PAR are separated or in contact.
We further analyzed the abundance of recombination proteins at the PAR, focusing on early pachytene spermatocytes with touching XY PARs. We scored the presence of these recombination proteins as a marker of recombination intermediates, following our previous methodology (Ma et al, 2022). In contrast to the genome-wide increase in TEX11 and M1AP foci at the early pachytene stage, their frequencies at the PAR are significantly decreased (TEX11: 95.24 ± 4.76% in controls versus 69.58 ± 3.60% in S100pbp knockouts; M1AP: 97.44 ± 2.56% in controls versus 69.02 ± 1.23% in S100pbp knockouts; Fig. 3C,F). Intriguingly, the frequency of nuclei with MSH4 foci at the PAR does not differ from that in controls (Fig. 3I). These observations suggested that the recruitment of TEX11 and M1AP is impaired at the PAR, while MSH4 recruitment remains unaffected. Collectively, these findings indicate that S100pbp constrains the formation of recombination intermediates committed to crossovers on autosomes but facilitates the XY crossover formation likely by promoting the recruitment of the ZZS complex at the PAR.
S100pbp is dispensable for follicle development and female fertility
S100pbp mRNA was detected in fetal ovaries (Fig. 4A), suggesting that it may also play a role in the meiosis of females. To check whether ablation of S100PBP would lead to meiotic defects in oocytes, oocyte spreads from embryos at 17.5 days postcoitus (dpc) were prepared for analyzing meiotic recombination. Similar to observations in S100pbp^−/−^ spermatocytes, the number of MLH1 foci is increased in S100pbp^−/−^ oocytes (Fig. 4B,C), along with more bivalents containing 2, 3, or 4 MLH1 foci per nucleus (Fig. 4D), when compared with those in controls. Nevertheless, it appears that the increased crossover formation does not impede the follicle development in S100pbp^−/−^ mice. Analysis of the ovaries from mice at 6.5 dpp of age revealed that the ovary size and morphology of S100pbp^−/−^ females are similar to those of control mice (Fig. 4E). Ovarian histological analyses indicated no notable difference in the numbers of total oocytes, primordial follicles, and primary follicles between control and S100pbp^−/−^ mice (Fig. 4E,F). Moreover, we did not notice any overt defects in fertility in S100pbp^−/−^ female mice. Together, these findings indicate that S100PBP, though not essential for female fertility and follicle development, constrains meiotic recombination in oocytes, which is in congruent with its function in autosomal recombination in spermatocytes.Figure 4. The number of crossovers is increased while follicle development remains unaffected in S100pbp^−/−^ ovaries.(A) Reverse transcription-PCR (RT-PCR) analysis of S100pbp expression in fetal ovaries (15 days postcoitum (dpc)) and testes (14 dpp) from wild-type mice. Actb served as an internal control. (B) Immunofluorescence staining with antibodies against SYCP3 (red) and MLH1 (green) on oocyte spreads from control and S100pbp^−/−^ embryos (17.5 dpc). Scale bars, 10 μm. (C) Number of MLH1 foci per nucleus in control and S100pbp^−/−^ oocytes. Data represent the mean ± SEM from at least two biological replicates. n, the number of cells scored. ****P < 0.0001; two-tailed Student’s t test. (D) The number of bivalents with 0, 1, and ≥2 MLH1 foci per nucleus. Data represent the mean ± SEM from at least two biological replicates. n, the number of cells scored. NS, not significant; ****P < 0.0001; two-tailed Student’s t test. (E) Representative haemotoxylin-stained ovarian sections of 6.5-dpp-old control and S100pbp^−/−^ mice. Scale bars, 50 μm. (F) Follicle counts and total oocyte counts per ovary at 6.5 dpp. n, the number of cells scored from at least three biological replicates. NS, not significant; two-tailed Student’s t test. Source data are available online for this figure.
