Systematic use of protein free energy changes for classifying variants of uncertain significance: the case of IFT140 in Mainzer-Saldino Syndrome
Macarena Gajardo, José Luis Guerrero, Bárbara Poblete, Esperanza Bayyad, Ignacio Castro, Jorge Maturana, Jaime Tobar, Víctor Faúndes, Paola Krall

TL;DR
This paper shows how changes in protein stability can help classify uncertain genetic variants in a rare disease called Mainzer-Saldino Syndrome.
Contribution
The study establishes a specific ΔΔG cut-off for IFT140 variants, enabling accurate reclassification of variants of uncertain significance.
Findings
IFT140 variants in Mainzer-Saldino Syndrome have significantly lower ΔΔG values compared to general population variants.
A ΔΔG cut-off of −1.3 kcal/mol effectively distinguishes pathogenic from benign IFT140 variants.
Applying this cut-off reclassified a previously uncertain variant as likely pathogenic, confirming a molecular diagnosis.
Abstract
Advanced genetic strategies have transformed our understanding of the genetic basis and diagnosis of many phenotypes, including rare diseases. However, missense variants (MVs) are frequently identified and often classified as variants of uncertain significance (VUS). Although changes in protein free energy (ΔΔG) were recently proposed as a tool for VUS classification, no objective cut-offs exist to distinguish between benign and pathogenic variants. We utilized the computational tool mCSM to calculate ΔΔG and predict the impact of MVs on protein stability. Specifically, we systematically analyzed the ΔΔG of MVs in IFT140 to identify those potentially pathogenic and associated with Mainzer-Saldino syndrome (MSS). To this end, we evaluated ΔΔG in IFT140 MVs sourced from ClinVar, gnomAD, and MSS patients, aiming to resolve the diagnosis of MSS in a child with a novel homozygous IFT140…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
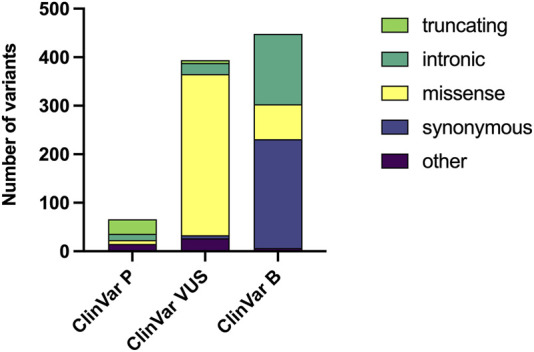 FIGURE 1
