Longitudinal dynamics of plasma bile acids and their associations with physiological parameters and fecal microbiome during the transition period in dairy cows
Feixiang Fan, Liang Chen, Huizeng Sun, Jianxin Liu, Kailun Yang, Fengfei Gu

TL;DR
This study tracks changes in bile acids and gut microbes in dairy cows around calving, linking them to health indicators.
Contribution
The study identifies key bile acids and microbial biomarkers associated with periparturient health in dairy cows.
Findings
Plasma bile acids and fecal microbiome diversity changed significantly during the transition period in dairy cows.
Ursodeoxycholic acid and Alistipes were identified as key bile acid and microbial biomarkers.
Changes in bile acid concentrations correlated with physiological parameters like antioxidant capacity and triglyceride levels.
Abstract
The aim of this study is to investigate the dynamic changes of plasma bile acids (BA) and their correlations with physiological parameters and fecal microbiome in transitional dairy cows. Twenty multiparous dairy cows were selected, the blood and fecal samples were collected on d −21, −7, +7, and +21 of calving. The targeted metabolome and 16s rDNA ampicon sequencing were utilized to identify BA profiles and fecal microbial composition, respectively. A total of 32 BAs were found, comprising 9 primary BAs (PBA) and 23 secondary BAs (SBA). Majority of the PBAs (7 out to 9) and SBAs (15 out to 23) exhibited significant increases postpartum compared to prepartum levels. Notably, ursodeoxycholic acid, taurocholic acid and 7-ketodeoxycholic acid showed higher importance. Correlation analysis showed the BAs concentrations positively correlated with the concentrations of aspartate…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
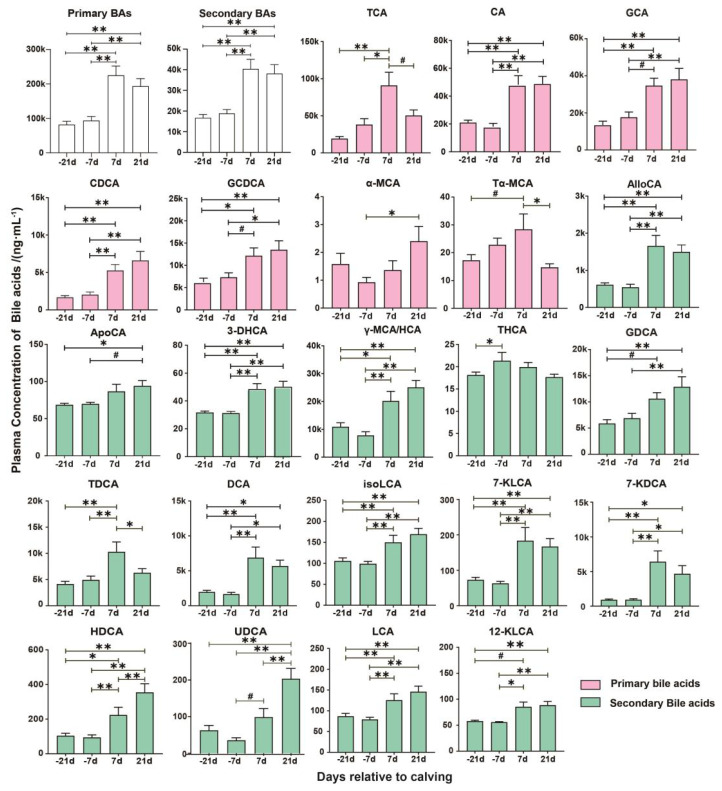 Figure 1
