Functional analysis of promoter element 2 within the viral polymerase gene of an emerging paramyxovirus, Sosuga virus
Lipi Akter, Junna Kawasaki, Tofazzal Md. Rakib, Takashi Okura, Fumihiro Kato, Shohei Kojima, Kosuke Oda, Yusuke Matsumoto

TL;DR
This study investigates the role of a specific promoter element in the Sosuga virus, revealing its impact on viral replication and the conservation of a protein motif in related viruses.
Contribution
The study demonstrates the functional importance of retaining agPE2 in SOSV minigenomes and identifies a conserved motif in the L protein across Rubulavirinae.
Findings
Including agPE2 in SOSV minigenomes significantly increases RNA synthesis activity.
The agPE2 region encodes a conserved motif at the C-terminus of the L protein in Rubulavirinae.
This motif is preserved even in viruses lacking agPE2 within the L gene.
Abstract
Paramyxovirus genomes carry bipartite promoters at the 3′ ends of both their genome and antigenome, thereby initiating RNA synthesis, which requires the viral polymerase to recognize two elements: the primary promoter element 1 (PE1) and the secondary promoter element 2 (PE2). We have previously shown that the antigenomic PE2 (agPE2) in many viruses in the Rubulavirinae subfamily is located within the coding region of the viral RNA polymerase L gene. Sosuga virus (SOSV), belonging to the Rubulavirinae subfamily, is highly pathogenic to humans, thus necessitating high-level containment facilities for infectious virus research. The use of a minigenome system permits studies of viral RNA synthesis at lower biosafety levels. Because minigenomes of negative-strand RNA viruses generally comprise only the untranslated regions, agPE2 within the L coding region—such as those found in…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
