Eumachiabrevipedunculata (Rubiaceae, Palicoureeae), a new species from Yunnan, China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1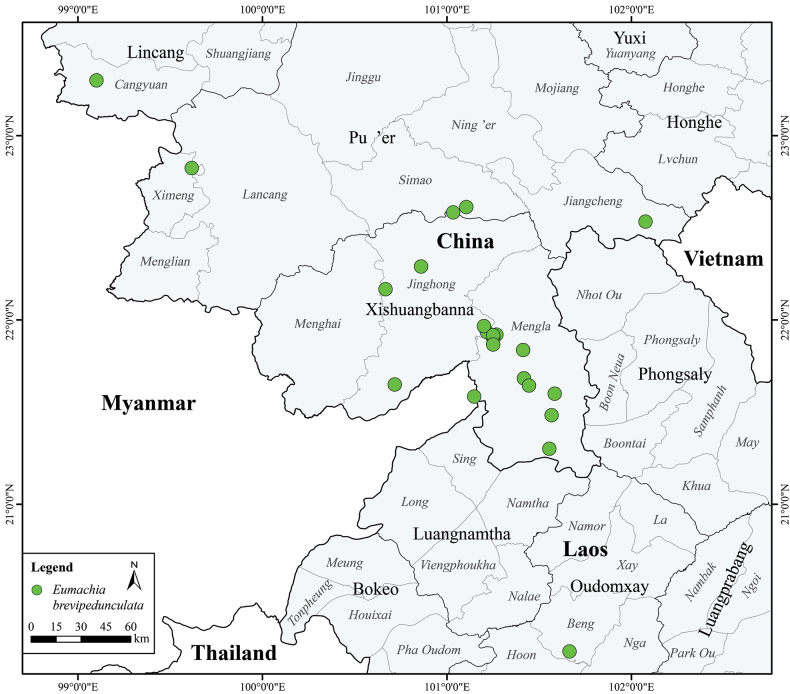 Figure 2
Figure 2 Figure 3
Figure 3| Characters |
|
|
|---|---|---|
| Stipule length | 2 mm | 2.5–6 mm |
| Petiole length | 0.5–1.2 cm | 1–2 cm |
| Leaf size | 5–18 × 2–8 cm | 10–25 × 4–10 cm |
| Inflorescence size | 1–1.2 cm in diameter | 1–4 × 1–2.5 cm |
| Length of inflorescence axes | ca. 0–0.3 cm | 0.3–1 cm |
| Peduncle length | up to 5 mm | 1–1.5 cm |
| Pedicel length | up to 1.5 mm | 1.5–4 mm |
| Calyx lobe length | 0.5–1.5 mm | up to 0.5 mm |
| Calyx lobe shape | linear-lanceolate to narrowly ligulate, apex obtuse | denticulate |
| Corolla colour | yellowish-green | white to cream |
| Corolla tube length | 1–1.5 mm | 1.5–2 mm |
| Anther length | ca. 0.5 mm | ca. 1 mm |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant and Fungal Species Descriptions · Plant Diversity and Evolution · Ethnobotanical and Medicinal Plants Studies
Introduction
The pantropical genus Eumachia DC. (Rubiaceae, Palicoureeae) is characterised by the following characteristics: raphides in the tissues; shrub or small tree habit; smooth branchlets; flattened young shoots and young internodes; opposite leaves, drying yellowish-green; interpetiolar stipules that are persistent or fall by fragmentation, usually united around the stem, generally glandular when young, but becoming indurated and yellowed to ochre when old; terminal inflorescences with green to white axes and typically short bracts; white to cream or yellowish-green corollas with valvate aestivation; usually barbate throats; pyrenes adaxially plane to concave, with marginal preformed germination slits and lacking ethanol-soluble pigments; non-ruminate endosperm (Andersson 2001; Taylor 2005; Barrabé et al. 2012; Taylor et al. 2017). In contrast, its close relative Psychotria L. features vegetative parts drying reddish-brown to grey, deciduous stipules and pyrenes containing ethanol-soluble pigment (Andersson 2001; Davis et al. 2001; Barrabé et al. 2012; Taylor et al. 2017; Razafimandimbison and Rydin 2024).