Figure 1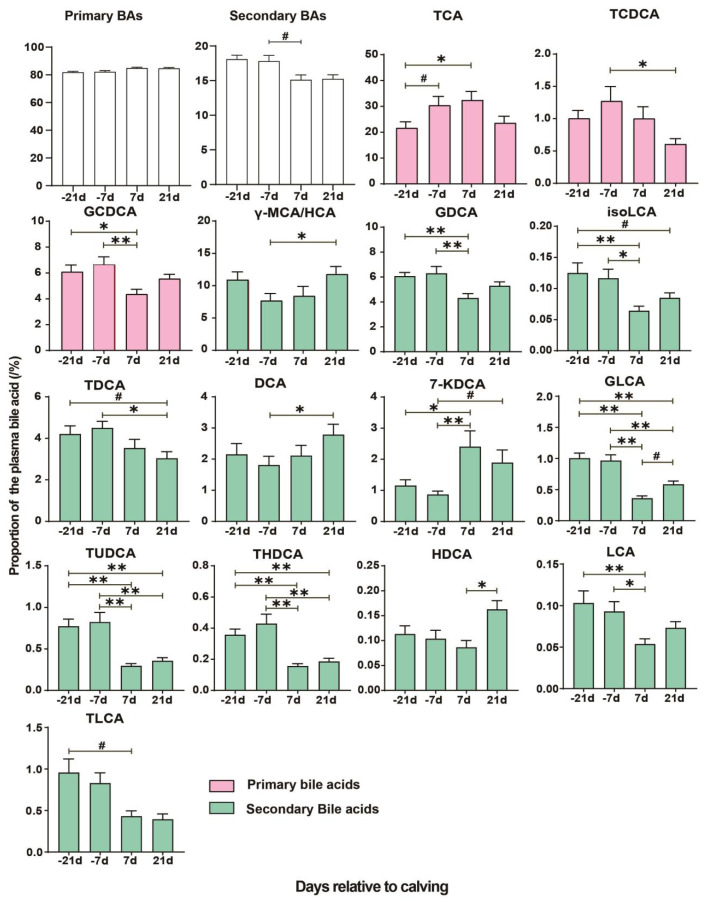 Figure 2
Figure 2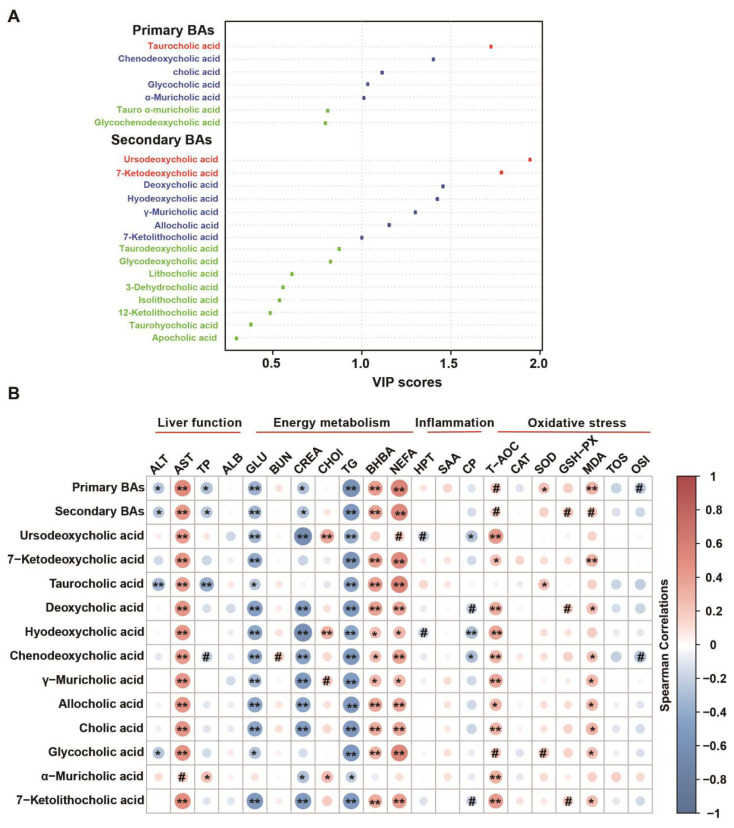 Figure 3
Figure 3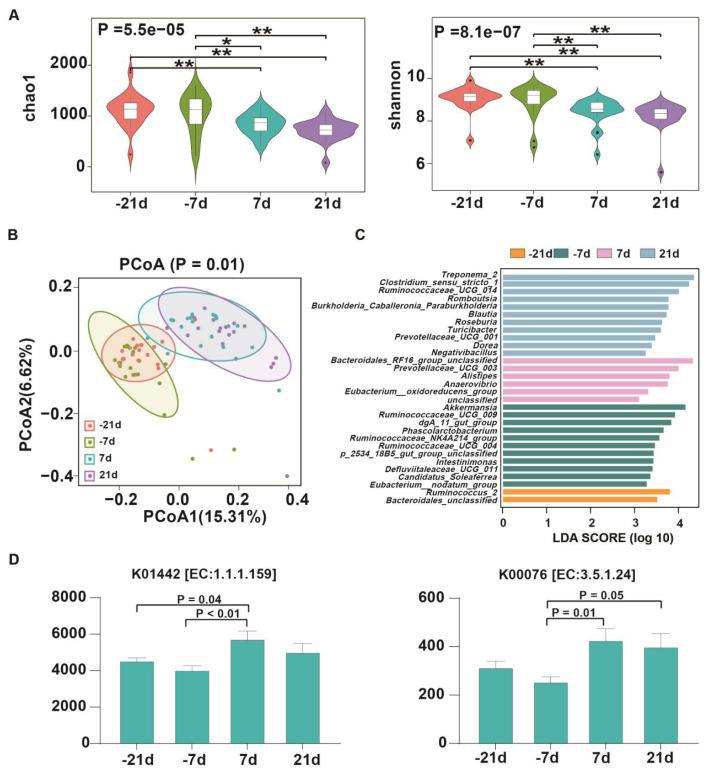 Figure 4
Figure 4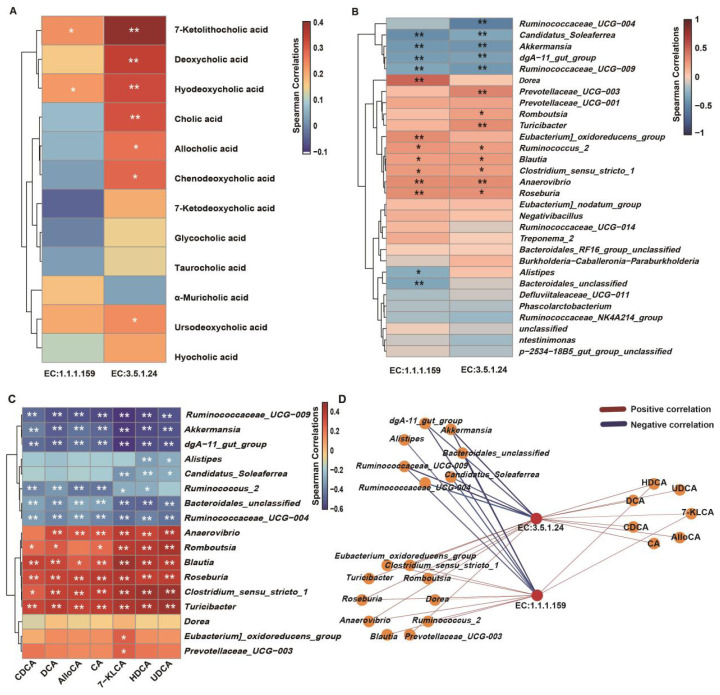 Figure 5
Figure 5Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsRuminant Nutrition and Digestive Physiology · Agricultural Productivity and Crop Improvement
INTRODUCTION
The transition period is a critical stage for dairy cows, during which the risk of diseases increases [1]. It has been reported that 30% to 50% of dairy cows experience health disorders, and approximately 75% of infectious diseases and metabolic disorders in dairy cows occur during this stage [2,3]. This is partly attributed to the complex metabolic adaptations during the transition period, when the cows are challenged by nutrient supply to meet the demands of calving and the onset of milk production [4]. Therefore, the metabolic changes during the transition period are as important and challenging as calving.
Bile acids (BAs), consisting of a polar carboxylate side chain and a nonpolar steroid carbon skeleton, are synthesized into primary BAs (PBAs) in the liver from cholesterol [5]. Meanwhile, gut microbiota modify PBAs to produce secondary BAs (SBAs) through dehydroxylation, deconjugation, or both [6]. Traditionally, BAs have been recognized for their roles in facilitating lipid and fat-soluble vitamin digestion and absorption [7,8]. Recently, numerous studies have unveiled their multifaceted functions in maintaining host metabolism [9], immune functions [10], intestinal health, and gut microbial homeostasis [11]. In terms of animal industry, BAs and related additives have been extensively employed in monogastric animals to improve growth, development, and health status [12–14]. However, the requirement and effects of BAs in dairy cows are not well understood.
