Two new species of larval Erythraeidae (Parasitengona) ectoparasites of leafhoppers from Southwestern China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
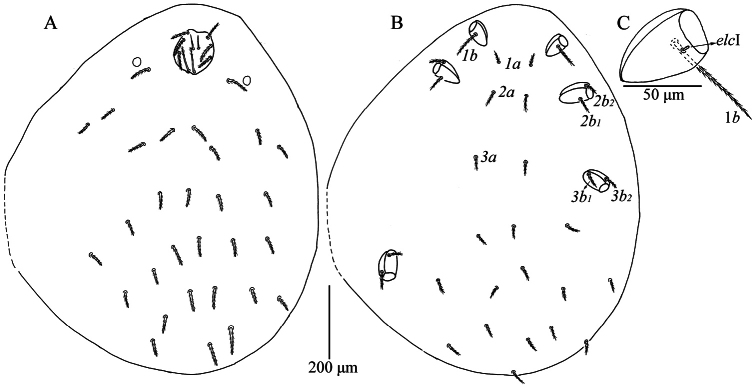 Figure 1
Figure 1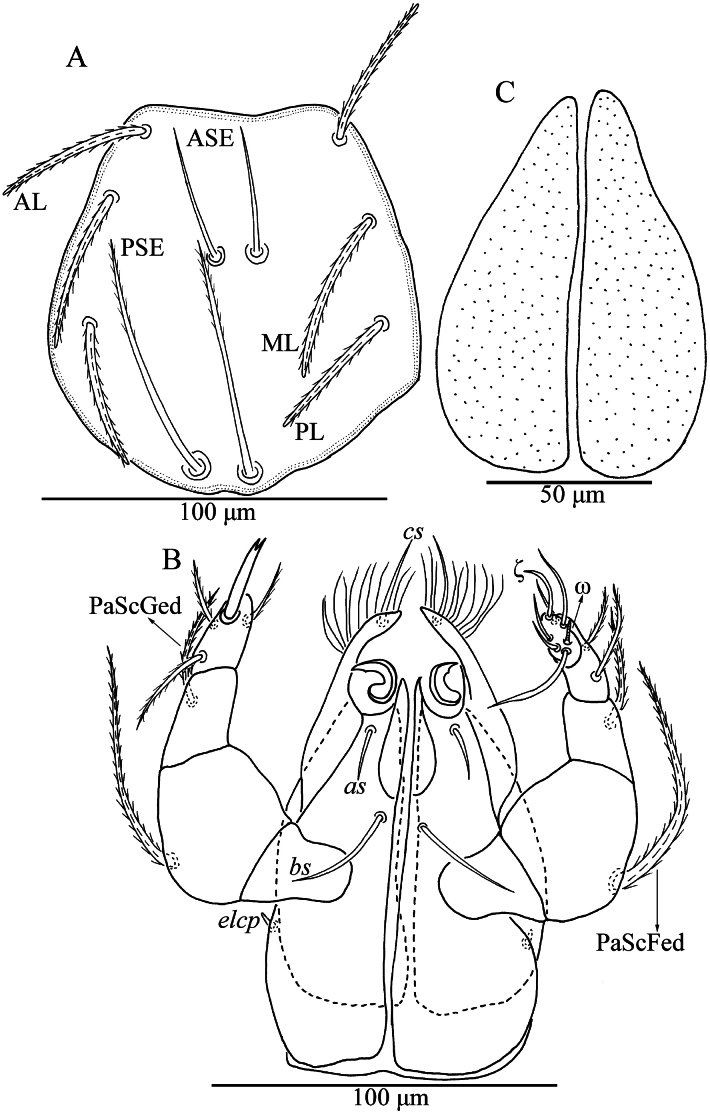 Figure 2
Figure 2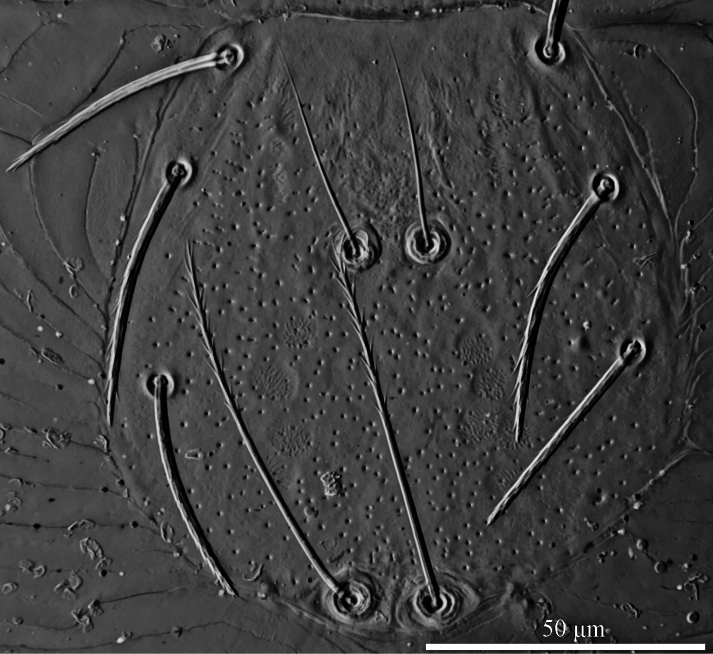 Figure 3
Figure 3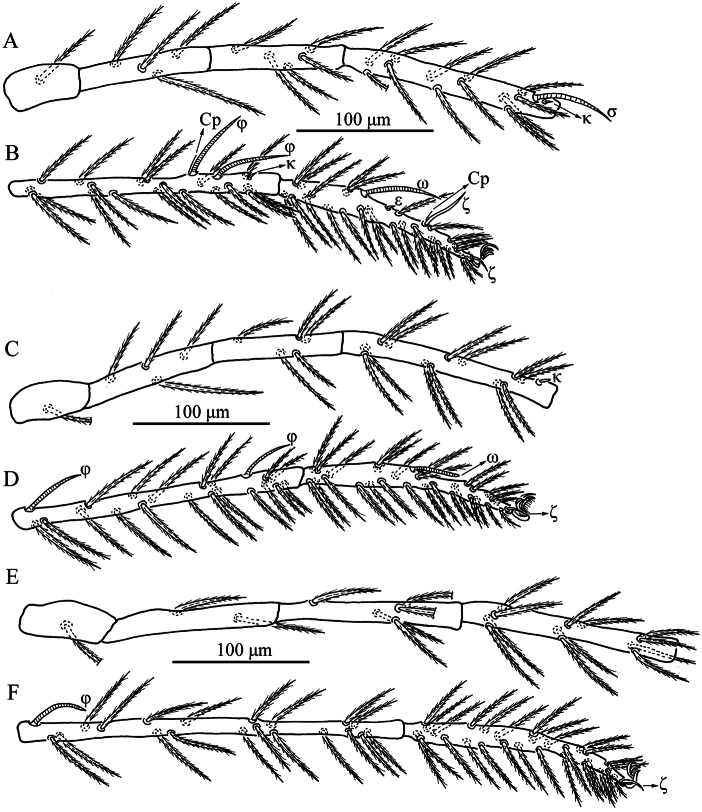 Figure 4
Figure 4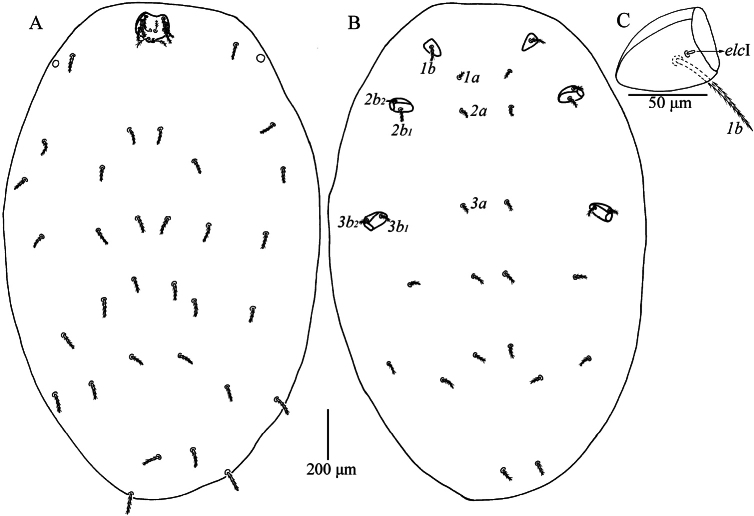 Figure 5
Figure 5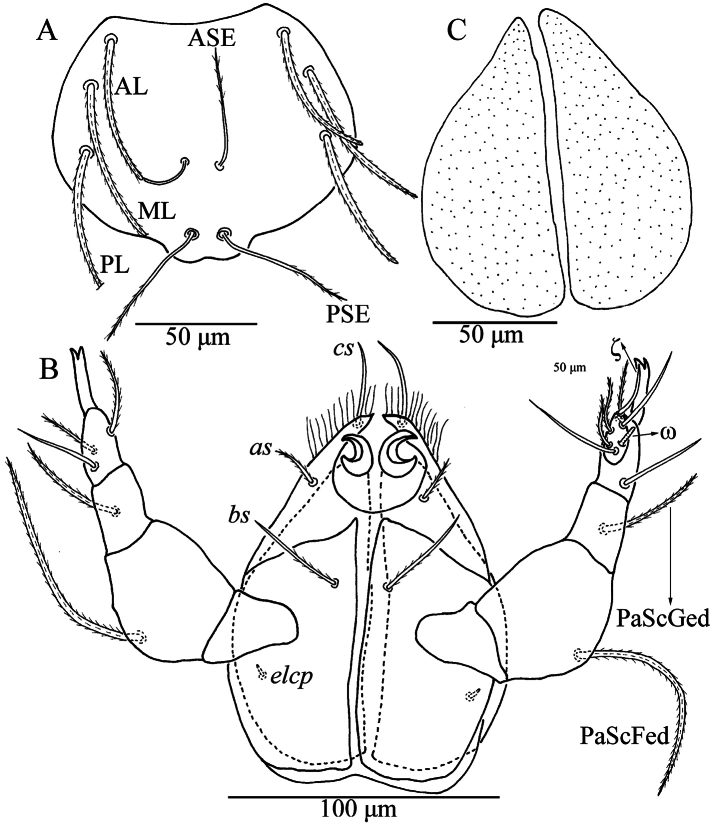 Figure 6
Figure 6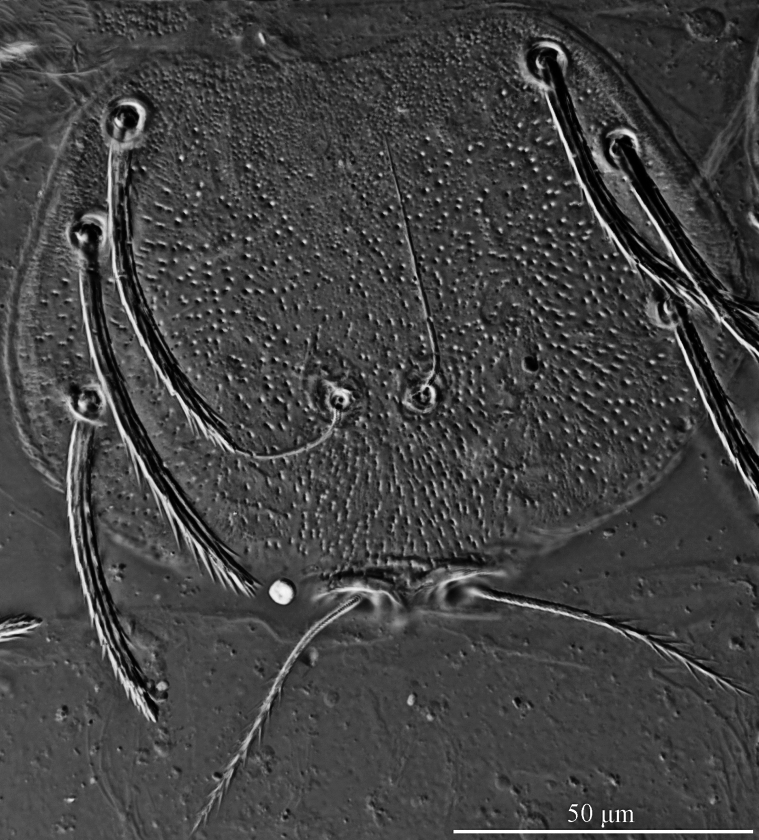 Figure 7
Figure 7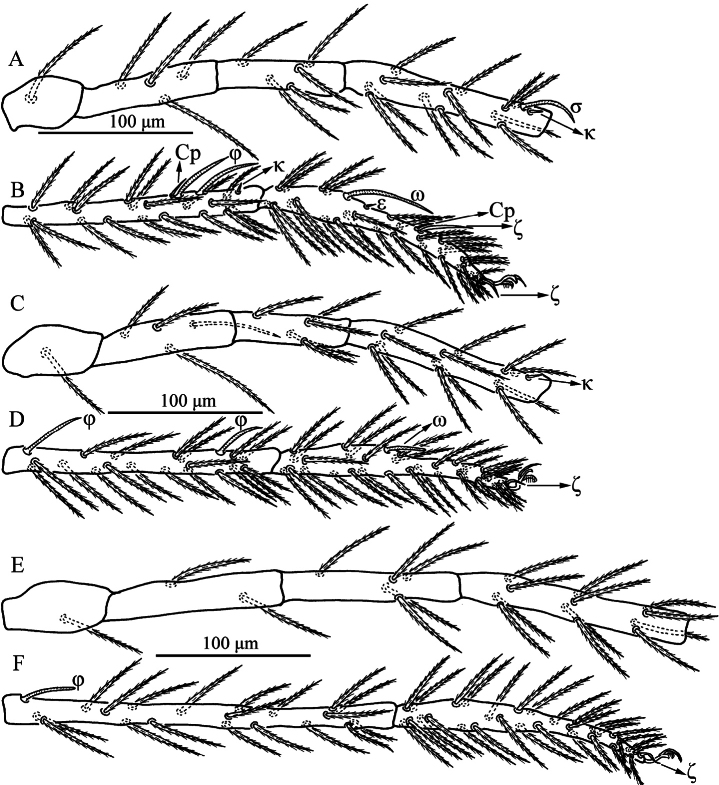 Figure 8
Figure 8| Species | Host | Distribution |
|---|---|---|
| Unknown | Yunnan | |
| Unknown | Xizang | |
| Chongqing, Guizhou | ||
| Chongqing | ||
| Unidentified | Yunnan | |
| Unknown | Guizhou | |
| Unknown. | Yunnan | |
| Unidentified moth ( | Xizang | |
| Unidentified | Yunnan | |
| Unidentified | Guizhou | |
| Guizhou | ||
| Unknown | Guizhou | |
| Unknown | Guizhou |
| Character | Holotype | a | b | c | d | e | f |
| range |
|---|---|---|---|---|---|---|---|---|---|
| fD | 30 | 28 | 30 | 30 | 28 | 30 | 28 | 0.99 | 28–30 |
| fV | 14 | 14 | 12 | 14 | 12 | 14 | 14 | 0.90 | 12–14 |