The genus Eumachia was established in 1830, with the type species being E.carnea (G.Forst) DC. (de Candolle 1830). Since then, the genus has been treated as a synonym of Psychotria (Robbrecht 1988, 1993), though the characteristics of E.carnea do not match the latter. More recently, studies based on molecular phylogeny and morphology resurrected the genus Eumachia and transferred it from the tribe Psychotrieae to Palicoureeae (Barrabé et al. 2012; Barrabé and Davis 2013; Applequist 2014; Razafimandimbison et al. 2014). Some comprehensive combinations and treatments were subsequently made, expanding the circumscription of Eumachia (Barrabé et al. 2013; Delprete and Kirkbride 2015; Taylor et al. 2017). So far, 86 species have been reported in the genus, ranging from Palaeotropical to Neotropical (POWO 2025). Only four species are known from continental Asia (Taylor et al. 2017; Turner 2018). Two species E.montana (Blume) I.M.Turner and E.straminea (Hutch.) Barrabé are currently known in China (Wilson 1916; Chen and Taylor 2011; Taylor et al. 2017; Bijmoer et al. 2024).
Through our study of Psychotria in China, we identified a species extensively collected across southern Yunnan and Laos. However, it does not correspond to any previously recognised taxa. A thorough morphological analysis and consultations of relevant literature and specimens indicate that this species shares characteristics with Eumachia and is new to science. While it is similar to E.straminea in features such as its glabrous stems, papery (when dried) and elliptic-lanceolate leaf blades with 6–8 pairs of secondary veins and terminal inflorescences, it can be distinguished by its smaller stipules, leaf blades and inflorescences and by differences in the floral parts, as outlined in the conclusions below.
We described and illustrated the new Eumachia species here, including formal nomenclatural and morphological details, an explanation of the epithet, selected photographs, an overview of its habitat and phenology and a comparison with similar species that may be confused. Additionally, we included data on studied specimens, a preliminary conservation assessment following the IUCN Red List Categories and Criteria (IUCN 2012; IUCN Standards and Petitions Committee 2024) and notes on its distinctive morphological characteristics and ecological traits.
Materials and methods
The materials studied were derived from living plants collected in the field and herbarium specimens deposited at Xishuangbanna Tropical Botanical Garden (HITBC). To identify the morphological differences with related species, we examined specimens from the Herbaria K, MO, NY, P, IBK, IBSC, LE, A, E etc., through online databases including JSTOR Global Plants (https://plants.jstor.org/), CVH (https://www.cvh.ac.cn/), NSII (http://www.nsii.org.cn/) and GBIF (https://www.gbif.org/). Descriptions were based on morphological observations and measurements of living plants and herbarium specimens.
Taxonomy
Eumachia
brevipedunculata
Taxon classificationPlantaeGentianalesRubiaceae
Y.H.Tan & D.L.Quan sp. nov.
95C4A7A6-1571-5E1D-8F36-ECC53A6B99F2
urn:lsid:ipni.org:names:77360587-1
Type.
China • Yunnan Province: Xishuangbanna Dai Autonomous Prefecture, Mengla County, Menglun Township, Xishuangbanna Tropical Botanical Garden, tropical rain forest, 21°54′52.51″N, 101°15′25.92″E, 579 m a.s.l., 13 May 2024, flowering, D.L.Quan & Y.H.Tan TYH3288 (holotype, HITBC0115875!; isotypes, HITBC0115876!, HITBC0115877!).
Eumachiabrevipedunculata Y.H.Tan & D.L.Quan A habitat B habit C stipules D leaves E flower showing calyx, hypanthium and corolla F inflorescence G front view of long-styled flower H interior view of longitudinally opened long-styled flower I abaxial and adaxial surface of stipule J inflorescence K front view of short-styled flower L interior view of longitudinally opened short-styled flower M front view of calyx lobes and disc N abaxial and adaxial view of longitudinally opened calyx O longitudinal section through hypanthium, disc and calyx P abaxial view of opened corolla Q infructescence R fruits at different developmental stages S cross-section of fruits, showing pyrenes and seeds T ventral (left) and dorsal (right) view of pyrenes. Photographs by Dong-Li Quan.
Diagnosis.