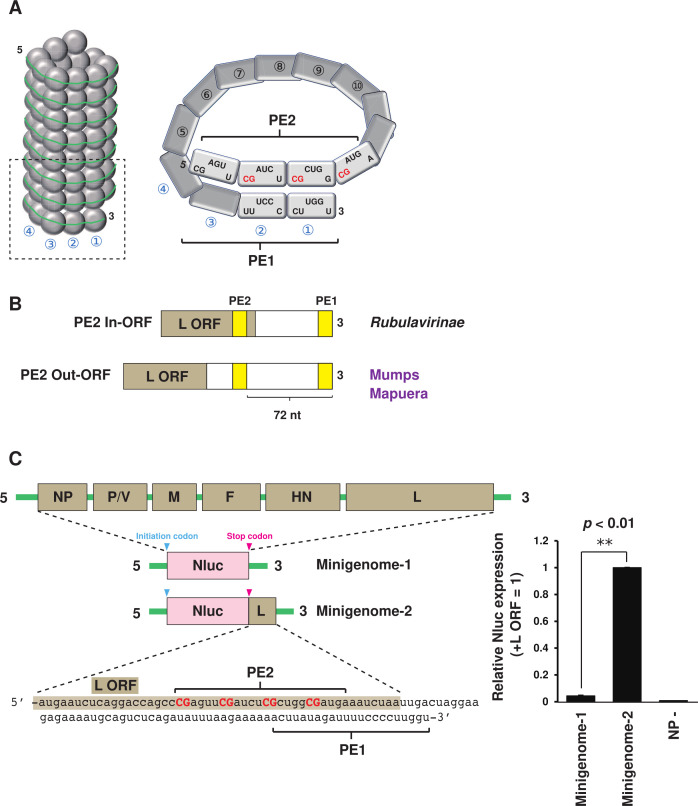 Fig 1
Fig 1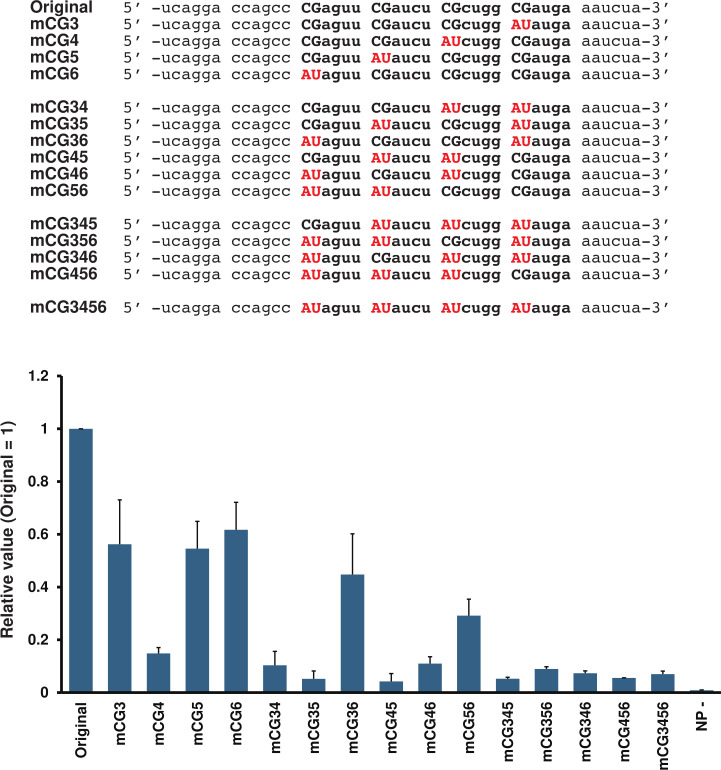 Fig 2
Fig 2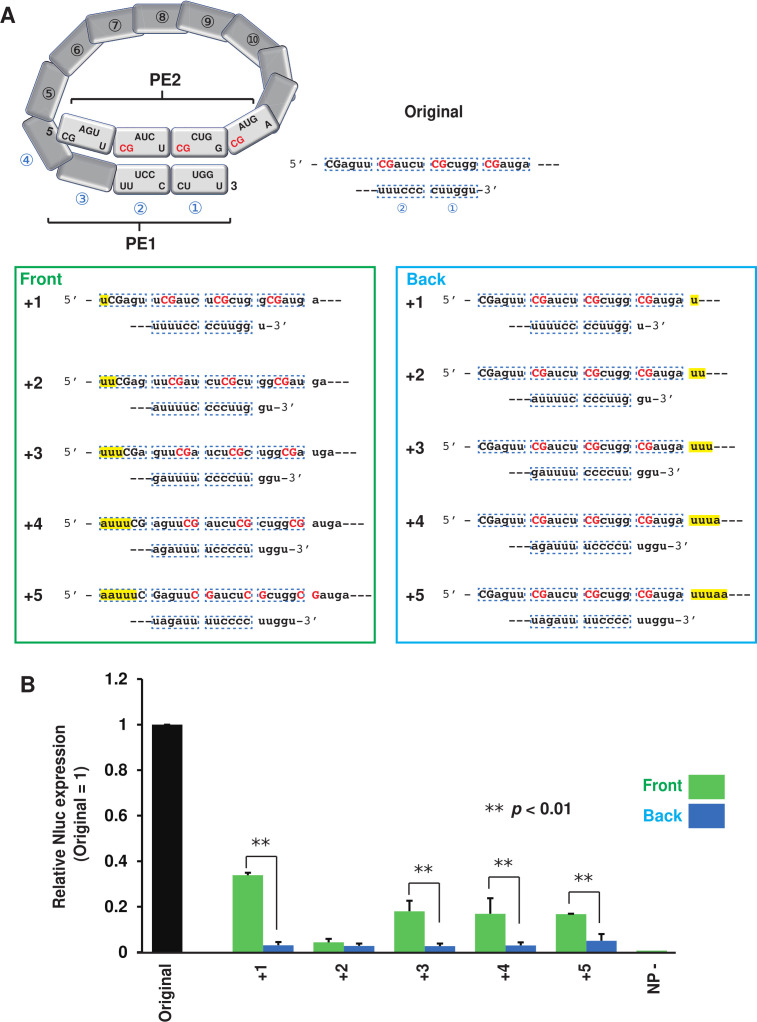 Fig 3
Fig 3 Fig 4
Fig 4 Fig 5
Fig 5 Fig 6
Fig 6- —Japan Agency for Medical Research and Development (AMED)
- —MEXT | Japan Society for the Promotion of Science (JSPS)
- —Takeda Science Foundationhttp://dx.doi.org/10.13039/100007449
- —Kato Memorial Bioscience Foundationhttp://dx.doi.org/10.13039/501100004051
- —Kieikai Research Foundationhttp://dx.doi.org/10.13039/501100012014
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsVirology and Viral Diseases · Viral Infections and Vectors · Plant Virus Research Studies
INTRODUCTION
Paramyxoviruses are single-stranded negative-sense RNA viruses that belong to the order Mononegavirales, which includes many important pathogens of animals and humans (1). The family Paramyxoviridae is mainly divided into the following subfamilies: Orthoparamyxovirinae (measles virus, Nipah virus, and Sendai virus), Avulavirinae (Newcastle disease virus), and Rubulavirinae (parainfluenza virus type 5 [PIV5], human parainfluenza virus type 2 [hPIV2], and mumps virus [MuV]). The negative-strand RNA genome of paramyxoviruses typically encodes about six genes (e.g., hPIV2; 3′-NP-P/V-M-F-HN-L-5′), which are completely encapsidated by the viral nucleoprotein (NP) to form a helical nucleocapsid. This nucleocapsid serves as a template for the viral RNA-dependent RNA polymerase (RdRp) complex, which consists of the large protein (L) and the phosphoprotein (P) cofactor (2).