FIGURE 1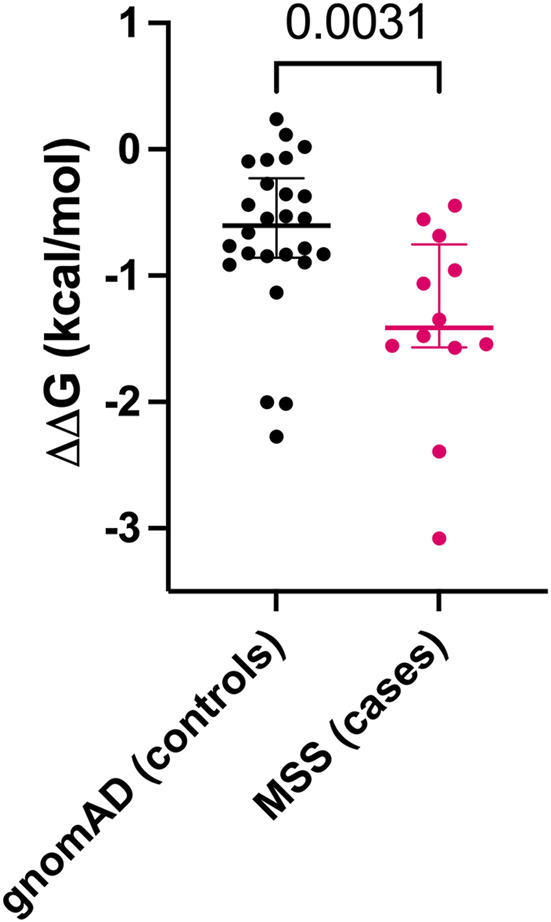 FIGURE 2
FIGURE 2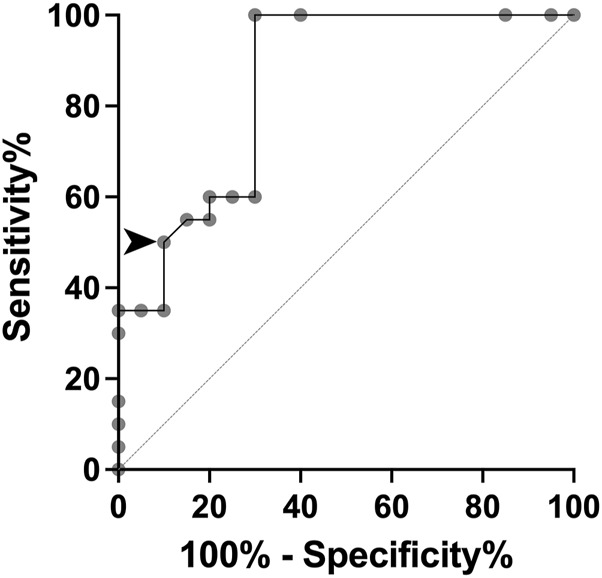 FIGURE 3
FIGURE 3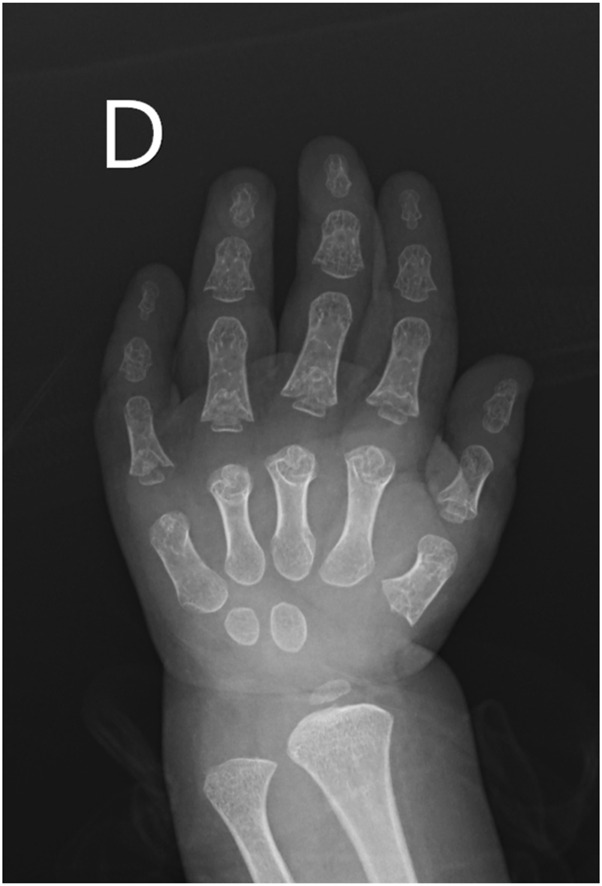 FIGURE 4
FIGURE 4| Index | Wild type residue | Position | Mutant residue | Predicted ΔΔG | Outcome |
|---|---|---|---|---|---|
| 1 | W | 80 | A | −3.989 | Highly Destabilizing |
| 2 | W | 80 | V | −3.545 | Highly Destabilizing |
| 3 | W | 80 | L | −3.277 | Highly Destabilizing |
| 4 | W | 80 | G | −4.16 | Highly Destabilizing |
| 5 | W | 80 | S | −3.146 | Highly Destabilizing |
| 6 | W | 80 | T | −2.99 | Highly Destabilizing |
| 7 | W | 80 | Q | −2.936 | Highly Destabilizing |
| 8 | W | 80 | E | −3.537 | Highly Destabilizing |
| 9 | W | 80 | C | −1.745 | Destabilizing |
| 10 | W | 80 | R | −1.994 | Destabilizing |
| 11 | W | 80 | P | −3.545 | Highly Destabilizing |
| 12 | W | 80 | D | −3.632 | Highly Destabilizing |
| 13 | W | 80 | F | −2.661 | Highly Destabilizing |
| 14 | W | 80 | I | −3.277 | Highly Destabilizing |
| 15 | W | 80 | H | −2.97 | Highly Destabilizing |
| 16 | W | 80 | N | −2.932 | Highly Destabilizing |
| 17 | W | 80 | M | −2.535 | Highly Destabilizing |
| 18 | W | 80 | Y | −2.418 | Highly Destabilizing |
| 19 | W | 80 | K | −2.516 | Highly Destabilizing |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsRNA regulation and disease · Fetal and Pediatric Neurological Disorders · Genetic and Kidney Cyst Diseases
Introduction
Advanced genetic strategies have transformed our understanding of the basis and diagnosis of many diseases. Protein-truncating variants (PTV) are usually considered pathogenic due to their strong functional effects, with few exceptions. However, missense variants (MV) are more frequent than PTV and often classified as variants of uncertain significance (VUS) due to limited population frequency data and functional evidence, underscoring the need for improved methods of resolution (Richards et al., 2015), (Appelbaum et al., 2022).