S100PBP interacts with TPR and localizes to the nuclear pores in a TPR-dependent manner
To explore the molecular mechanism by which S100PBP promotes meiotic recombination, we performed co-immunoprecipitation (IP) in testicular lysates from 10-week-old wild-type mice using the anti-S100PBP antibody, followed by mass spectrometry (MS) analysis (Fig. 5A). S100pbp^−/−^ mice at the same age were used as a parallel negative control. However, it is important to note that differences in cell composition between WT and mutant testes at this age may affect the significance of the observed differences in protein interactions.Figure 5S100PBP interacts with TPR and is localized to the nuclear pores of spermatocytes in a TPR-dependent manner.(A) Co-immunoprecipitation (co-IP) and mass spectrometry (MS) to identify the S100PBP-interacting proteins. Proteins, which were detected by IP with the anti-S100PBP antibody in the wild-type (WT) testes but absent in groups by anti-rabbit IgG antibody in the wild-type testes and anti-S100PBP antibody in the S100pbp^−/−^ testes, were considered as candidate S100PBP-interacting proteins. The number in the brackets indicates the number of proteins. TPR ranks first in the number of unique peptides. (B) Co-IP using anti-S100PBP antibody with whole-testis lysates of WT mice and S100pbp^−/−^ mice, followed by western blot analyses of S100PBP and TPR. (C) Immunofluorescence staining with antibodies against SYCP3 (red) and S100PBP (green) on spermatocyte smears of control and S100pbp^−/−^ testes. L leptotene, Z zygotene, EP early pachytene, M/LP mid/late pachytene, D diplotene. Scale bars, 10 μm. (D) Confocal imaging of zygotene and pachytene spermatocyte on spermatocyte smears of adult testes from WT mice after immunofluorescence staining of TPR (green), S100PBP (red), and SYCP2 (purple). The nuclei were counterstained with Hoechst 33342 (blue). Z zygotene, P pachytene. Scale bars, 10 μm. (E) Immunofluorescence staining with antibodies against TPR (green), S100PBP (red), and SYCP2 (purple) on spermatocyte smears of control and Tpr^flox/flox^; Stra8-cre testes. The nuclei were counterstained with Hoechst 33342 (blue). Scale bars, 10 μm. Source data are available online for this figure.
Among the 85 candidate S100PBP-interacting proteins (Dataset EV1), TPR, a major structural constituent of the nuclear basket (Krull et al, 2004), ranks first in the number of unique peptides. Western blotting using an anti-TPR antibody validated the presence of TPR in the testicular lysates precipitated by the anti-S100PBP-antibody from the wild-type testes, but not S100pbp^−/−^ testes (Fig. 5B). To further confirm the interaction between S100PBP and TPR, we performed co-IP in lysates of HEK-293T cells co-expressing Flag-tagged S100PBP (Flag-S100PBP) and GFP-tagged TPR (GFP-TPR). The result showed that S100PBP interacts with TPR when expressed in cultured cells (Fig. EV3A,B). Interestingly, we noticed that Flag-S100PBP shows a nuclear localization when expressed in HEK-293T cells (Fig. EV3C), consistent with the previous report (Dowen et al, 2005). But when GFP-TPR is co-expressed, Flag-S100PBP anchors to the nuclear envelope and shows a complete co-localization with TPR at the nuclear pores (Fig. EV3D). These observations in cultured cells provided us a hint that S100PBP may localize to the nuclear pores in a TPR-dependent manner.
To further classify the localization of S100PBP, we performed immunofluorescence staining of S100PBP on spermatocyte smears. S100PBP signals were specifically found on the nuclear membrane of leptotene, zygotene, and early pachytene spermatocytes from wild-type mice, and were not observed in other testicular cells or the S100pbp^−/−^ spermatocytes (Fig. 5C). Furthermore, co-staining for S100PBP and TPR revealed a high degree of co-localization in both the spermatocytes and oocytes under the confocal laser scanning microscope (Figs. 5D and EV4).