| NDV | 44 | 42 | 42 | 44 | 40 | 44 | 42 | 1.40 | 40–44 |
| IL | 999 | 689 | 1121 | 901 | 1069 | 1061 | 1543 | 240.10 | 689–1543 |
| IW | 843 | 415 | 840 | 608 | 720 | 678 | 1028 | 181.47 | 415–1028 |
| DS | 30–66 | 33–57 | 33–59 | 32–62 | 36–64 | 33–60 | 33–55 | 1.64–3.58 | 30–66 |
| DS min. | 30 | 33 | 33 | 32 | 36 | 33 | 33 | 1.64 | 30–36 |
| DS max. | 66 | 57 | 59 | 62 | 64 | 60 | 55 | 3.58 | 55–66 |
| PDS | 34–66 | 36–57 | 37–59 | 34–62 | 37–64 | 36–60 | 33–55 | 1.48–3.58 | 33–66 |
| PDS min. | 34 | 36 | 37 | 34 | 37 | 36 | 33 | 1.48 | 33–37 |
| PDS max. | 66 | 57 | 59 | 62 | 64 | 60 | 55 | 3.58 | 55–66 |
| Oc | 21 | 22 | 22 | 23 | 21 | 20 | 19 | 1.25 | 19–23 |
|
| 35 | 30 | 32 | 27 | 21 | 25 | 24 | 4.53 | 21–35 |
|
| 44 | 47 | 43 | 38 | 39 | 44 | 40 | 3.00 | 38–47 |
|
| 38 | 36 | 38 | 32 | 34 | 35 | 33 | 2.17 | 32–38 |
|
| 74 | 67 | 74 | 66 | 72 | 64 | 71 | 3.73 | 64–74 |
|
| 47 | 49 | 43 | 44 | 47 | 48 | 40 | 2.97 | 40–49 |
|
| 36 | 34 | 29 | 32 | 33 | 34 | 31 | 2.12 | 29–36 |
|
| 48 | 44 | 46 | 44 | 46 | 46 | 40 | 2.36 | 40–48 |
|
| 35 | 37 | 36 | 36 | 34 | 34 | 32 | 1.55 | 32–37 |
| L | 114 | 109 | 109 | 110 | 111 | 114 | 103 | 3.46 | 103–114 |