For transitional dairy cows, the metabolic changes of BAs and their subsequent effects warrant attention. It is widely recognized that the profile and concentration of BAs are intricately associated with hepatic and intestinal microbiota [15]. However, the liver of postpartum dairy cows often suffers from metabolic stress and may develop varying degrees of hepatic steatosis, attributed to negative energy balance (NEB) and subsequent adaptive responses [16]. Our previous research revealed substantial alterations in the relative abundance, microbial composition, and functions of fecal microbiota over the transition period in dairy cows [17]. Furthermore, postpartum dairy cows experience adipose mobilization due to NEB [18]. Overall, it can be reasonably assumed that the profiles and contents of BAs change significantly due to variations in hepatic, gut microbiota, and lipid metabolism in transitional dairy cows. However, there is currently a dearth of systematic research on alterations in BAs during the transition period in dairy cows, with only sporadic discussions available [19]. On the other hand, a subsequent inquiry is the potential additional ramifications of the adapting alterations in BA on transitional dairy cows. Many studies have demonstrated the pivotal role of specific BAs such as ursodeoxycholic acid (UDCA) or bile salts in regulating inflammation [20] and oxidative stress [21]. Our previous study has also established a link between BAs and immunosuppression in dairy cows [22]. Hence, the relationships between dynamic changes in BAs and health status, such as inflammation and oxidative stress, are areas of significant interest in dairy cows during the perinatal period, yet remain incompletely understood.