To confirm whether the localization of S100PBP to the nuclear pores of meiocytes depends on TPR, we tried to knockout Tpr specifically in spermatocytes by crossing Tpr^flox/flox^ mice with Stra8-Cre transgenic mice in which Cre is activated in germ cells of testes at approximately 3 dpp (Lin et al, 2017) (Fig. EV5A,B). Tpr^flox/flox^; Stra8-cre mice are infertile and spermatogenesis is arrested at an early spermatogenic stage with few pre-leptotene/leptotene spermatocytes observed (Fig. EV5C–G). Immunofluorescence staining of testicular cell smears confirmed the absence of TPR signals in these spermatocytes, and interestingly, S100PBP loses the nuclear pore localization and becomes dispersed to the nucleus (Fig. 5E).
Taken together, these findings indicate that S100PBP is likely a nuclear pore-associated protein and localizes to the nuclear pores of meiocytes in a TPR-dependent manner.
The N-terminal amino acid residues 27-94 of S100PBP are crucial for interacting with TPR
To determine which regions of S100PBP interact with TPR, GFP-tagged various deletion mutants of S100PBP, including S100PBP-∆N (2–155 aa deletion), S100PBP-∆M (156–269 aa deletion), and S100PBP-∆C (270–396 aa deletion), were co-expressed with Flag-tagged TPR in HEK-293T cells, and the interaction of these S100PBP mutants with TPR was analyzed by co-IP using GFP-trap magnetic beads (Fig. 6A). S100PBP-∆M and S100PBP-∆C retain their interaction with TPR, while S100PBP-∆N is unable to interact with TPR (Fig. 6B). We next divided the N-terminal amino acid residues (2–155 aa) into three fragments and found that deletion of either 2-50 aa (∆N1) or 51-100 aa (∆N2), abolishes the interaction with TPR, whereas the deletion of 101-155 aa (∆N3) has no adverse effect on the interaction. This suggests that the N-terminal residues 2–100 are essential for the interaction between S100PBP and TPR (Fig. 6A,B).Figure 6. The N-terminal amino acid residues 27-94 of S100PBP are essential for the interaction with TPR.(A) Schematic representation of the mouse proteins: TPR tagged with Flag, and S100PBP and its mutants tagged with GFP. The Flag tag is indicated by the black box, and GFP tags are indicated by the green boxes. (B) Co-IP assays with HEK-293T cells transfected with TPR tagged with Flag and S100PBP or its mutant proteins tagged with GFP. IDR is an intrinsically disordered region. WT wild-type. Source data are available online for this figure.
The N-terminal residues 2–100 do not contain any characterized domain. In silico analysis using the Predictor of Natural Disordered Region (PONDR) (v.VLXT) predicts that S100PBP is enriched in intrinsically disordered regions (IDR), with the highest IDR predictive score value for the region spanning amino acid residues 27-94 (Appendix Fig. S4). To understand whether the interaction between S100PBP and TPR is mediated via this IDR region, we performed co-IP in HEK-293T cells co-expressing GFP-tagged S100PBP-∆IDR (27-94 aa) and Flag-tagged TPR. Intriguingly, the deletion of IDR abolishes the interaction of S100PBP and TPR (Fig. 6). These results showed that the N-terminal amino acid residues 27-94 of S100PBP are crucial for the interaction between S100PBP and TPR.
Discussion
Our study uncovers the crucial role of S100pbp in mouse meiotic recombination, i.e., facilitating XY crossover formation while limiting excess autosomal crossovers. We also show that S100PBP interacts with TPR and localizes to the nuclear pores of meiocytes likely in a TPR-dependent manner.