Short-rib thoracic dysplasia 9 (SRTD9, OMIM#266920), or Mainzer-Saldino Syndrome (MSS), is an extremely rare ciliopathy affecting fewer than 1 in 1,000,000 individuals. MSS is characterized by chronic kidney disease (CKD), progressive vision impairment, with distinctive skeletal features such as cone-shaped epiphyses (Geoffroy et al., 2018). Around 20 MSS cases have been reported, involving homozygous or compound heterozygous variants in the Intraflagellar Transport 140 (IFT140) gene. IFT140 contains multiple WD40 repeat (WD) and tetratricopeptide-repeat (TPR) domains that enable the protein to interact with others to form complex structures, facilitating the transport of proteins along the cilia. Accurate interpretation of IFT140 variants is essential for MSS diagnosis, guiding personalized treatment, avoiding unnecessary interventions, informing prognosis, and identifying at-risk family members (Walsh et al., 2024). The identification of novel MVs in IFT140 has a high likelihood of being classified as VUS, delaying diagnosis and limiting their clinical utility (Chen et al., 2023).
Several methods have been developed to enable VUS reclassification. These include population allele frequencies, functional assays, and machine learning models, with structural models specifically providing insights into the underlying molecular mechanisms. Gibbs free energy (ΔG) is a thermodynamic measure used to quantify protein stability, based on the principle that structures tend to adopt more negative energy states. If the difference between the folded and native forms of a protein results in a more negative value, the process occurs spontaneously. In this context, the change in Gibbs free energy (ΔΔG) evaluates the difference between the Gibbs free energies of a wild-type and a mutant protein. A negative ΔΔG value indicates that the mutant protein is destabilizing (Sapozhnikov et al., 2023). Recently, the Association for Clinical Genomic Science (ACGS 2024) recommended the use of ΔΔG caused by MVs for their classification, based on a successful clinical experience (Caswell et al., 2022), (ACGS, 2024). However, the proposed ΔΔG cut-offs were subjectively determined as they did not consider the full spectrum of ΔΔG seen in both benign and pathogenic MVs (Caswell et al., 2022).
The mutation Cutoff Scanning Matrix (mCSM) was introduced in 2014 to predict MVs effects by assessing protein structure changes and stability using ΔΔG (Pires et al., 2014). It offers a computationally efficient, easy-to-use and well-validated approach with optimal correlation to experimental ΔΔG data. When analyzing amino acid substitutions in different human diseases, mCSM predicted a higher proportion of variants as destabilizing and demonstrated better predictive power compared to other structure-based tools (Choudhury et al., 2022). Herein, we aimed to systematically establish a ΔΔG cut-off for IFT140 MVs using mCSM, which helped to confirm the molecular diagnosis of MSS in a child with a novel homozygous IFT140 variant, initially classified as VUS.
Materials and methods
First, we aimed to compare the ΔΔG seen in benign versus pathogenic IFT140 MVs to establish a cut-off that allowed us to distinguish between these two groups and reclassify VUS deposited in ClinVar. For this purpose, we downloaded IFT140 variants from ClinVar Miner (version 06-30–2024) with their ACMG classifications: P and LP were combined into the pathogenic (ClinVar-P) group, B and LB were combined into the benign (ClinVar-B) group, and VUS (ClinVar-VUS) were kept as a separate group (Henrie et al., 2018). Minor allele frequency (MAF) was assessed in each group using the gnomAD v4.1 database. Homozygous IFT140 MVs were obtained from gnomAD, focusing on LB/B classifications according to ClinVar and assumed as controls (Chen et al., 2024). Homozygous or compound heterozygous IFT140 MVs associated with MSS phenotype were collected and considered as cases (Geoffroy et al., 2018), (Patel et al., 2023), (Perrault et al., 2012), (Yeh et al., 2022), (Soyaltın et al., 2018), (Schmidts et al., 2013), (Oud et al., 2018). PTVs or synonymous IFT140 variants were excluded. The PDB file for IFT140 (8BBG, 3.50 Å, chain B, positions 1–1462) was downloaded from UniProt and used to calculate ΔΔG for each MV using mCSM (https://biosig.lab.uq.edu.au/mcsm/) (Pires et al., 2014). Mean ΔΔG values from gnomAD and MSS variants were compared using Mann-Whitney U test, and Receiver Operating Characteristic (ROC) curve was constructed to define a ΔΔG cut-off. In addition, the distribution of variants in IFT140 was evaluated, focusing on the 7 WD and 9 TPR domains according to the information available on UniProt. We made a statistical analysis of the position within and outside these domains comparing gnomAD and MSS patients. Chi square test was considered to determine statistical significances of these distributions.