Viruses belonging to the order Mononegavirales use the 3′ terminus of their genome as a replication promoter (promoter element 1, PE1). In addition, viruses in the family Paramyxoviridae possess a bipartite promoter system that requires a secondary promoter element (PE2) located within the internal region of the genome (Fig. 1A) (3–6). The PE1/2 located at the 3′ end of the negative-strand genome is referred to as the genomic PE and functions in the production of the positive-strand antigenome. Conversely, the PE in the positive-strand antigenome is termed the antigenomic PE (agPE) and is essential for the production of the negative-strand genome. Furthermore, paramyxoviruses follow the “Rule of Six,” which requires that the nucleotide (nt) numbers in the genome should be a multiple of six for efficient viral replication (3, 7). The genomic RNA is coated by NP monomers, with each monomer binding precisely to 6 nts (8–10).
Establishment of Sosuga virus (SOSV) minigenome assay. (A) Helical structure of SOSV nucleocapsid. Promoter element 1 (PE1) and PE2 are shown. (B) Viruses in the Rubulavirinae subfamily are categorized into PE2 In-ORF and Out-ORF types. (C) Schematic diagram of SOSV minigenome construction. All viral genes in the genome are exchanged to be Nluc. Because the SOSV genome contains the PE2 within the L gene, two minigenomes with or without PE2 are created. The relative Nluc expression using the SOSV minigenome assay is shown. Data represent means and standard deviations from three different experiments. NP− indicates the values using an empty plasmid instead of SOSV NP.
Understanding the genome structure and replication mechanisms of paramyxoviruses is crucial for elucidating how these viruses propagate and exert their pathogenicity. Research on highly pathogenic viruses plays a significant role in developing new therapeutic and preventive strategies. Sosuga virus (SOSV) belongs to the genus Pararubulavirus in the subfamily Rubulavirinae (11–14). SOSV was first identified in clinical samples from a wildlife biologist who became infected while collecting bat specimens in South Sudan and Uganda in 2012. The patient exhibited symptoms including fever, generalized myalgia, arthralgia, neck stiffness, and a sore throat. Due to its pathogenicity, experiments using infectious SOSV must be conducted in high-biocontainment facilities at biosafety level (BSL)-4 (or in some cases, BSL-3). As a result, research on SOSV using the authentic virus is restricted, creating a need for genome replication modeling systems that allow for the safe study of the viral life cycle at lower biosafety levels. One such model is the minigenome system, which can be utilized in a BSL-2 environment. The development of an experimental system that efficiently measures the function of RdRp in highly pathogenic paramyxoviruses is required for a detailed study of the molecular basis of genome replication.
RESULTS
An improved SOSV minigenome system reveals the importance of agPE2 for replication
The minigenome system is designed to indirectly quantify viral gene expression by creating a construct in which a reporter gene is flanked by the 3′ and 5′ untranslated regions (UTRs) of the viral genome. When introduced into cultured cells along with the viral polymerase complex, this system enables the assessment of viral gene replication. In the case of SOSV, a minigenome system using Zoanthus green fluorescent protein (ZsGreen1) as a reporter gene has been previously reported (12). In this study, we developed a new system in which the reporter gene of the SOSV minigenome system was replaced with the easily quantifiable Nano Luciferase (Nluc). During the process for minigenome construction, the initiation codon of the first gene NP and the stop codon of the last gene L are replaced with the corresponding codons from ZsGreen1 or Nluc, and the total nucleotide count is adjusted to ensure it remains a multiple of six. The ZsGreen1 version of minigenome created by the process above was reported previously (12). We prepared a Nluc-expressing SOSV minigenome by the method above, termed as the minigenome-1 (Fig. 1C). The minigenome-1 expressing plasmid was transfected into baby hamster kidney (BHK) cells constitutively expressing T7 RNA polymerase (BHK/T7-9 cells) together with the SOSV NP, P, and L expressing helper plasmids, and Nluc expression was examined 48 h post-transfection. The minigenome-1 system showed only approximately five times the activity of the negative control (no NP expression plasmid) (Fig. 1C; minigenome-1). We have previously shown that the agPE2 of Rubulavirinae is located within the L open reading frame (ORF), termed PE2 In-ORF, with the exception of MuV and Mapuera virus, termed PE2 Out-ORF (Fig. 1B) (15). This SOSV minigenome-1 does not retain the correct agPE2 (12). Therefore, we prepared the minigenome-2 by incorporating part of the nucleotides corresponding to the L ORF after the stop codon of the Nluc gene (Fig. 1C; minigenome-2). The minigenome-2 system showed 21 times the activity of the minigenome-1 system and 106 times the activity of the negative control NP−. This shows that agPE2 in the L ORF is essential for the replication activity of SOSV.