Thus, the objectives of this study are to unravel dynamic alterations in peripheral BA profiles and concentrations in dairy cows over the transition period and to decipher the intricate associations between changes in BA and physiological metabolism and gut microbiome. Capturing the dynamic shifts in BA metabolism during this exceptional transition window and delineating its connections with health outcomes and gut microbiota hold great potential for improving transitional care in dairy cows.
MATERIALS AND METHODS
Animals and experimental design
The animal experimental design was described in our companion study [17] with minor modification. In brief, twenty multiparous healthy Chinese Holstein dairy cows at d 28 before parturition (body weight: 690 kg, standard deviation [SD] = 50; Parity: 1.93, SD = 1; BCS: 3.21, SD = 0.41) with similar body weight were selected. The definition of a healthy dairy cow includes two aspects: first, the cow was not experienced any clinical diseases within one month before the dry period according to veterinary records; second, during the trial, the cow is free from any clinical disease as confirmed by veterinary. The first 7 d (wk -4 relative to parturition) was the adaptation period. Ingredients of the basal diet before calving and after calving have been reported by Zhu et al [17] (Supplement 1). The experiment ended at d 21 after calving. Throughout the whole trial period, cows were housed in a barn with individual tie stalls and had free access to water. Prolonged labor lasting over 4 hours without urination and with the amniotic sac ruptured is considered dystocia, and assistance from manual delivery is required. High-risk cattle including those with retained placenta, twin calves, stillbirth, and birth canal tears, are closely monitored, and the characteristics of uterine discharge and the health status of the cattle are carefully recorded. If watery foul-smelling or purulent foul-smelling uterine discharge is present, uterine inflammation is diagnosed.
Blood and fecal sampling
Blood samples were collected into tubes containing an anticoagulant (heparin lithium) from the coccygeal vein of cows at 3 hours after the morning feeding at d −21, −7, 7, and 21 relative to the calving date. The samples were centrifuged at 3,000×g for 15 min at 4°C to collect plasma and then frozen at −20°C until subsequent analysis. An autoanalyzer 7020 instrument (Hitachi High-Technologies Corporation, Tokyo, Japan) was used to quantify concentrations of glucose (GLU; #ZH2079T), non-esterified fatty acids (NEFA; #ZH2045Z), β-hydroxybutyrate (BHBA; #ZH2029T), blood urea nitrogen (#ZH2017S), cholesterol (#ZH2040Z), triglycerides (TG; #ZH2039Z), albumin (#ZH2013G), alanine aminotransferase (ALT; #ZH2001G), aspartate aminotransferase (AST; #ZH2002G), total protein (TP; #ZH2012G), superoxide dismutase (SOD; #ZH2058F) and creatinine (#ZH2020S2) in plasma with commercial kits (Ningbo Medical System Biotechnology Co., Ltd., Ningbo, China). Likewise, commercial assay kits from Nanjing Jiancheng Bioengineering Institute (Nanjing, China) were used to quantify concentrations of plasma catalase (#A007-1-1), total antioxidant capacity (T-AOC; #A015-2-1), glutathione peroxidase (GSH-px; #A005-1-2), malondialdehyde (MDA; #A003-1-2), haptoglobin (HPT; #H136), amyloid (#H134) and ceruloplasmin (#A029-1-1) following the manufacturer’s instructions. Finally, plasma total oxidative status (TOS; #KC5100, Bensheim, Germany) was measured with commercial assay kits following manufacturer’s instructions [23]. The oxidative status indicator (OSI) was defined as the ratio of TOS to T-AOC [24,25]. Rectal fecal samples were collected before the morning feeding at 6:00 am using sterilized gloves and stored in 50 mL frozen storage tubes.