The localization of S100PBP suggests that it is more likely to play an indirect role in recombination, than directly processing the recombination intermediates. Indeed, among the candidate interacting proteins of S100PBP identified by MS, we did not find any DSB repair proteins, but the nucleoporin TPR, one of the main components of the nuclear basket (Krull et al, 2004), and some RNA-binding proteins. The nuclear basket, composed of TPR, NUP50, and NUP153, is well-known to regulate the nucleocytoplasmic transport of mRNAs and proteins in somatic cells and also participates in somatic DSB repair (Aksenova et al, 2020; Lee et al, 2020; Simon et al, 2018), but the role of nuclear basket components in mammalian meiosis has not been studied. This could be partially due to that, ablation of these proteins would cause developmental defects before the stage of meiosis, for example, primordial germ cells could not survive after deletion of Nup50 (Park et al, 2016), as well as what we found (few cells could reach the meiotic stage) after knocking out Tpr using the Stra8-Cre which is commonly used for conditional gene knockout in meiocytes. Previous reports have shown that even a modest change in the abundance of certain recombinant proteins can influence crossover formation in mice (Reynolds et al, 2013; Yang et al, 2015). It is thus hypothesized that S100PBP may regulate meiotic recombination by mediating the nucleocytoplasmic transport of recombination-related mRNAs and proteins through its interaction with TPR. Therefore, our S100PBP mutant mice could offer valuable insights into the role and underlying mechanisms of nuclear pore complexes in meiosis.
Several mutant mice have been reported to have decreased XY crossovers due to defects in DSB repair, such as mice mutant for Hfm1, Palb2, Tex11, and M1ap (Adelman and Petrini, 2008; Guiraldelli et al, 2013; Li et al, 2023; Simhadri et al, 2014; Xie et al, 2022). It should be noted that, in all these mutant mice, the autosomal crossovers are also decreased. In addition, we previously found that in Rad51ap2 mutant mice, XY crossover is decreased but autosomal crossover formation is not affected (Ma et al, 2022). Recently, it was reported that in mice lacking of ATF7IP2, a SETDB1-interacting protein localizing at the sex body, autosomes have more crossovers but XY crossover is reduced along with meiotic sex chromosome inactivation failure (Alavattam et al, 2024; Shao et al, 2023). They suggested that ATF7IP2 constraints autosomal axis length to influence autosomal crossover and modulates the histone modifications on sex chromosomes to ensure XY crossover, which imply an indirect function in recombination (Shao et al, 2023). In this study, we also found that XY and autosomal recombination present opposing changes after S100pbp is disrupted. Interestingly, the increased autosomal recombination in the S100pbp mutants is irrelevant to chromosome length, nor any obvious alteration in the formation of sex body, which are different from those in the Atf7ip2 knockouts. Though the molecular basis of the difference in recombination between sex chromosomes and autosomes in S100pbp mutants remains elusive, our study adds new evidence to the speculation that the DSB repair at the XY could be controlled by several different factors and are different from the autosomal recombination.
MSH4 and TEX11 are well-known for their roles in stabilizing intermediates for crossover formation. They show similar foci kinetics, with approximately 60% of TEX11 foci overlapping with MSH4 foci (Yang et al, 2008). Typically, their numbers change similarly when crossover formation is disturbed; for example, both are reduced in Redic1^−/−^ mice (Fan et al, 2023) and increased genome-wide in S100pbp^−/−^ spermatocytes. Intriguingly, though both MSH4 and TEX11 could localize to PAR (Ma et al, 2022), they are differentially affected in S100pbp knockouts. The reduction of the frequency of cells harboring TEX11/M1AP foci at the PAR at the early pachytene stage with touching XY PARs (69.58%/69.02% in S100pbp^−/−^ versus 95.24%/97.44% in controls) paralleles the increased frequency of untouching XY PARs in S100pbp^−/−^ mice from early to mid-pachytene stage (controls: 6.55% and 13.76% in the early and mid-pachytene stage, respectively; S100pbp^−/−^: 35.53% and 65.32% in the early and mid-pachytene stage, respectively), implying that the separation of XY chromosomes occurring from early to mid-pachytene stage could be associated with the absence of TEX11/M1AP at the PAR in early pachytene speramtocytes. In contrast, the localization of MSH4 at the PAR appears not adversely affected by the S100pbp mutation. Differences in TEX11 and MSH4 localization at the PAR were also reported in Atf7ip2^−/−^ pachytene spermatocytes but with MSH4 reduced and TEX11 unaffected (Shao et al, 2023). Based on these observations, we speculate that MSH4 and TEX11 are possibly differently regulated at the PAR, though further experiments are needed to confirm this.