| W | 108 | 103 | 103 | 100 | 103 | 102 | 97 | 3.10 | 97–108 |
| AW | 57 | 60 | 57 | 53 | 55 | 61 | 56 | 2.56 | 53–61 |
| MW | 76 | 71 | 71 | 71 | 74 | 72 | 69 | 2.14 | 69–76 |
| PW | 85 | 78 | 79 | 77 | 79 | 80 | 76 | 2.70 | 76–85 |
| MA | 41 | 38 | 40 | 43 | 40 | 43 | 38 | 1.92 | 38–43 |
| AA | 13 | 12 | 12 | 13 | 12 | 13 | 11 | 0.70 | 11–13 |
| SB | 15 | 14 | 15 | 14 | 15 | 16 | 14 | 0.70 | 14–16 |
| ISD | 63 | 60 | 60 | 57 | 59 | 58 | 56 | 2.14 | 56–63 |
| AP | 56 | 52 | 54 | 55 | 54 | 53 | 54 | 1.20 | 52–56 |
| AL | 46 | 45 | 37 | 44 | 44 | 45 | 40 | 3.02 | 37–46 |
| ML | 50 | 50 | 49 | 48 | 46 | 51 | 47 | 1.67 | 46–51 |
| PL | 41 | 41 | 37 | 37 | 37 | 36 | 35 | 2.19 | 35–41 |
| ASE | 39 | 34 | 33 | 39 | 31 | 36 | 33 | 2.88 | 31–39 |
| PSE | 71 | 66 | 60 | 70 | 60 | 67 | 57 | 5.04 | 57–71 |
|
| 15 | 14 | 16 | 15 | 17 | 18 | 16 | 1.25 | 14–18 |
|
| 34 | 27 | 30 | 36 | 35 | 33 | 34 | 2.91 | 27–36 |
|
| 26 | 23 | 20 | 28 | 25 | 22 | 25 | 2.47 | 20–28 |
| PaScFed | 74 | 78 | 76 | 81 | 83 | 85 | 73 | 4.24 | 73–85 |
| PaScGed | 36 | 33 | 31 | 34 | 37 | 36 | 34 | 1.92 | 31–37 |
| GL | 136 | 141 | 132 | 135 | 136 | 140 | 138 | 2.85 | 132–141 |
| Ta I (H) | 22 | 19 | 22 | 20 | 18 | 18 | 15 | 2.29 | 15–22 |