Profiling of blood bile acids
Profiling of BA was conducted according to a method described by Hu et al [26]. Briefly, plasma (100 μL) was directly incubated with precooled methanol and internal standard. After centrifuging at 14,000×g for 15 min, the supernatants were collected for vacuum drying and then resuspended in 100 μL methanol:water (1:1, v/v). Analyses were performed using an UHPLC (Waters Ltd., Milford, MA, USA) coupled online to 5500 QTRAP Mass Spectrometry (AB SCIEX, Framingham, MA, USA). The peak area and retention time were generated by Multiquant software. The internal standards of BAs were used to correct the retention time and to identify metabolites.
Microbial DNA extraction, sequencing, and pre-processing analysis
Microbial DNA from fecal samples was extracted using the E.Z.N.A. Stool DNA Kit (#D4015; Omega, Inc., Norcross, GA, USA). The universal 16S primers corresponding to the V3-V4 region (341F: 5’-CCTACGGGNGGCWGCAG-3’; 805R: 5’-GACTACHVGGGTATCTAATCC-3’) were used to amplify the bacterial 16S rRNA gene fragments. Purification and quality testing were performed using AMPure XT beads (Beckman Coulter Genomics, Danvers, MA, USA) and Qubit (Invitrogen, Waltham, MA, USA). Polymerase chain reaction (PCR) products were then pooled together and sequenced on an Illumina NovaSeq PE250, which was provided by LC-Bio Technology Co., Ltd, Hangzhou, China. The FLASH (V1.2.8) was applied to demultiplex the raw sequence data into paired-end fastq files. Alpha and beta diversities were calculated using QIIME2 [27]. The adonis test was used to determine significant differences caused by different categories of feed and host related metabolites. The significant differential microbiota was identified by LEfSe analysis. The functional components of the microbiome were predicted by PICRUSt2 program [28]. Among the twenty cows, eleven cows microbial data were shared with our companion study [17].
Correlation analysis
The Spearman’s rank correlation analysis (R packages Hmisc v 4.6.0) was used to determine the associations among the BAs, microbiota and plasma parameters in current study. The P-values were generated using the t or F distributions, and p<0.05 was regarded as significantly correlation.
Statistical analysis
The plasma BA and physiologic parameters data were analyzed using the PROC MIXED procedure with repeated measurement in SAS software (version 9.4), and covariance type AR (1) was selected. The model included the fixed effect of week and cow within the week as a random effect The experimental results were reported as least squares means, and the differences were separated using the PDIFF option in the LSMEANS treatment. Significance was declared at p≤0.05, and 0.05<p≤0.10 was considered a trend.
RESULTS
Dynamics of plasma bile acids concentration and relative proportion during transition period
In total, 32 BAs including 9 PBAs and 23 SBAs were identified from 80 plasma sample within 4 time points throughout the transition period, and the plasma concentration of BAs is presented in Supplement 2. Most of PBAs (7 out of 9) and SBAs (15 out of 23) showed significant differences (p<0.05) during the transition period, except for the taurochenodeoxycholic acid (TCDCA), tauroursodeoxycholic acid (TUDCA), β-muricholic acid (β-MCA),taurolithocholic acid (TLCA), taurohyodeoxycholic acid (THDCA), glycohyodeoxycholic acid (GHDCA), glycoursodeoxycholic acid (GUDCA) and ω-MCA.
Overall, total PBAs and SBAs significantly increased postpartum (p<0.01). Taurocholic acid (TCA) concentration peaked at d +7 compared to other time points (p<0.01), with no significant difference between d+21 and prepartum. At d +7, tauro α-MCA (Tα-MCA) was notably higher than at d +21 (p<0.01) and tended to be higher than at d −21 (p = 0.06). Conversely, α-MCA significantly increased at d +21 compared to −7 d (p<0.01), with no significant differences at other time points. Other PBAs, including cholic acid (CA), glycocholic acid (GCA), glycochenodeoxycholic acid (GCDCA), chenodeoxycholic acid (CDCA), all significantly increased postpartum. Regarding SBAs, apocholic acid (ApoCA) was significantly higher at d +21 than at d −21(p<0.01) and tended to be higher than at d −7 (p = 0.06). Taurohyocholic acid (THCA) notably increased at d −7 compared to d −21 (p<0.01), with no differences observed at other time points, glycodeoxycholic acid (GDCA) gradually increased, peaking at d +21 when significantly higher than at d −21 and d −7 (p<0.01). Taurodeoxycholic acid (TDCA) gradually increased, peaking at d +7, with significant differences compared to other time points. The UDCA concentration peaked at d +21, significantly higher than at d −21, −7, and +7 (p<0.01). Other SBAs content also significantly increased postpartum (Figure 1).