Eumachiabrevipedunculata is morphologically similar to E.straminea, but can be distinguished by its smaller stipules (ca. 2 mm vs. 2.5–6 mm long), shorter petioles (0.5–1.2 cm vs. 1–2 cm long), smaller leaf blades (5–18 × 2–8 cm vs. 10–25 × 4–10 cm); smaller inflorescences (1–1.2 × 1–1.2 cm vs. 1–4 × 1–2.5 cm) without developed axes (vs. axes 3–10 mm long), shorter peduncles (up to 0.5 cm vs. 1–1.5 cm long), shorter pedicels (up to 1.5 mm vs. 1.5–4 mm long); longer calyx lobes (0.5–1.5 mm vs. up to 0.5 mm long), linear-lanceolate to narrowly ligulate calyx lobes (vs. denticulate), shorter corolla tubes (1–1.5 mm vs. 1.5–2 mm long), yellowish-green corolla (vs. white to cream) and shorter anthers (ca. 0.5 mm vs. ca. 1 mm long).
Description.
Shrubs, 0.5–2.5 m tall, branched; stems terete, glabrous; internodes flattened. Leaves opposite, without domatia; petiole 0.5–1.2 cm long, glabrous; leaf blade concavo-convex, elliptic to lanceolate, 5–18 × 2–8 cm, green, often paler abaxially, adaxially somewhat shiny in life, drying papery, glabrous on both surfaces, base cuneate to attenuate, apex acute to acuminate, margins flat or slightly undulant, usually thinly revolute; secondary veins 6–8 at each side of the mid-rib, free or forming a weakly- to a well-developed looping submarginal vein, adaxially costa thickened to prominent and secondary veins sometimes prominent, abaxially costa and secondary veins prominent, remaining venation flat; stipules persistent or falling by fragmentation with persistent portion becoming indurated and yellow to ochre, interpetiolar to shortly fused around stem or forming a sheath, ca. 2 mm long, triangular to ovate, with 2 costae bearing 2 lobes, lobed to 1/4–1/2, lobes subulate with glandular tip, abaxially glabrous, adaxially at base with well-developed drying red-brown colleters. Inflorescences terminal, cymose, congested, subglobose to corymbiform, 1–1.2 × 1–1.2 cm, branched to 1–3 orders without developed secondary axes, generally 3- to 15-flowered, glabrous, subsessile to shortly pedunculate with peduncle up to 5 mm long; bracts narrowly triangular, ca. 0.5 mm long, pubescent to glabrescent outside, apex obtuse; pedicels up to 1.5 mm long. Flowers 5-merous, rarely 4- or 6-merous, usually bent down, distylous; hypanthium obconic, ca. 1.5 mm long, glabrous; calyx green to brown, glabrous, limb ca. 0.5 mm long, lobes 0.5–1.5 mm long, lobes linear-lanceolate to narrowly ligulate, apex obtuse; corolla yellowish-green, campanulate, tube 1.5–2.5 mm in diameter, 1–1.5 mm long and slightly shorter than half the corolla length, glabrous outside, inside white villous in throat, lobes valvate in bud, ovate, 2–2.5 × 1–1.5 mm, adaxially rostrate, abaxially smooth, apex obtuse, bent inwardly; stamens glabrous, in long-styled form 1.5–2 mm long, slightly exserted from corolla tube, in short-styled form 2.5–3 mm long, completely exserted from corolla tube; anthers elliptic to oblong, obtuse, dorsifixed, ca. 0.5 mm long, always exserted from corolla tube; filaments in long-styled form 1–1.5 mm long, in short-styled form 2–2.5 mm long; stigmas 2-lobed, lobes ca. 0.2 mm long; style erect, clavate; style and stigma in long-styled form 2–2.5 mm long, can exceed 1/2 of the corolla tube, but is never exserted from the corolla tube, in short-styled form 1.5–2 mm long, equal in length to or partially exserted from corolla tube; ovary inferior, 2-celled, ovules 1 in each cell, disc glabrous. Fruit drupaceous, fleshy, ellipsoid to subglobose, 0.6–1 cm in diameter, stipitate, glabrous, black at maturity, not reddening, ribbed when dried, calyx lobes persistent; pyrenes 2, hemispherical, bony, dorsally convex with granulose ornamentation or 1–5 shallowly ribbed, ventrally plano-concave, with marginal preformed germination slits, without ethanol-soluble pigments; seeds ellipsoid to hemispherical, 4.5–6 × 5–7 mm; endosperm fleshy, non-ruminate.