Second, we aimed to reclassify a novel homozygous IFT140 MV seen in a patient with a phenotype compatible with MSS and considering the obtained cut-off, and to study the relevance of the affected residue for the syndrome. For these purposes, detailed clinical history and exome data were collected and informed consent was obtained from parents. The identified IFT140 variant and all possible residue changes underwent systematic ΔΔG analysis using mCSM, to compare those values with the cut-off. The study was revised and approved by the Ethics Committee from the Servicio de Salud Los Ríos.
Results
We obtained 908 genetic changes registered in ClinVar as variants compromising IFT140. These changes included 412 (45.4%) MVs, 230 (25.3%) synonymous variants, 181 (19.9%) intronic variants with different possible impacts, 49 (5.4%) rearrangements that involved IFT140, and only 36 (4.0%) PTVs.
When considering their classification, 66 variants were ClinVar-P, 394 variants were considered ClinVar-VUS, and 448 variants were ClinVar-B (Figure 1). In the ClinVar-P group, 43 (65.2%) variants were PTVs. MVs represented 84.3% of all variants in ClinVar-VUS, while they were found in 16.1% and 12.1% of the ClinVar-B and ClinVar-P groups, respectively.
Distribution of genetic variants by pathogenicity classification and variant type. The number of genetic variants downloaded from ClinVar Miner are displayed, categorized by clinical significance as Pathogenic (P), Variant of Uncertain Significance (VUS), and Benign (B). Within each bar, the abundance of different variant types (e.g., truncating, missense) is represented.
The gnomAD database showed the MAF for 196 of all the MVs in the different ClinVar groups. The average MAFs in the ClinVar-P, ClinVar-VUS and ClinVar-B groups were 0.475, 0.250 and 38.76 per 10,000 individuals, respectively. Of note, 29 (49.1%) out of 59 MVs in the ClinVar-B group had a MAF of less than 1 in 10,000, while in the ClinVar-VUS group, this characteristic was observed in 130 out of 133 MVs.
We calculated ΔΔG for gnomAD individuals with homozygous IFT140 MVs (n = 26) and for MSS patients (3 homozygous and 9 heterozygous MVs). The mean ΔΔG for gnomAD MVs was −0.681 kcal/mol (95%CI: −0.937 to −0.426) and for MSS MVs it was −1.389 kcal/mol (95%CI: −1.872 to −0.907). The difference in ΔΔG values between gnomAD and MSS IFT140 MVs was significant (U = 64, p = 0.0031) (Figure 2).
Comparison of ΔΔG values for missense variants (MVs) between gnomAD individuals (controls) and Mainzer-Saldino Syndrome (MSS) patients (cases). Each dot represents a single MV, with median values and interquartile ranges shown for both groups.
By using our ΔΔG data in gnomAD individuals and MSS patients, we constructed the ROC curve and obtained an AUC of 0.8488 (p = 0.0002), indicating an optimal discriminative ability to identify likely pathogenic MVs. The cut-off point was established at ΔΔG -1.3 kcal/mol, yielding a sensitivity of 50% and a specificity of 90% (Figure 3). For ClinVar groups, the average ΔΔG values were −1.432 kcal/mol (95%CI: −2.074 to −0.789), −0.798 kcal/mol (95%CI: −0.878 to −0.717), and −0.796 kcal/mol (95%CI: −0.953 to −0.640), for IFT140 MVs classified as pathogenic, VUS, or benign, respectively. Of note, in the ClinVar-VUS group, 75/323 MVs had ΔΔG values below −1.3 kcal/mol (Supplementary Figure S1).
ROC curve based on ΔΔG values for missense variants (MVs) from gnomAD controls and MSS cases. The arrowhead indicates the established cut-off value to achieve 50% sensitivity and 90% specificity.
The analysis from gnomAD individuals and MSS patients regarding their MV positions within and outside WD and TPR domains showed that 7 out of 26 variants (27%) reported in gnomAD homozygous individuals and 5 out of 9 variants (56%) identified in MSS patients were in these domains. However, this difference did not reach statistical significance (p = 0.1255).