Importance of CG repeats in agPE2 for SOSV minigenome replication
The SOSV minigenome-2 system enables us to study the significance of CG repeats in viral replication. Although the three repeated CGnnnn in hexamers 13, 14, and 15 were shown to be important for PIV5 (Rubulavirinae) genome replication, SOSV agPE2 possesses four consecutive CGnnnn repeats. The activity of the minigenome was assessed by systematically replacing the CGs in the four CGnnnn repeats of SOSV agPE2 with AU pairs, 1–4 at a time (Fig. 2). When a single pair of CGs was substituted, the activity dropped to 15% upon replacing hex14 with AU; however, when hex13, hex15, and hex16 were substituted, the activity remained stable at 50%–60% (Fig. 2; mCG3 to mCG6). With two pairs of CG substitutions, it was observed that replacing hex13 and hex16 or hex15 and hex16 had a relatively minimal impact on activity (Fig. 2; mCG34 to mCG56). When three and four pairs were replaced, all substitutions exhibited a significant negative effect (Fig. 2; mCG345 to mCG3456). These results indicate that the CG in hex14 is critical for the SOSV promoter. The relatively high activity observed in the two pairs of mutations involving hex16, when hex14 was omitted (mCG36 and mCG56), suggests that the importance of hex16 may be somewhat lower than that of the other repeats. It is suggested that the three or four CG repeats within agPE2 in SOSV are not equally important, but rather, their significance for genome replication varies.
Importance of CG repeats in PE2 for SOSV genome replication. Mutations in the PE2 sequences are shown in the left panel. The relative Nluc expressions using the SOSV minigenome assay system are shown in the right panel. The relative value when the value using an original minigenome is 1. Data represent means and standard deviations from three different experiments. NP− indicates the value using an empty plasmid instead of SOSV NP.
The relationship between the positions of PE1 and PE2 and their function as replication promoters
The paramyxoviral genome with bipartite promoters is recognized by viral RdRp as a template only when NP-encapsidated PE1 and PE2 adopt a configuration at the 3′ end of the helical nucleocapsid, as depicted in Fig. 3A (3, 4). During genome (or antigenome) replication, the NP encapsidates the newly synthesized genome/antigenome incrementally by 6 nts from the 5′ end (16). If the total number of nucleotides in the genome/antigenome is not a multiple of six, the relative positions of PE1 and PE2 become misaligned. In this case, it is considered that the viral RdRp cannot recognize the correct promoters. This is thought to be a mechanism for RdRp to recognize that the template is a multiple of six. We added 1–5 nts to the front and back of agPE2 in the SOSV minigenome-2, assessing their replication activity when they were not multiples of six (Fig. 3A). In the original state, agPE1 and agPE2 are correctly aligned: 5′-CGcugg-3′ in hex14 is aligned with 5′-cuuggu-3′ in hex1, and 5′-CGaucu-3′ in hex15 is aligned with 5′-uuuccc-3′ in hex2 (Fig. 3A; original). The method of adding 1 to 5 nucleotides to the front of agPE2 produces a 6n + 1 to 6n + 5 minigenome without changing the juxtaposition of nucleotide sequences in [hex14 and hex1] and [hex15 and hex2] (Fig. 3; front +1 to +5). In this case, the SOSV minigenome activity was slightly observed, and in particular, the front +1, +3, +4, and +5 minigenomes showed about 20% of the original activity (Fig. 3B; front). The method of adding 1 to 5 nucleotides to the back of agPE2 changes the order of [hex14 and hex1] and [hex15 and hex2], but the natural nucleotide sequence within the hex13 to hex16 NPs is not changed (Fig. 3; back +1 to +5). In all cases of these mutations, the SOSV minigenome activity resulted in a remarkable loss of activity (Fig. 3B; back). These results suggest that it is not the specific placement of the CG motif within NP that is essential for promoter activity, but rather the preservation of the parallel arrangement between PE1 and PE2—even if the precise positioning of the CG motif within NP is lost.
The replication of SOSV minigenome constructed by nucleotides not a multiple of six. (A) Effect of nucleotide addition in the 5′ or 3′ position of agPE2 for minigenome expression. One to five nucleotide additions (+1 to +5) at the indicated position and their effect for the shift between PE1 and PE2 are shown. (B) The relative value when the value using an original minigenome is 1. Data represent means and standard deviations from three different experiments.
Comparison of the agPE2 sequence of viruses in the subfamily Rubulavirinae. The agPE2 sequence of viruses in the genera Pararubulavirus and Orthorubulavirus are shown. Numbers in the circles indicate the numbers of the NP hexamer from the 3′ terminus. hPIV, human parainfluenza virus; Porcine, Porcine rubulavirus.