Phenology.
Flowering from April to July, fruiting from June to April.
Etymology.
The specific epithet ‘brevipedunculata’ refers to the new species’ distinguishing feature of reduced inflorescences with short peduncles, axes and pedicels. Its Chinese name is given as 短序肉沛木 (Pinyin: duǎn xù ròu pèi mù).
Habitat and distribution.
This species thrives in tropical rainforests and humid evergreen broadleaf forests at 500–1200 m a.s.l. It has been collected from 23 locations in Yunnan and one site in Laos (Fig. 2).
Distribution of Eumachiabrevipedunculata Y.H.Tan & D.L.Quan.
Preliminary IUCN Conservation status.
According to the current distribution of Eumachiabrevipedunculata, the species is found in 23 locations in Yunnan and Laos. Eumachiabrevipedunculata relies heavily on the formal conservation of primary habitat. More than 10 locations are situated within well-protected nature reserves or rainforests. However, some populations are located at the edge of these reserves or near roads, making them vulnerable to threats such as vegetation clearance, destruction by livestock or construction activities. The subpopulations are dispersed and relatively small in size and area, with an estimated total population of about 4,500 individuals. The extent of occurrence (EOO) for E.brevipedunculata is 41,638 km^2^ and the area of occupancy (AOO) is 88 km^2^, as calculated by ShinyGeoCAT (Moat et al. 2023). Based on the IUCN Red List Categories and Criteria (IUCN 2012), notably on the number of locations and the EOO, this species qualifies for classification as Least Concern (LC).
Notes.
Eumachiabrevipedunculata features persistent or marcescent stipules, preformed germination slits, lacks ethanol-soluble pigments in the pyrenes and has a yellowish-green drying colour, which are general characteristics of the tribe Palicoureeae. In Eumachia, fruits are typically orange to red when ripe; however, the fruits of E.brevipedunculata and E.straminea are black. However, colourful fruits, for example, red, blue, white or black, are known to occur in Palicoureeae. Eumachia is not easy to diagnose morphologically because some characteristics were found to be widely variable, such as the stipule type (fused into a tube), the stem texture (smooth or corky), the structure of the inflorescences (lax to subcapitate), the shape of the pyrenes (abaxially smooth to ribbed and adaxially plano-concave to having one or two longitudinal grooves), the type of endosperm (entire to variously ruminate) (Andersson 2001, 2002; Razafimandimbison et al. 2014; Taylor et al. 2017; Taylor 2020; Santos et al. 2021). Moreover, the yellowish-green drying colour of the vegetative parts is generally regarded as a typical and diagnostic characteristic of Eumachia contrasted with reddish-brown to grey in Psychotria (Barrabé et al. 2012; Taylor et al. 2017). However, some studies considered that the colour of the dried leaves varies considerably with how the specimen was prepared and conserved (Taylor 2020; Taylor et al. 2023). All these variations increase the difficulty of intergeneric and interspecific identification of Eumachia. While these characteristics may be homoplasious and not necessarily diagnostic, they remain taxonomically useful (Taylor 2020). Additional taxonomically informative features of Eumachia include chemical constituents, the development of tertiary and quaternary leaf venation, leaf anatomy and the morphology of domatia and stomata (Taylor 2016; Berger et al. 2022).
Eumachiabrevipedunculata is widely distributed in southern Yunnan, China. It has long been misidentified as a Psychotria species, for example, as P.straminea (synonym of E.straminea), P.henryi H.Lév., P.asiatica L., P.siamica Hutch. and so on. It is most similar to E.straminea (Fig. 3), but differs by its smaller stipules and leaf blades, shorter petioles, reduced inflorescences and differences in the floral parts such as longer calyx lobes, shorter corolla tubes and anthers and yellow-green (vs. white-cream) flowers (Table 1). The new Eumachia species is often confused with P.henryi because of the terminal inflorescences, the subcapitate or congested cymose and subsessile inflorescences, but it is distinct by the broader and glabrous leaf blades, the abaxially glabrous, persistent stipules, the undeveloped inflorescence axes, the smaller bracts, the glabrous calyx, the yellowish-green and campanulate corolla, the ovate and larger corolla lobes and the green-black and smaller fruits.