The changes in the relative proportion of BAs are depicted in Supplement 3 and the Figure 2. While the proportion of total PBA remained stable during the perinatal period, the total SBA tended to be lower at d +7 compared to −7 d (p = 0.07). However, temporal differences in 3 PBAs (TCA, TCDCA, GCDCA) and 12 SBAs were observed. Specifically, the proportion of TCA followed its concentration changes, with significantly higher proportions at d −7 and +7 compared to −21 d (p<0.01). Significant differences between −7 d and +21 d were observed in the proportion changes of γ-MCA/HCA and TCDCA, but the trend was converse. For GCDCA, the proportion at d +7 was significantly lower than at d −21 and −7 (p<0.05). In terms of dynamic changes in SBA proportion, the proportion of 8 SBAs (GDCA, isoLCA, TDCA, GLCA, TUDCA, THDCA, LCA, TLCA) decreased significantly postpartum compared to prepartum. Conversely, the proportion of 7-ketodeoxycholic acid (7-KDCA) increased significantly postpartum (p<0.05). At d +21, the proportion of deoxycholic acid (DCA) and hyodeoxycholic acid (HDCA) was significantly higher than at d −7 (p<0.01) and +7 (p<0.01), respectively.
Correlations of plasma bile acids with the blood parameters of the dairy cows during transition period
The variable importance in the projection (VIP) values were calculated to evaluate the importance of each BAs with time differences during the transition period, as shown in Figure 3A. Twelve BAs including 5 PBAs and 7 SBAs with VIP>1 and designated as key BAs. Among which, the TCA, UDCA, and 7-KDCA, exhibited higher VIP scores (VIP>1.5), with UDCA having the highest VIP score (VIP = 1.95).
The Spearman correlations between the total PBAs, SBAs and key BAs and plasma parameters refer to liver function, energy metabolism, inflammation and oxidative stress were presented in Figure 3B. The results showed that AST was positively correlated with total PBAs and SBAs, and most of the key BAs significantly (p<0.05) except for α-MCA. Conversely, ALT and TP showed negative correlations with total PBAs, SBAs, TCA significantly (p<0.05). The GCA is significantly negatively correlated with ALT (p = 0.03). Among the key BAs, the strongest correlations were observed with indicators of energy metabolism. Specifically, GLU and TG exhibited negative correlations with most key BAs, except for α-MCA. Conversly, BHBA and NEFA showed positive correlations, except for UDCA and α-MCA. Regarding inflammation parameters, only CP present negative correlations with three key BAs such as UDCA significantly, while HPT tended to correlate with UDCA and HDCA. Total PBAs and CDCA tended to correlate with OSI (p = 0.09; p = 0.06). SOD is significantly positively correlated with TCA (p = 0.04), and shows a positive correlation trend with total PBAs (p = 0.05) and GCA (p = 0.05). GSH-Px shows a significant positive correlation trend with total SBAs (p = 0.07), DCA (p = 0.09). MDA exhibited positive correlations with most key BAs, except for UDCA, TCA, HDCA and α-MCA.
Dynamics of fecal microbiome and associations with bile acids during transition period
A total of 5,500,172 raw reads were obtained from the sequencing data of 79 fecal samples collected from 20 dairy cows at four peripartum time points (−21 d, −7 d, and +21 d: N = 20; +7d : N = 19). Following quality control, 22,749 ASVs were assigned from these reads. The results revealed that Chao1 and Shannon indices at −21 d and −7 d were significantly higher than at +7 d and +21 d (p<0.001), while −21 d did not differ significantly from −7 d (Figure 4A). Moreover, microbial β-diversity among the four time points exhibited significant differences (Figure 4B, p = 0.001).