Comprehensive analysis of agPE2 of the subfamily Rubulavirinae
To gain more insight into agPE2 function of all other Rubulavirinae, we comprehensively compared the conservation of nucleotides among all Rubulavirinae agPE2 in sequences in the National Center for Biotechnology Information (NCBI) refseq database. The agPE2 region (73–96 nts from the 3′ terminus of the antigenome) was extracted from all sequences, and their conservation among virus species was analyzed using the WebLogo sequence logo generator (https://weblogo.berkeley.edu/logo.cgi) (Fig. 4). There are a total of 12 species of Rubulavirinae whose sequences have been accurately registered in NCBI refseq, including six each from the genera Pararubulavirus (SOSV, Tuhoko, Menangle, Achimota, Teviot, and Tioman viruses) and Orthorubulavirus (hPIV2, PIV5, Porcine rubulavirus, Alston, mumps, and Mapuera viruses). Notably, SOSV, Porcine rubulavirus, and Alston virus exhibit four consecutive CGnnnn repeats within the hexamer numbers 13 to 16 from the 3′ terminal (namely hex13 to hex16) (Fig. 4). In the genus Pararubulavirus, Tuhoko and Achimota viruses, as well as in the genus Orthorubulavirus, PIV5 and MuV, the hex13, hex14, and hex15 exhibit three consecutive CGnnnn repeats. Several other patterns emerged, including hPIV2, which appears to have four consecutive CGnnnn repeats interspersed with a single GGnnnn, and Mapuera virus, characterized by three CGnnnn repeats in hexamers 14, 15, and 16. Overall, our findings indicate the conservation of CGnnnn repeats of three or more in the agPE2. We observed no clear distinctive features in the agPE2 sequences between PE2 In-ORF viruses and PE2 Out-ORF viruses (MuV and Mapuera virus).
Amino acid sequences of L protein at the agPE2 region
The agPE2 region of In-ORF Rubulavirinae serves dual roles as a promoter element and as a codon for amino acids in the L protein. We investigated the specific amino acids encoded by the agPE2 region across all Rubulavirinae L proteins including In-ORF and Out-ORF viruses (Fig. 5A). This region is situated at the C-terminus of the L protein and is characterized by 3–4 CGnnnn repeats, which restrict codon usage. Consequently, as illustrated in Fig. 5B, these amino acid sequences exhibit the D/G-I/L-x-G-B motif (where B denotes D or E). Notably, MuV and Mapuera virus, which are PE2 Out-ORF viruses, lack agPE2 in the L ORF, but the amino acid sequences at the same position of the C-terminus of the L protein have the same D/G-I/L-x-G-B motif as In-ORF Rubulavirinae.
Amino acid sequences of PE2 encoding region in the RNA-dependent RNA polymerase gene. (A) Comparison of nucleotide and amino acid sequences of PE2 of viruses in the genera Pararubulavirus and Orthorubulavirus. (B) Homology of amino acid sequences of the L protein analyzed using WebLogo sequence logo generator. (C) Predicted model of P protein-bound SOSV L protein. RdRp, RNA-dependent RNA polymerase (colored in light blue); PRNTase, polyribonucleotidyltransferase (colored in green); CD, connecting domain (colored in orange); MTase, methyltransferase (colored in yellow); CTD, C-terminal domain (colored in blue). In an enlarged view of the C-terminal loop, carbon atoms of which are colored in pink, residues from Asp2266 to Asp2270 are shown in stick diagrams. P protein tetramer bound to L protein are colored purple. (D) The SOSV minigenome-2 system was used to study the effect of amino acid substitution in the D2265LAGD2269 of SOSV L. Minigenome-2 and helper plasmids containing SOSV NP, P, and L (normal or mutant) were transfected to BHK/T7-9 cells. The relative Nluc expression when L normal is set to 1 is shown. Data represent means and standard deviations from three different experiments. L− indicates the values using an empty plasmid instead of SOSV L.
We constructed a predicted structural model of the SOSV L protein to visualize the position of the D_2265_LAGD_2269_ motif within the SOSV L context (Fig. 5C). The structure of the C-terminal loop containing the D_2265_LAGD_2269_ motif is relatively less reliable, perhaps due to protruding into the solvent region. In the model, this D_2265_LAGD_2269_ motif was bound to the concave on the molecular surface, which is surrounded by methyltransferase (MTase) domain, C-terminal domain, and connecting domain.
To assess the functionality of the D_2265_LAGD_2269_ motif of SOSV L protein, we replaced it with A_2265_AAAA_2269_ and used it as a helper plasmid for the SOSV minigenome-2 system (Fig. 5D). In this experiment, we used an original SOSV minigenome-2 (no mutation in its agPE2) as a template and transfected it with helper plasmids including SOSV NP, P, and L (normal or mutant). However, the A_2265_AAAA_2269_ mutation did not affect polymerase activity (Fig. 5D). Therefore, it appears that the SOSV L D_2265_LAGD_2269_ motif may not play a significant role in polymerase function, at least in the SOSV minigenome-2 system.
DISCUSSION
For studying the genome replication mechanism of highly pathogenic negative-strand RNA viruses such as Ebola, Marburg, and Nipah viruses, the minigenome systems have been used (17–20). In the process of developing a minigenome system for pathogenic SOSV using Nluc as a reporter, we have shown that the minigenome activity increased by more than 20 times simply depending on the presence of agPE2 within the L ORF (Fig. 1C). Due to this high minigenome activity, we were able to analyze the properties of agPE2 for In-ORF type Rubulavirinae.