| Ta I (L) | 152 | 153 | 160 | 159 | 152 | 160 | 157 | 3.44 | 152–160 |
| Ti I | 199 | 191 | 200 | 192 | 201 | 207 | 188 | 6.22 | 188–207 |
| Ge I | 162 | 155 | 166 | 151 | 151 | 162 | 152 | 5.76 | 151–166 |
| TFe I | 93 | 88 | 93 | 88 | 92 | 96 | 89 | 2.81 | 88–96 |
| BFe I | 105 | 96 | 100 | 94 | 97 | 101 | 95 | 3.61 | 94–105 |
| Tr I | 54 | 47 | 48 | 48 | 47 | 44 | 52 | 3.11 | 44–54 |
| Cx I | 56 | 51 | 57 | 58 | 63 | 64 | 62 | 4.27 | 51–64 |
| Ta II (H) | 18 | 20 | 19 | 21 | 22 | 17 | 17 | 1.81 | 17–22 |
| Ta II (L) | 157 | 153 | 158 | 154 | 157 | 159 | 161 | 2.56 | 153–161 |
| Ti II | 215 | 198 | 214 | 204 | 211 | 216 | 194 | 8.17 | 194–216 |
| Ge II | 161 | 150 | 161 | 154 | 157 | 163 | 148 | 5.39 | 148–163 |
| TFe II | 95 | 86 | 85 | 85 | 91 | 90 | 84 | 3.78 | 84–95 |
| BFe II | 95 | 93 | 96 | 96 | 96 | 102 | 91 | 3.16 | 91–102 |
| Tr II | 58 | 54 | 50 | 53 | 48 | 51 | 54 | 3.02 | 48–58 |
| Cx II | 61 | 66 | 69 | 67 | 68 | 74 | 75 | 4.44 | 61–75 |
| Ta III (H) | 15 | 19 | 14 | 13 | 14 | 18 | 14 | 2.12 | 13–19 |
| Ta III (L) | 170 | 167 | 170 | 165 | 171 | 169 | 176 | 3.19 | 165–176 |
| Ti III | 287 | 266 | 277 | 264 | 271 | 277 | 273 | 7.17 | 264–287 |