In addition, a total of 30 significantly different genera (p<0.01, linear discriminant analysis [LDA]>3) were identified throughout the transition period. Among these genera, Bacteroidales_unclassified and Ruminococcus_2 were regarded as biomarkers at −21 d; eleven genera, such as Ruminococcaceae_NK4A214_group and dgA_11_gut_group, were significantly higher at −7 d. At +7 d and +21 d, six and eleven biomarkers at the genus level were identified, respectively, with Eubacterium_oxidoreducens_group, Treponema_2, Clostridium_sensu_stricto_1, and Ruminococcaceae_UCG_014 showing higher relative abundances (Figure 4C). Functional prediction using PICRUSt2 revealed significant differences (p<0.05) in functions related to SBA biosynthesis between prepartum and postpartum (Figure 4D). Specifically, K01442 (choloylglycine hydrolase, EC:3.5.1.24) was significantly higher at +7 d compared to −21 d (p = 0.04) and −7 d (p<0.01), while K00076 (7-alpha-hydroxysteroid dehydrogenase [7α-HSDH], EC:1.1.1.159) showed significantly higher levels at +7 d (p = 0.01) and +21 d (p = 0.05) compared to −7 d. The correlations between microbial functions and key BAs are shown in Figure 5A. 7-KLCA and HDCA, exhibited positive correlations with EC:1.1.1.159 and EC:3.5.1.24 significantly (p< 0.05). DCA, CA, AlloCA, CDCA, and UDCA, exhibited positive correlations with EC:3.5.1.24 significantly (p<0.05). A total of 17 biomarker genera, including 7 and 10 genera such as Anaerovibrio and Ruminococcaceae_UCG_009 identified in Figure 4C, showed significant negative and positive correlations with EC:1.1.1.159 and EC:3.5.1.24, respectively (Figure 5B), and these genera also correlated with the seven BAs except for Dorea (Figure 5C). Consequently, a total of 17 important genera, 2 BA biosynthetic enzyme, and 7 key BAs were identified as playing crucial roles in BA metabolism during the transition period in dairy cows (Figure 5D).
DISCUSSION
The postpartum period in dairy cattle is characterized by significant metabolic changes, including alterations in endocrine hormones, glucose, and lipid metabolism. Among these, the pronounced lipid metabolism, resulting from energy imbalance, serves as the primary contributor to various metabolic disorders during this phase. BA play a crucial role in lipid digestion, absorption, and metabolism [7]. However, there is a paucity of research on dynamic changes in BA levels in transition dairy cows. This study aims to explore the dynamic changes in peripheral plasma BA levels in dairy cows during the transition period and analyze their associations with basic physiological metabolism. Additionally, the role of gut microbiota in this process was investigated. These findings provide fundamental insights into BA metabolism in transitional dairy cows and contribute to the development of strategies for healthy feeding practices in dairy cows.
The peripheral concentrations of PBAs and SBAs are all significantly elevated at postpartum, with an increase exceeding nearly twofold. This result could be considered a metabolic adaptation of dairy cows to cope with various physiological changes in transition period. The outcome is comprehensible easily, the first explanation is that the postpartum diets for dairy cows typically contain higher fat and energy content. Additionally, the NEB of postpartum cows is more serious and the body fat is mobilized to meet the energy requirements of milking [29]. Consequently, the lipid metabolism is heightened in postpartum dairy cows. Consistent with this, the concentrations of NEFA and BHBA in this study showed a positive correlation with BA concentration, while glucose and TG were correlated negatively, indicating that body fat mobilization and lipid metabolism caused by NEB during perinatal period would promote the production of BA. Our previous study found that the BA concentration was significantly higher in the postpartum cows that with higher plasma NEFA concentration compared to thatcows with lower NEFA [22]. Currently, Dicks et al [30] reported the higher BAs content in postpartum cows and showed strong correlations with body scores and lipid mobilization. These results corroborate our experimental findings and support our hypothesis that postpartum cows require more BAs to adapt to the intense lipid metabolism.
The homeostasis of peripheral BAs depends on the regulations of liver and gut microbiome [6]. Previous research have shown a significant increase in the “Bile secretion” pathway in postpartum cows compared to the prepartum cows through liver the transcriptome analysis, although they did not focus on this topic [31,32]. Although this study lacked liver tissue data, the significant correlation between liver function biomarkers, especially AST, and total BAs and key BAs partially explains this phenomenon.
The gastrointestinal microbiome play important roles in the growth and development of calves [33] as well as the shape of traits in dairy cows [34]. Here, we investigated the response of the intestinal microbiome to the increased demand for BAs in transitional dairy cows. Consistent with the increase in BA concentration postpartum, the relative abundance of certain bacteria genus enriched with BA metabolism genes was significantly increased, for instance, the genus Alistipes. Lin et al [35] found the bile salt hydrolase (BSH)-carrying metagenome-assembled genomes predominately assigned to genus Alistipes by using the genome-centric approach in dairy cows. Zhuge et al [36] reported the positive correlation between BAs and Alistipes. In addition, the relative abundance of Ruminococcaceae_UCG-014 and Negativibacillus were also higher in postpartum, and they belong to the BSH and 7α-HSDH-active family Ruminococcaceae, indicating they may have the ability to convert some PBAs into SBAs [37,38]. Consistent with the changes of bacterial genus, the abundance of two microbial enzymes, BSH and 7α-HSDH which involved in SBA biosynthesis were identified significantly increased in the postpartum dairy cows based on PICRUSt2 functional prediction. Taken together, these results suggest that both the liver and gut microbiota of dairy cows undergo functional changes to adapt to the increased demand for BAs during the transition period.