The paramyxovirus promoter operates only when NP-encapsidated PE1 and PE2 at the 3′ terminus of the helical nucleocapsid are properly aligned, as shown in Fig. 1A (3, 4, 21). In PIV5 and hPIV2, the repeated CG sequence within hexamers 13, 14, and 15 plays a crucial role in the agPE2 region (21–23). It was thought the specific positioning of the 5′-CG-3′ sequence within NP monomers (with C as the first and G as the second in 5′-CGnnnn-3′)—referred to as “phasing”—was critical for promoter activity of these Rubulavirinae (22, 23). However, our results using the SOSV minigenome-2 system demonstrated that even if the 5′-CG-3′ motif is not positioned at the first or second nucleotide of hexamers 13 through 16, the polymerase still exhibits a certain level of activity, provided that the nucleotide sequences of PE1 and PE2 are properly juxtaposed (Fig. 3B; front). Conversely, even when the 5′-CGnnnn-3′ sequence is correctly positioned within NP monomers in hexamers 13 through 16, promoter activity is lost if the alignment between PE1 and PE2 is disrupted (Fig. 3B; back). Thus, we found that even if the position of the 5′-CG-3′ motif within NP monomers is inaccurate, promoter function is maintained at a certain level of activity as long as the nucleotide sequences corresponding to [hex14 and hex1] and [hex15 and hex2]—e.g., hex14 (5′-CGcugg-3′) and hex1 (5′-cuuggu-3′)—are juxtaposed. In nature, when PE1 and PE2 sequences act as a promoter for genome replication, their nucleotide sequences are encapsidated by NP. Nevertheless, the juxtaposition of these two sequences is critical for promoter function. Further structural analysis is required to understand how the L protein recognizes the complex promoter structure formed by PE1 and PE2. Moreover, to understand the functions of PE1 and PE2, comprehensive studies are needed not only on SOSV but also on PIV5, hPIV2, and other viruses belonging to the genera Orthorubulavirus and Pararubulavirus.
In the Rubulavirinae subfamily, the agPE2 region contains three or four consecutive CGnnnn repeats (Fig. 4). To date, only three Rubulavirus species have been found to harbor four such repeats in their agPE2, and SOSV is the sole member of the Pararubulavirus genus (Fig. 4). Notably, our data showed that this fourth CG repeat is functional for minigenome promoter (Fig. 2), indicating that viruses bearing four repeats may gain a distinct advantage over those with only three.
A distinguishing feature of Rubulavirinae compared to Orthoparamyxovirinae and Avulavirinae is their different RNA editing mode (24). Paramyxoviruses utilize an RNA editing mechanism within the P/V gene, which encodes two proteins: the P protein, an essential polymerase cofactor for viral replication, and the V protein, an accessory protein that interacts with multiple host cellular proteins and acts as an interferon antagonist (25, 26). During mRNA transcription, the viral RdRp recognizes a cis-acting RNA editing signal within the P/V gene, and non-template-dependent nucleotides are co-transcriptionally inserted into the newly synthesized mRNA, allowing the synthesis of both P and V proteins from a single P/V gene (27–29). In many Orthoparamyxovirinae and Avulavirinae, the P protein is produced when RNA editing does not occur, while the V protein is generated when RNA editing occurs (referred to as “V-mode viruses”). In contrast, in viruses belonging to the subfamily Rubulavirinae (including MuV and Mapuera virus), the opposite occurs: V protein is produced when RNA editing does not occur, and P protein is synthesized when RNA editing occurs (known as “P-mode viruses”). Interestingly, P-mode viruses are also found outside the Rubulavirinae subfamily, such as Fer-de-lance virus (Orthoparamyxovirinae) (30), Anaconda paramyxovirus (Orthoparamyxovirinae) (31), and avian paramyxovirus 11 (Avulavirinae) (32). These viruses generate P proteins through RNA editing but belong to the “PE2 Out-ORF” type, where the agPE2 is located outside the L ORF, a feature they share with MuV and Mapuera virus. Therefore, the presence of agPE2 within the L ORF is not a strict requirement for a virus to adopt the P-mode. To date, no V-mode virus with an In-ORF type has been discovered, suggesting that if these two characteristics are indeed mutually exclusive, understanding the underlying mechanism could shed light on why paramyxoviruses have evolved to adopt either P-mode or V-mode, as well as In-ORF or Out-ORF configurations.