A novel homozygous IFT140 variant was discovered in a 1.5-year-old girl who was evaluated for CKD stage 4 with small kidneys without cystic pattern, hepatic fibrosis, neurological developmental delay, and unilateral moderate sensorineural hearing loss. The array comparative genomic hybridization was normal, PKD1 analysis by long-range PCR sequencing was negative, but exome sequencing identified the IFT140 c.240G>T (p.W80C) homozygous MV categorized as VUS. This variant was inherited from both heterozygous parents aged 34 and 43 years, who underwent abdominal ultrasounds, with no relevant findings observed, particularly in the kidney structures (Dordoni et al., 2024). Subsequent evaluations in the patient revealed retinal dystrophy, and phalangeal cone-shaped epiphyses in both hands (Figure 4), confirming the clinical diagnosis of MSS. The ΔΔG value for IFT140:p.W80C was −1.745 kcal/mol, and further systematic evaluation of the W80 residue indicated that all possible amino acid substitutions were destabilizing (Table 1).
X-ray image of the right hand from the pediatric patient suspected of having Mainzer-Saldino Syndrome (MSS). The image reveals characteristic skeletal abnormalities associated with MSS (cone-shaped epiphyses).
Discussion
Rare diseases are a heterogeneous group of conditions with an emerging global impact, a population prevalence of 3.5%–5.9% and with a genetic origin in almost 80% (Nguengang et al., 2020). The rate of VUS in genes associated with rare diseases is 41.5%, and MVs represent the largest proportion of them (86.6%) (Chen et al., 2023). Half of the IFT140 benign MVs had a MAF<1/10,000 suggesting that rarity of a variant is not a reliable indicator of pathogenicity and reinforcing the need of additional methods for reanalysis.
In recent years, increasing interest in predicting the pathogenicity of MVs VUS using structure-based algorithms has emerged (Pan and Theesfeld, 2024). One such approach involves calculating the ΔΔG between the wild-type and variant residues; however, establishing a valid cut-off value is essential (Pires et al., 2014), (David and Sternberg, 2023). Recent studies have demonstrated that ΔΔG is a useful tool for evaluating other MVs in genes such as transcription factor FOXD2 and TBC1D31 which are now implicated in syndromic congenital anomalies of the kidney and urinary tract (Riedhammer et al., 2024; Saygılı et al., 2023), as well as NUP85, associated with steroid-resistant nephrotic syndrome (Şükür et al., 2025). In the field of protein stability, various computational tools aid in the assessment of MVs, including mCSM, Dynamut2 (Rodrigues et al., 2021), FoldX (Schymkowitz et al., 2005) and PremPS (Chen et al., 2020), among others. These tools evaluated stability, folding and dynamics of proteins. Additionally, molecular dynamics simulations offer valuable insights into atomic movements within a protein by modeling interatomic interactions (Hollingsworth and Dror, 2018).
The ΔΔG in IFT140 MVs revealed significant differences between benign (gnomAD individuals) and pathogenic (MSS patients) variants. We established a cut-off value of −1.3 kcal/mol with an optimal sensitivity and specificity for IFT140 MVs. When analyzing the totality of MVs in the ClinVar-VUS group, we identified that 23.2% had ΔΔG values below the cut-off value, suggesting that these might be re-classified as likely pathogenic.
The novel variant IFT140:p.W80C found in our female patient, initially classified as VUS, is a good example of the impact of our work. The child met all clinical features to be diagnosed with MSS (Soyaltın et al., 2018), and the ΔΔG value of −1.745 kcal/mol for p.W80C, below our calculated cut-off, confirms the deleteriousness of this variant. Additionally, the destabilizing values of all possible substitutions at this position indicate its intolerance to amino acid changes, which allowed us to reclassify the p.W80C as likely pathogenic, following ACGS 2024 recommendations (ACGS, 2024).
Our study has limitations in different aspects. Firstly, few patients with MSS and gnomAD IFT140 homozygous individuals have been reported, which decreases the sensibility of our findings. However, this is the first study to our knowledge that systematically evaluates a ΔΔG cut-off value in patients with IFT140 VUS, which has the potential to be used for the evaluation of other similar patients. Further calculations of ΔΔG cut-offs for other monogenic diseases are required to implement the recent ACGS 2024 recommendations of structural damage for MVs classification (ACGS, 2024).