In conclusion, our study revealed that S100pbp deficiency results in decreased XY recombination and increased autosomal recombination, offering a new model for exploring the differences in crossover formation between sex chromosomes and autosomes. In addition, our findings suggest that S100PBP’s regulatory role in meiotic recombination is likely linked to the nuclear pore complex and add a new dimension to the understanding of the regulatory processes underlying this core event of meiosis.
Methods
Reagents and tools tableReagent/resourceReference or sourceIdentifier or catalog number Experimental models HEK-293 cellsATCCCRL-3216C57BL/6JGemPharmatechStrain No. N000013ICRGemPharmatechStrain No. N000294S100pbp^−/−^ miceThis studyN/ATpr-flox miceGemPharmatechStrain No.T008224Stra8-Cre miceLin et al, 2017N/A Recombinant DNA Flag*-S100pbpThis studyN/AFlag-TprThis studyN/AGFP-S100pbpThis studyN/AGFP-TprThis studyN/AGFP-S100pbp-∆NThis studyN/AGFP-S100pbp-∆MThis studyN/AGFP-S100pbp-∆CThis studyN/AGFP-S100pbp-∆N1This studyN/AGFP-S100pbp-∆N2This studyN/AGFP-S100pbp-∆N3This studyN/AGFP-S100pbp-*∆IDRThis studyN/A Antibodies Details are available in Appendix Table S2 Oligonucleotides and sequence-based reagents
Primers for qPCR and RT-PCR Mouse S100pbp-FAAGGATTCTGGGGAGGCGAAMouse S100pbp-RAGGGCATCTAGCTCTCCCAAMouse Actb cDNA-FTAGGCACCAGGGTGTGATGGMouse Actb cDNA-RCGTACATGGCTGGGGTGTTG Primers for cloning vectors p-N1-FAGCGGCCGCGACTCTAGATCP-N1-GFP-linker-RTCCTGCAGCTCCACCGCTCGACTTGTACAGCTCGTCCATGCGFP-S100PBP-FTCGAGCGGTGGAGCTGCAGGAATGACGTGTTCACTCTTGCCGFP-S100PBP-RGATCTAGAGTCGCGGCCGCTTCACTGTTGATGGGATGAGAGFP-S100PBP-∆N-FTCGAGCGGTGGAGCTGCAGGAATGTCCTCCAAAGAAACGGAAAAGFP-S100PBP-∆N-RCATTCCTGCAGCTCCACCGCTCGAGFP-S100PBP-∆M-FTTGATAAAGACAAGATCGATTCTGGGGAGGCGAAAGGTGAGFP-S100PBP-∆M-RATCGATCTTGTCTTTATCAAGFP-S100PBP-∆C-FGACATGCCTTTGACAAGGATTGAAGCGGCCGCGACTCTAGGFP-S100PBP-∆C-RATCCTTGTCAAAGGCATGTCGFP-S100PBP-∆2-50 aa-FTCGAGCGGTGGAGCTGCAGGAATGTTCAGTTTCACAGAGGAAGAGFP-S100PBP-∆2-50 aa-RCATTCCTGCAGCTCCACCGCTCGAGFP-S100PBP-∆51-100 aa-FGAGAAGAAGATGATGGCCATTTCCTAAAACTACCTCAACTGFP-S100PBP-∆51-100 aa-RATGGCCATCATCTTCTTCTCGFP-S100PBP-∆101-155 aa-FCAGCAGCTGAAACCCCTGGCTCCTCCAAAGAAACGGAAAAGFP-S100PBP-∆101-155 aa-RGCCAGGGGTTTCAGCTGCTGGFP-S100PBP-∆27-94 aa-FGCAATGCCTCATTTCCTTGGGCAGCTGAAACCCCTGGCTTGFP-S100PBP-∆27-94 aa-RCCAAGGAAATGAGGCATTGCpEGFP-FGACTCTGGGGTTCGAAATGApEGFP-RTCATTTCGAACCCCAGAGTCP-MCS-Flag-RATCGTCATCGTCTTTGTAATCCATGGTGGCGACCGGTGGATCCCFlag-S100PBP-FGATTACAAAGACGATGACGATAAAATGACGTGTTCACTCTTGCCFlag-S100PBP-RGATCTAGAGTCGCGGCCGCTTCACTGTTGATGGGATGAGAGFP-TPR-FTCGAGCGGTGGAGCTGCAGGAATGACCTCTGGTGGCTCGGFP-TPR-RGATCTAGAGTCGCGGCCGCTTTAATTAATATTCCCTCTATTTATTCCTCCTCTTCCTTPR-HF1AGACCTGCGCTCACAAAACATPR-HR1TGTTTTGTGAGCGCAGGTCTTPR-HF2GAAGCAAGAAGTCTCCAGGATPR-HR2TCCTGGAGACTTCTTGCTTCFlag-TPR-FGATTACAAAGACGATGACGATAAAATGACCTCTGGTGGCTCGFlag-TPR-RGATCTAGAGTCGCGGCCGCTTTAATTAATATTCCCTCTATTTATTCCTCCTCTTCCT Primers for sgRNA transcription