Eumachiastraminea (Hutch.) Barrabé, C.M.Taylor & Razafim A, B habit C stipule D infructescence E inflorescence F inflorescence of long-styled flowers in lateral view G inflorescence of long-styled flowers in front view H inflorescence of short-styled flowers in front view. Photographs by Xiao-Wen Liao (A–C, E–H) and Xiao-Dong Zeng (D).
Additional specimens examined (paratypes).
China • Yunnan Province: Xishuangbanna Dai Autonomous Prefecture: Mengla County: rainforest, 21°29′N, 101°34′E, 800 m a.s.l., 29 May 1982, flowering, expedition team 32655 (HITBC0002822) • Mengla Township: Bubeng Village, 21°36′N, 101°35′E, 720 m a.s.l., 27 April 1982, flowering, expedition team 31873 (HITBC0002825) • Menglun Township: forestry farm, 21°55′N, 101°15′E, 12 May 1975, flowering, P.Z.Zhu 12612 (HITBC0002833) • in the Menglun sub-reserve of Xishuangbanna National Nature Reserve, 21°56′N, 101°13′E, 650 m a.s.l., 11 August 1975, fruiting, G.D.Tao 13730 (HITBC0002839) • Xiaola highway 53 km, dry open forest on the mountaintop, 21°58′N, 101°12′E, 900 m a.s.l., 22 October 1973, fruiting, P.Z.Zhu 10402 (HITBC0002834) • Qixiang, dry forest, 21°55′N, 101°15′E, 1200 m a.s.l., 23 May 1961, flowering, Y.H.Li 3278 (HITBC0002828) • Chengzi Village, 21°52′N, 101°15′E, 20 September 1972, fruiting, G.D.Tao 7035 (HITBC037229, HITBC0002836) • Mengxing Village, ravine rainforest over limestone, 21°50′12.45″N, 101°24′45.25″E, 609 m a.s.l., 16 May 2021, flowering, J.W.Li 6810 (HITBC0075681) • Xishuangbanna Tropical Botanical Garden: 21°54′52.69″N, 101°15′24.30″E, 576 m a.s.l., 24 January 2024, fruiting, D.L.Quan & Y.H.Tan TYH3271 (HITBC0121858) • 21°55′11.86″N, 101°16′4.19″E, 553 m a.s.l., 28 April 2024, flowering, D.L.Quan & Y.H.Tan TYH3280 (HITBC0121859) • 21°55′10.63″N, 101°16′7.14″E, 561 m a.s.l., 30 November 2024, fruiting, D.L.Quan TYH3299 (HITBC0121860) • 21°54′N, 101°15′E, 540 m a.s.l., 2 January 1959, fruiting, Y.H.Li 213 (HITBC0002841) • 21°54′N, 101°15′E, 570 m a.s.l., 13 July 1959, fruiting, Y.H.Li 1602 (HITBC0002840) • 21°41′N, 101°25′E, 570 m a.s.l., 27 April 2011, flowering, J.X.Hu C420152 (HITBC0022962) • 21°41′N, 101°25′E, 570 m a.s.l., 28 April 2011, fruiting, J.X.Hu C420196 (HITBC0022961) • 21°41′N, 101°25′E, 570 m a.s.l., 24 June 2009, fruiting, W.Q.Xiao C400518 (HITBC0022957, HITBC0034323, HITBC0034324) • Guanlei Township, near border, 21°35′5.88″N, 101°8′51.30″E, 596 m a.s.l., 29 April 2021, flowering, J.W.Li 6636 (HITBC0075499) • on the way from Mengyuan Village to Longlin Village, 21°38′37.89″N, 101°26′39.59″E, 1005 m a.s.l., 16 April 2021, flowering, S.K.Peng P1341 (HITBC0076112) • Shangyong Township, Longmen Village, 21°18′4.84″N, 101°33′16.03″E, 971 m a.s.l., 10 May 2019, flowering, S.S.Zhou, J.H.Li & L.X.Wang G4-285 (HITBCXSBN002713) • Yiwu Township, beside the river, 1000 m a.s.l., 9 December 2010, fruiting, J.T.Yin 1879 (HITBC0030358) • Jinghong: Nabanhe, beside the river, 22°10′N, 100°40′E, 700 m a.s.l., 7 November 1988, fruiting, G.D.Tao 44896 (HITBC0002835) • Dadugang Township, Dahuangba