| Ge III | 161 | 153 | 164 | 153 | 156 | 166 | 156 | 4.87 | 153–166 |
| TFe III | 136 | 124 | 132 | 133 | 128 | 131 | 126 | 3.89 | 124–136 |
| BFe III | 124 | 119 | 121 | 120 | 119 | 128 | 118 | 3.28 | 118–128 |
| Tr III | 66 | 57 | 57 | 53 | 53 | 54 | 51 | 4.61 | 51–66 |
| Cx III | 66 | 66 | 67 | 71 | 72 | 78 | 66 | 4.20 | 66–78 |
| Leg I | 821 | 781 | 824 | 790 | 803 | 834 | 795 | 18.28 | 781–834 |
| Leg II | 842 | 800 | 833 | 813 | 828 | 855 | 807 | 18.34 | 800–855 |
| Leg III | 1010 | 952 | 988 | 959 | 970 | 1003 | 966 | 20.68 | 952–1010 |
| IP | 2673 | 2533 | 2645 | 2562 | 2601 | 2692 | 2568 | 56.05 | 2533–2692 |
| Character | Holotype | a | b | c |
| range |
|---|---|---|---|---|---|---|
| fD | 32 | 34 | 32 | 32 | 0.87 | 32–34 |
| fV | 12 | 12 | 12 | 12 | 0.00 | 12–12 |
| NDV | 44 | 46 | 44 | 44 | 0.87 | 44–46 |
| IL | 1928 | 1998 | 1672 | 1733 | 134.32 | 1672–1998 |
| IW | 1213 | 1320 | 1366 | 1276 | 56.45 | 1213–1366 |
| DS | 46–77 | 42–70 | 47–78 | 48–77 | 2.28–3.20 | 42–78 |
| DS min. | 46 | 42 | 47 | 48 | 2.28 | 42–48 |
| DS max. | 77 | 70 | 78 | 77 | 3.20 | 70–78 |
| PDS | 54–77 | 53–70 | 51–78 | 52–77 | 1.12–3.20 | 51–78 |
| PDS min. | 54 | 53 | 51 | 52 | 1.12 | 51–54 |