S100pbp-knockout (KO) -sgRNA-FGAAATTAATACGACTCACTATAGGGAGATCTTCATCCAAGGAACCCCAGTTTTAGAGCsgRNA-RAAAAAAGCACCGACTCGGTG Primers for mouse genotyping S100pbp-KO-FAGCCTCACCAACCCAAATCAS100pbp-KO-RCTGGTCCCAAGCTGTACAAAStra8-cre-FAACATTTGGGCCAGCTAAACStra8-cre-RCATCCTTAGCGCCGTAAATCTpr-cKO-5’-FACCTGTAGTTAGTCATAGACAGCTGGTpr-cKO-5’-RTGCTTTCACACTTAGAAACCGCTpr-cKO-3’-FTACTTCCGGGATAGGTAAGCATTGTpr-cKO-3’-RCTGTTTAAGACCCTTTTATACCACTG Chemicals, enzymes, and other reagents Anti-GFP nanobody magarose beadsAlpalifebioKTSM1334Bouin’s solutionSigmaHT10132Bromophenol blueSinoreagent71008060ClonExpress MultiS One Step Cloning KitVazymeC113-02DMEMVivaCellC3113-0500Dithiothreitol (DTT)SolarbioD0632Eosin YSolarbioE8080FastStart Universal SYBR Green Master (ROX)Roche04913850001Fetal Bovine Serum (FBS)GIBCO16000-044Giemsa stainSolarbioDM0002GlycerolBIO BASIC INCGB0232HCLSinoreagent10011018HematoxylinSangonA426825Lipofectamine 3000InvitrogenL3000015NaClSinoreagent10019318Opti-MEMGIBCO31985-070Paraformaldehyde (PFA)Sinoreagent80096618ParaplastLeica39601095Peniciuin-streptomycinGIBCO15140-122Phanta Max Master MixVazymeP525MEGA shortscript T7 kitThermo FisherAM1354Phenylmethanesulfonyl fluoride (PMSF)Thermo Fisher Scientific36978PrimeScript RT reagent kitTakaraRR047AProtein A/G agarose beadsSanta Cruzsc-2003Sodium dodecyl sulfate (SDS)Sinoreagent30166428SucroseSigmaS5390Tris-baseBiofroxx1115GR500Trisodium citrate dihydrateSinoreagent10019418Triton X-100SangonA600198-0500TRIzolTakara9109VECTASHIELDVectorH-1000 Software Prism 8GraphpadRRID:SCR_002798Image-Pro PlusMediaCyberneticsRRID:SCR_016879
Mice
S100pbp mutant mice were generated using CRISPR/Cas9 technology as we described previously (Jiang et al, 2017; Li et al, 2023). Briefly, the sgRNAs were designed targeting the exon 1 (for generating S100pbp^−/−^ mice) and transcribed in vitro using MEGA shortscript T7 kit (Thermo Fisher Scientific, AM1354). The obtained sgRNAs were microinjected into the zygotes of C57BL/6J mice together with Cas9 mRNA, and the zygotes were then transferred into the oviducts of pseudopregnant ICR female mice. Sanger sequencing with genomic DNA extracted from mouse toes was performed to detect the mutations in the mice of F_0_ generation. The founder mice carrying the mutation of interest in S100pbp were backcrossed with wild-type C57BL/6 mice for at least one generation and the obtained heterozygous mutant mice were intercrossed to produce homozygous mice. Tpr-flox mice (Strain NO.T008224) were purchased from GemPharmatech. Stra8-Cre mouse was gifted by Prof. Ming-Han Tong (Lin et al, 2017). All mouse experiments were conducted using adult mice aged between 8 and 12 weeks (unless specifically stated) and were carried out following the guidelines approved by the Institutional Animal Care Committee of USTC (approval number: USTCACUC25120122053). The primers for genotyping and sgRNAs used in this study are listed in Reagents and Tools Table.