Village, tropical lowland rainforest, 22°17′24.64″N, 100°51′34.40″E, 995 m a.s.l., 7 July 2019, flowering, J.W.Li 4857 (HITBC0035650) • Menglong Township: Manyangguang Village, 21°39′N, 100°43′E, 16 October 1978, fruiting, G.D.Tao 19704 (HITBC037215, HITBC037221, HITBC037231, HITBC0002838) • Guanglong Mountain, 21°39′N, 100°43′E, 800 m a.s.l., 6 November 1958, fruiting, S.W.Zhao 0169 (HITBC0002814) • Mengyang Township, in the Mengyang sub-reserve of Xishuangbanna National Nature Reserve, 22°17′25″N, 100°51′35″E, 992 m a.s.l., 7 July 2019, flowering, J.W.Li & L.Wang G3-788 (HITBC-XSBN001702) • Puwen, Dakaihe, 22°35′N, 101°2′E, 840 m a.s.l., 14 August 1977, fruiting, G.D.Tao 16937 (HITBC0002837) • Lincang: Cangyuan County: Banhong Township, humid ravine rainforest, 23°18′N, 99°6′E, 750 m a.s.l., 19 May 1974, flowering, Y.H.Li 11528 (HITBC0002827) • Pu ‘er City: Jiangcheng County: ravine rainforest beside the Tuka River, 4 August 2011, fruiting, S.S.Zhou 10482 (HITBC0002897) • Simao District: Nanping Township, Taiyanghe Provincial Nature Reserve, forest by the ravine, 1 September 2011, fruiting, S.S.Zhou 10760 (HITBC0002787) Ximeng County: Zhongke Township, near river, 22°49′28.47″N, 99°36′58.35″E, 854 m a.s.l., 26 June 2020, flowering, D.P.Ye 1067 (HITBC0062765, HITBC0062766). Laos • Oudomxay Province: Beng Township: Nahom Village, sandy soil wet valley rain forest, 880 m a.s.l., 3 November 1996, flowering, H.Wang 2392 (HITBC0002829).
Supplementary Material
XML Treatment for Eumachia brevipedunculata
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Andersson L (2001) Margaritopsis (Rubiaceae, Psychotrieae) is a pantropical genus.Systematics and Geography of Plants 71(1): 73–85. 10.2307/3668754 · doi ↗
- 2Andersson L (2002) Relationships and generic circumscriptions in the Psychotria complex (Rubiaceae, Psychotrieae).Systematics and Geography of Plants 72: 167–202.
- 3Applequist WL (2014) Report of the nomenclature committee for vascular plants: 66.Taxon 63(6): 1358–1371. 10.12705/636.20 · doi ↗
- 4BarrabéL Davis AP (2013) (2207) Proposal to conserve the name Margaritopsis against Eumachia (Rubiaceae).Taxon 62(5): 1069–1070. 10.12705/625.36 · doi ↗
- 5BarrabéL Buerki S Mouly A Davis AP Munzinger J Maggia L (2012) Delimitation of the genus Margaritopsis (Rubiaceae) in the Asian, Australasian and Pacific region, based on molecular phylogenetic inference and morphology.Taxon 61(6): 1251–1268. 10.1002/tax.616007 · doi ↗
- 6BarrabéL Mouly A Florence J (2013) Psychotriae (Rubiaceae) neocaledonicarum specierum nomenclator.Adansonia 35(2): 281–357. 10.5252/a 2013 n 2a 6 · doi ↗
- 7Berger A Valant-Vetschera K Schinnerl J Brecker L (2022) A revised classification of the sister tribes Palicoureeae and Psychotrieae (Rubiaceae) indicates genus-specific alkaloid accumulation.Phytochemistry Reviews 21: 941–986. 10.1007/s 11101-021-09769-x · doi ↗
- 8Bijmoer R Arias Guerrero S Creuwels J (2024) Naturalis Biodiversity Center (NL) – Botany. Naturalis Biodiversity Center. 10.15468/ib 5ypt [accessed 1 March 2025] · doi ↗