Inflammation and oxidative stress are major challenges suffered by perinatal dairy cows [39]. The UDCA, an SBA, is widely used in the treatment of cholestatic liver diseases [40], and has been shown to alleviate intestinal inflammation in neonatal dairy calves [41], early-weaned lambs [42], and piglets [20]. Additionally, certain BAs, such as TCA, have been reported to alleviate oxidative stress [43]. Consistent with these findings, our study observed a negative correlation between BA concentrations and inflammatory markers, as well as oxidative stress indicators. Our previous study has been revealed the links between BAs and immune-suppression of monocyte in transitional dairy cows [22]. Theses results suggest that the increased BAs may have benefits in alleviating inflammation and oxidative stress and improving the health of perinatal dairy cows. Future research should focus on systematically investigating the impact of BAs on the health of dairy cows, which could yield novel nutritional regulatory strategies for optimal management of dairy cows in transition period.
CONCLUSION
In conclusion, this study unveiled the dynamic changes in peripheral BAs in dairy cows during the transition period. Our findings indicate a significant increase in BA concentrations postpartum, which positively correlate with indicators of energy metabolism such as NEFA and BHBA and oxidative stress markers, while negatively correlating with inflammation. Key BAs, including UDCA, were identified as significant contributors to these dynamics. We further explored the adaptation of the intestinal microbiome to BAs, revealing significant increases in the relative abundance of BSH and 7A- HSDH in postpartum stage. Importantly, we identified key genera, such as Alistipes and members of the Ruminococcaceae family, as pivotal players in this process. These findings underscore a heightened demand for BAs to accommodate robust lipid metabolism after calving in dairy cows, suggesting potential regulatory effects of BAs on dairy cow health. Future studies should systematically investigate the effects and regulatory mechanisms of supplementing key BAs or bile salt additives on the performance and health of transitional dairy cows. Such endeavors hold promise for offering novel perspectives and strategies to promote the healthy feeding of perinatal dairy cows.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ingvartsen KL Feeding- and management-related diseases in the transition cow: physiological adaptations around calving and strategies to reduce feeding-related deseases Anim Feed Sci Technol 200612617521310.1016/j.anifeedsci.2005.08.003 · doi ↗
- 2Ingvartsen KL Dewhurst RJ Friggens NC On the relationship between lactational performance and health: is it yield or metabolic imbalance that cause production diseases in dairy cattle? a position paper Livest Prod Sci 20038327730810.1016/s 0301-6226(03)00110-6 · doi ↗
- 3Le Blanc SJ Lissemore KD Kelton DF Duffield TF Leslie KE Major advances in disease prevention in dairy cattle J Dairy Sci 20068912677910.3168/jds.s 0022-0302(06)72195-616537959 · doi ↗ · pubmed ↗
- 4Pascottini OB Leroy JLMR Opsomer G Maladaptation to the transition period and consequences on fertility of dairy cows Reprod Domest Anim 202257213210.1111/rda.1417635686392 · doi ↗ · pubmed ↗
- 5Russell DW Setchell KD Bile acid biosynthesis Biochemistry 19923147374910.1021/bi 00135 a 0011591235 · doi ↗ · pubmed ↗
- 6Collins SL Stine JG Bisanz JE Okafor CD Patterson AD Bile acids and the gut microbiota: metabolic interactions and impacts on disease Nat Rev Microbiol 2023212364710.1038/s 41579-022-00805-x 36253479 PMC 12536349 · doi ↗ · pubmed ↗
- 7Peet DJ Turley SD Ma WZ Janowski BA Mangelsdorf DJ Cholesterol and bile acid metabolism are impaired in mice lacking the nuclear oxysterol receptor LXRαCell 19989369370410.1016/s 0092-8674(00)81432-49630215 · doi ↗ · pubmed ↗
- 8van Olst N Meiring S Brauw MD Small intestinal physiology relevant to bariatric and metabolic endoscopic therapies: incretins, bile acid signaling, and gut microbiome Tech Innov Gastrointest Endosc 2020221091910.1016/j.tige.2020.03.003 · doi ↗