As repeatedly mentioned, with the exception of MuV and Mapuera virus, Rubulavirinae possesses an agPE2 sequence within the L ORF, where the sequence “CGnnnn” is repeated at least three times (Fig. 5A) (15). This agPE2 sequence imposes strong codon selection constraints, leading to the semi-essential presence of the “D/G-I/L-G-B motif” at the C-terminus of the L protein (Fig. 5A and B). However, in the experiment using the SOSV minigenome-2 system, we observed that mutations in the corresponding “D_2265_LAGD_2269_ sequence” of this motif do not affect polymerase activity (Fig. 5D). While we cannot conclude that this motif is essential for viral replication, we cannot dismiss the possibility that it played some functional roles in viral evolution. From an evolutionary standpoint, it is likely that MuV and Mapuera virus evolved as PE2 In-ORF viruses, like many other viruses in the Rubulavirinae. Phylogenetic analysis of the L protein, as shown in Fig. 6A, suggests that MuV and Mapuera virus are not direct ancestors of Rubulavirinae and are unlikely to have emerged as a new independent lineage. Instead, it is more plausible that the PE2 In-ORF type virus appeared first in Rubulavirinae, and the PE2 Out-ORF type viruses, such as MuV and Mapuera virus, emerged later. Moreover, it is likely that these two viruses are not independent evolutionary lineages, but rather variants of PE2 In-ORF Orthorubulavirus genus that incidentally arose. The D/G-I/L-G-B motif likely played a critical role in this evolutionary process, which may explain why it remains conserved in MuV and Mapuera virus. Then, how did the L proteins of MuV and Mapuera virus arise? Their genomes and L amino acid sequences are not particularly long compared to other Rubulavirinae (Fig. 6B). Therefore, it seems more likely that the agPE2 was originally present within the L ORF of an ancestral virus (Fig. 6C; ancestral rubulaviruses). Over time, the C-terminus of the L protein in these viruses may have been lost, and despite the loss of the agPE2 sequences (Fig. 6C; truncation of L C-terminus), the virus could have evolved to reacquire the D/G-I/L-G-B motif (Fig. 6C; re-acquisition of D/G-I/L-G/B motif). This suggests that the D/G-I/L-G-B motif was crucial for Rubulavirinae and needed to be reacquired even after being lost. Because this motif is absent in the Orthoparamyxovirinae and Avulavirinae subfamilies, understanding its functional significance will be key to uncovering the unique features of the Rubulavirinae subfamily within the Paramyxoviridae family. In the structural model we constructed, the SOSV D_2265_LAGD_2269_ motif was found to be positioned in close proximity to a domain involved in RNA synthesis (e.g., the MTase domain), suggesting its involvement in gene expression (Fig. 5C). However, it is also possible that this motif is associated with other functions, such as virion assembly or interaction with host factors to improve the efficiency of viral propagation.
Predicted evolutionary history of Rubulavirus. (A) Phylogenetic tree created from an alignment of the viral L protein of Rubulavirinae. (B) The length of genome nucleotides and L protein amino acids. The GenBank accession numbers used in this study are Mapuera (NC_009489.1), Porcine (NC_009640.1), mumps (NC_002200.1), PIV5 (KY685075.1), hPIV2 (AB176531.1), Alston (NC_055508.1), Menangle (AF326114.2), Sosuga (KF774436.1), Achimota (JX051319.1), Teviot (MH708896.1), Tioman (AF298895.2), and Tuhoko (GU128080.1). (C) Predicted evolutionary history of PE2 Out-ORF Rubulavirinae.
Currently, experiments using the SOSV minigenome-2 system (Pararubulavirus genus) have shown the importance of fourth CGnnnn for genome replication (Fig. 2) and that the D/G-I/L-G-B motif does not affect polymerase activity (Fig. 5D). However, comparative studies with other Rubulavirinae, such as Orthorubulavirus genus, may yield different results. Comprehensive comparative experiments would provide new insights into the differences between Pararubulaviruses and Orthorubulaviruses genera. Such research may further clarify the characteristics of SOSV, the only Pararubulavirus known to exhibit high pathogenicity in humans.
MATERIALS AND METHODS
Cells
Baby hamster kidney (BHK) cells constitutively expressing T7 RNA polymerase (BHK/T7-9 cells) (33) were cultured in Dulbecco’s modified Eagle’s medium with 5% fetal calf serum and penicillin/streptomycin. Cells were cultured at 37°C in 5% CO_2_.
Plasmid construction
The Nluc-expressing SOSV minigenome plasmid (SOSV-Nluc) was constructed using the pUC57 plasmid backbone. The Nluc gene was flanked by the 3′ UTR containing the leader sequence and the 5′ UTR containing the trailer sequence of the SOSV genome (GenBank accession number: NC_025343.1). The minigenome is set under the control of the T7 RNA polymerase promoter, and the transcript expressed as a negative sense RNA is cleaved at both ends by a hammerhead ribozyme and a hepatitis delta virus ribozyme (34). The SOSV NP, P, and L genes are set into the cloning site of the pCAGGS vector. All SOSV plasmids were generated by GenScript’s custom gene synthesis services (GenScript Japan, Tokyo, Japan). A pCAGGS-derivative plasmid for the expression of firefly luciferase (Fluc) was also constructed (15). The mutations for the agPE2 in SOSV-Nluc were introduced by a standard cloning method.