RNA isolation, RT‒PCR, and qPCR
Total RNAs were extracted using TRIzol reagents (Takara, 9109) and cDNAs were synthesized from total RNAs using the PrimeScript RT reagent kit (Takara, RR047A) according to the manufacturer’s protocol. Phanta Max Master Mix (Vazyme, P525) was used for subsequent PCR under the following conditions: 3 min at 95 °C, followed by 35 cycles of 15 s at 95 °C, 15 s at 60 °C, and 30 s at 72 °C. The qPCR was performed with FastStart Universal SYBR Green Master (ROX) (Roche, 04913850001) using a StepOne Real-Time PCR System (Applied Biosystems). The qPCR reactions were performed under the following conditions: 10 s at 95 °C, 40 cycles of 10 s at 95 °C, and 30 s at 60 °C. Actb was used as the internal control. The sequences of the primers are listed in the Reagents and Tools Table.
Histology
Mouse testicular tissues were harvested and fixed in Bouin’s solution. Mouse ovaries were harvested and fixed in modified Davidson’s fluid overnight. After embedding in paraffin, the tissue was sectioned at 5 μm. Paraffin-embedded ovaries were serially sectioned and mounted on slides for counting the number of follicles. Hematoxylin and eosin (H&E) staining of the testes and hematoxylin staining of the ovaries were performed as we described previously (Ma et al, 2022).
Diakinesis/metaphase I chromosome spreading
Chromosome spreads of meiotic diakinesis/metaphase cells were prepared, as previously described, and stained with Giemsa (Jiang et al, 2017).
Spermatocyte chromosome spreads and immunofluorescence staining
Spermatocyte chromosome spreading and subsequent immunofluorescence staining was performed as previously reported (Jiang et al, 2014; Ma et al, 2022). Information about antibodies is available in Appendix Table S2.
Oocyte chromosome spreads
Fetal ovaries were collected from embryos at 17.5 dpc and oocyte spread as previously described (Hwang et al, 2018; Li et al, 2023). Briefly, ovaries were placed in the hypoextraction buffer, containing 30 mM tris (pH 8.2), 50 mM sucrose, 17 mM trisodium citrate dihydrate, 5 mM EDTA, 2.5 mM dithiothreitol (DTT), and 1 mM PMSF, for 15 min, then transferred into 100 mM sucrose, and teased apart with two needles. Twenty microliters of the cell suspension were added onto slides mounted with 200 μL of 1% paraformaldehyde containing 0.15% Triton X-100, evenly spread on the slides, and then incubated in a humid chamber for 2 h at room temperature. The slides were then air-dried and immunofluorescence staining was subsequently performed.