Bioinformatic tools that evaluate ΔΔG have several limitations, many of which depend on structural data availability. Most require a high-resolution 3D protein structure, typically from Protein Data Bank (PDB). If the variant of interest is in a flexible or disordered region that is not well-captured in the structure, predictions can be unreliable. When no structure is available, these tools must rely on homology modeling, which introduces additional errors compared to crystal structures. Additionally, environmental effects are often poorly accounted for in these tools. Factors such as solvent interactions, post-translational modifications, in vivo physiological conditions, and protein-protein interactions are either oversimplified or completely ignored. This is particularly problematic when multiple pathogenic mechanisms coexist. Finally, ΔΔG prediction tools can yield conflicting results, as they rely on different algorithms and training datasets. To improve reliability, it might be reasonable to use multiple tools and compare their outputs.
By 2030, VUS in coding regions are expected to be resolved through several advancements, including refinements in variant classification standards, improved performance of computational variant effect predictors, the establishment of large-scale ΔΔG datasets, the development of hybrid in silico and experimental approaches, and enhanced data-sharing efforts that maximize the information gained from each newly sequenced individual and interpreted variant (Fowler and Rehm, 2024). Additionally, machine learning approaches will play a key role by integrating large-scale genomic, functional, and clinical data, improving predictive accuracy, and identifying complex patterns that traditional methods might overlook (Quazi, 2022; Khalifa and Albadawy, 2024).
In conclusion, the analysis of ΔΔG can aid in the evaluation of IFT140 MVs for diagnostic purposes. The appropriate approach should include the assessment of clinical findings, the evaluation of genetic variants using meta-predictors or other assays, with ΔΔG serving as one of the components of the comprehensive strategy in patients suspected to have MSS or other rare diseases. Future advancements in variant classification standards, computational predictors, multiplexed assays, and machine learning approaches will further enhance the interpretation of ΔΔG and its role in variant assessment, especially those of uncertain significance.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1ACGS (2024). ACGS best practice guidelines for variant classification in rare disease. Available online at: https://www.acgs.uk.com/media/12533/uk-practice-guidelines-for-variant-classification-v 12-2024.pdf.
- 2Appelbaum P. S.Burke W.Parens E.Zeevi D. A.Arbour L.Garrison N. A. (2022). Is there a way to reduce the inequity in variant interpretation on the basis of ancestry? Am. J. Hum. Genet. 109 (6), 981–988. 10.1016/j.ajhg.2022.04.012 35659933 PMC 9247826 · doi ↗ · pubmed ↗
- 3Caswell R. C.Gunning A. C.Owens M. M.Ellard S.Wright C. F. (2022). Assessing the clinical utility of protein structural analysis in genomic variant classification: experiences from a diagnostic laboratory. Genome Med. 14 (1), 77. 10.1186/s 13073-022-01082-2 35869530 PMC 9308257 · doi ↗ · pubmed ↗
- 4Chen E.Facio F. M.Aradhya K. W.Rojahn S.Hatchell K. E.Aguilar S. (2023). Rates and classification of variants of uncertain significance in hereditary disease genetic testing. JAMA Netw. Open 6 (10), e 2339571. 10.1001/jamanetworkopen.2023.39571 37878314 PMC 10600581 · doi ↗ · pubmed ↗
- 5Chen S.Francioli L. C.Goodrich J. K.Collins R. L.Kanai M.Wang Q. (2024). A genomic mutational constraint map using variation in 76,156 human genomes. Nature 625, 92–100. 10.1038/s 41586-023-06045-0 38057664 PMC 11629659 · doi ↗ · pubmed ↗
- 6Chen Y.Lu H.Zhang N.Zhu Z.Wang S.Li M. (2020). Prem PS: predicting the impact of missense mutations on protein stability. P Lo S Comput. Biol. 16 (12), e 1008543. 10.1371/journal.pcbi.1008543 33378330 PMC 7802934 · doi ↗ · pubmed ↗
- 7Choudhury A.Mohammad T.Anjum F.Shafie A.Singh I. K.Abdullaev B. (2022). Comparative analysis of web-based programs for single amino acid substitutions in proteins. P Lo S One 17 (5), e 0267084. 10.1371/journal.pone.0267084 35507592 PMC 9067658 · doi ↗ · pubmed ↗
- 8David A.Sternberg M. J. E. (2023). Protein structure-based evaluation of missense variants: resources, challenges and future directions. Curr. Opin. Struct. Biol. 80, 102600. 10.1016/j.sbi.2023.102600 37126977 · doi ↗ · pubmed ↗