SOSV Nluc minigenome assay
The SOSV Nluc minigenome assay was performed in BHK/T7-9 cells cultured in 12-well plates (1 × 10^5^ cells/well). Plasmids including SOSV-Nluc (0.5 µg), pCAGGS-L (0.4 µg), pCAGGS-P (0.4 µg), and pCAGGS-NP (0.4 µg) or an empty vector, and Fluc (0.05 µg) were transfected using XtremeGENE HP (Merck, Darmstadt, Germany). At 48 h post-transfection, the cells were lysed with passive lysis buffer (Promega), and the Nluc and Fluc activities were measured using the Nano-Glo Dual-Luciferase Reporter Assay System (Promega, Madison, WI, USA) according to the manufacturer’s instructions. All activity values measured for Nluc were normalized to the expression levels of Fluc.
Comparative analysis of Rubulavirinae PE2
Sequences of viruses in the subfamily Rubulavirinae were downloaded from the NCBI refseq database as previously reported (15). For each sequence, we extracted the sequences of the agPE2 (73–96 nts from the 3′ terminus of the antigenome). The sequence conservation was analyzed using the WebLogo sequence logo generator (https://weblogo.berkeley.edu/logo.cgi).
AlphaFold predictions
The AlphaFold models of SOSV L protein were predicted using the AlphaFold version 2.0 algorithm on the Colab server (https://colab.research.google.com/github/sokrypton/ColabFold/blob/main/AlphaFold2.ipynb), accessed on 25 July 2024 (35). Predictions were performed with default multiple sequence alignment generation using the MMSeqs2 server, with 48 recycles and templates (homologous structures). The complex structure of L protein and P protein tetramer was generated based on the best out of L protein AlphaFold models (rank 1) and the structure of mumps virus L protein bound by P protein tetramer (PDB code 8IZL). The figures were drawn using PyMOL software (36).
Phylogenetic tree of Rubulavirinae
Alignment and phylogenetic reconstructions were performed using the function “build” of ETE3 3.1.3 (37) as implemented on the GenomeNet (https://www.genome.jp/tools/ete/). ML tree was inferred using PhyML v20160115 ran with model and parameters: --pinv e --alpha e --nclasses 4 -o tlr -f m --bootstrap -2 (38). Branch supports are the χ^2^-based parametric values returned by the approximate likelihood ratio test.
Statistical analysis
A one-way analysis of variance (ANOVA) was performed, followed by Tukey’s honest significant difference (HSD) test for post hoc pairwise comparisons. The ANOVA was conducted using the aov() function, and the subsequent Tukey’s HSD test was performed with the TukeyHSD() function in R (version 4.4.0, R Foundation for Statistical Computing, Vienna, Austria). Statistical significance was assigned when P values were <0.05.
Supplementary Material
Reviewer comments
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Duprex WP, Dutch RE. 2023. Paramyxoviruses: pathogenesis, vaccines, antivirals, and prototypes for pandemic preparedness. J Infect Dis 228:S 390–S 397. doi:10.1093/infdis/jiad 12337849400 PMC 11009463 · doi ↗ · pubmed ↗
- 2Whelan SPJ, Barr JN, Wertz GW. 2004. Transcription and replication of nonsegmented negative-strand RNA viruses. Curr Top Microbiol Immunol 283:61–119. doi:10.1007/978-3-662-06099-5_315298168 · doi ↗ · pubmed ↗
- 3Kolakofsky D, Roux L, Garcin D, Ruigrok RWH. 2005. Paramyxovirus m RNA editing, the “rule of six” and error catastrophe: a hypothesis. J Gen Virol 86:1869–1877. doi:10.1099/vir.0.80986-015958664 · doi ↗ · pubmed ↗
- 4Kolakofsky D. 2016. Paramyxovirus RNA synthesis, m RNA editing, and genome hexamer phase: A review. Virology 498:94–98. doi:10.1016/j.virol.2016.08.01827567257 · doi ↗ · pubmed ↗
- 5Gutsche I, le Mercier P, Kolakofsky D. 2020. A paramyxovirus-like model for Ebola virus bipartite promoters. P Lo S Pathog 16:e 1008972. doi:10.1371/journal.ppat.100897233152032 PMC 7643936 · doi ↗ · pubmed ↗
- 6le Mercier P, Kolakofsky D. 2019. Bipartite promoters and RNA editing of paramyxoviruses and filoviruses. RNA 25:279–285. doi:10.1261/rna.068825.11830587495 PMC 6380270 · doi ↗ · pubmed ↗
- 7Calain P, Roux L. 1993. The rule of six, a basic feature for efficient replication of Sendai virus defective interfering RNA. J Virol 67:4822–4830. doi:10.1128/JVI.67.8.4822-4830.19938392616 PMC 237869 · doi ↗ · pubmed ↗
- 8Alayyoubi M, Leser GP, Kors CA, Lamb RA. 2015. Structure of the paramyxovirus parainfluenza virus 5 nucleoprotein-RNA complex. Proc Natl Acad Sci U S A 112:E 1792–9. doi:10.1073/pnas.150394111225831513 PMC 4394319 · doi ↗ · pubmed ↗