Co-IP and mass spectrometry
The mouse testes were lysed in IP buffer (50 mM Tris-HCl pH 7.5, 150 mM NaCl, 0.5% Triton X-100, and 2.5 mM EDTA) supplemented with 1 mM PMSF. The lysates were sonicated for 15 cycles (2 s on/off) with 12% pulses and centrifuged at 15,000 × g at 4 °C for 15 min. The supernatant was incubated with precleared Protein A/G agarose beads (Santa Cruz, sc-2003) and 2 μg anti-S100PBP antibody (epitope: residues 19–33 of mouse protein; customizedly produced by ABclonal) or rabbit IgG antibody (ABclonal, AC005). After incubation at 4 °C for 8 h, the agarose beads were washed in IP buffer three times and stored at −80 °C. Western blotting using the anti-S100PBP antibody, as well as silver staining, was performed to assess the quality of Co-IP. The IPed samples were subjected to mass spectrometry (MS) at the National Center for Protein Science Shanghai. The proteins identified by MS and the candidate proteins interacting with S100PBP are available in Dataset EV1.
Plasmids
To construct vectors expressing Flag-tagged or GFP-tagged mouse S100PBP protein and mouse TPR protein, the coding sequences of S100pbp and Tpr were amplified from mouse testis cDNA by PCR and ligated with the backbone from pEGFP-N1 vectors using the ClonExpress MultiS One Step Cloning Kit (Vazyme, C113) according to the manufacturer’s instructions. The mutated variants, including GFP-S100pbp-∆N, GFP-S100pbp-∆M, GFP-S100pbp-∆C, GFP-S100pbp-∆N1, GFP-S100pbp-∆N2, GFP-S100pbp-∆N3, and GFP-S100pbp-∆IDR, were constructed based on GFP-S100pbp. All vectors were verified by Sanger sequencing. The sequences of the primers for plasmid construction are listed in the Reagents and Tools Table.
Cell culture, transfection, and immunofluorescence
HEK-293T cells (ATCC, CRL-3216) were cultured in high-glucose Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10% FBS (Gibco, 16000-044), 100 U/mL penicillin, and 100 mg/mL streptomycin (Gibco, 15140-122) and maintained at 5% CO_2_. The cells were tested negative for mycoplasma contamination. Cells were passaged 2–3 times after thawing and transfected at 70%-90% confluency. Transfection of plasmids was performed using Lipofectamine 3000 (Invitrogen, L3000015) according to the manufacturer’s instructions.
Twenty-four hours after cell transfection, cells were fixed in 4% paraformaldehyde followed by immunofluorescence. The primary antibodies were incubated at 4 °C overnight and followed with secondary antibodies for 1 h at 37 °C. Finally, the slides were mounted with Vectashield medium (Vector Laboratories, H-1000). The antibodies used for cell immunofluorescence are listed in Appendix Table S2.
Co-IP in cultured cells
Forty-eight hours after cell transfection, cells were harvested for protein extraction and lysed in IP buffer supplemented with 1 mM PMSF. The obtained protein lysates were incubated with anti-GFP nanobody magarose beads (Alpalifebio, KTSM1334) under gentle rotation at 4 °C for 6 h. After washing with IP buffer three times, the beads were boiled in 1× SDS sample buffer (100 mM Tris-HCl pH 7.4, 2% SDS, 15% glycerol, 0.1% bromophenol blue and 5 mM DTT) for 10 min, followed by western blotting analyses.
Cell smear
Testes from adult mice and ovaries from embryos (16.5 dpc) were harvested. The tissues were placed in PBS supplemented with 10% FBS and teased apart to release cells. Cell suspension was added to one end of the glass slide and spread using a glass capillary tube gently. After air-drying, the slide was fixed in 4% PFA for 10 min and permeabilized in 0.2% PBST for 30 min, followed by immunofluorescence staining. Confocal images were captured using a Nikon C2 Plus Confocal Laser Scanning Microscope.
Statistical analysis
The results of each group were presented as mean ± SEM. All statistical analyses comparing S100pbp mutant mice with control mice (wild-type or heterozygous littermates) were performed using a two-tailed unpaired Student’s t test, as specified in the figure legends. The difference was considered significant when the P value was <0.05. No blinding, randomization, and sample size estimations were performed.
Supplementary information
Appendix Peer Review File Dataset EV1 Source data Fig. 1 Source data Fig. 2 Source data Fig. 3 Source data Fig. 4 Source data Fig. 5 Source data Fig. 6 Expanded View Figures