| PDS max. | 77 | 70 | 78 | 77 | 3.20 | 70–78 |
| Oc | 20 | 20 | 21 | 23 | 1.22 | 20–23 |
|
| 24 | 28 | 25 | 26 | 1.48 | 24–28 |
|
| 32 | 37 | 34 | 37 | 2.12 | 32–37 |
|
| 33 | 31 | 34 | 34 | 1.22 | 31–34 |
|
| 54 | 63 | 55 | 65 | 4.82 | 54–65 |
|
| 42 | 40 | 41 | 44 | 1.48 | 40–44 |
|
| 40 | 39 | 40 | 39 | 0.50 | 39–40 |
|
| 40 | 39 | 40 | 42 | 1.09 | 39–42 |
|
| 34 | 34 | 37 | 36 | 1.30 | 34–37 |
| L | 103 | 100 | 109 | 106 | 3.35 | 100–109 |
| W | 124 | 123 | 137 | 130 | 5.59 | 123–137 |
| AW | 72 | 70 | 81 | 77 | 4.30 | 70–81 |
| MW | 91 | 88 | 98 | 97 | 4.15 | 88–98 |
| PW | 99 | 98 | 109 | 104 | 4.39 | 98–109 |
| MA | 60 | 59 | 63 | 62 | 1.58 | 59–63 |
| AA | 14 | 15 | 16 | 16 | 0.83 | 14–16 |
| SB | 14 | 14 | 18 | 17 | 1.79 | 14–18 |
| ISD | 30 | 34 | 42 | 37 | 4.38 | 30–42 |
| AP | 47 | 47 | 56 | 60 | 5.68 | 47–60 |
| AL | 56 | 53 | 61 | 59 | 3.03 | 53–61 |
| ML | 70 | 68 | 76 | 71 | 2.95 | 68–76 |
| PL | 59 | 57 | 64 | 62 | 2.69 | 57–64 |
| ASE | 44 | 46 | 46 | 45 | 0.83 | 44–46 |
| PSE | 55 | 59 | 66 | 61 | 3.96 | 55–66 |
|
| 18 | 16 | 15 | 20 | 1.92 | 15–20 |
|
| 37 | 35 | 37 | 41 | 2.18 | 35–41 |
|
| 32 | 34 | 31 | 35 | 1.58 | 31–35 |
| PaScFed | 81 | 76 | 77 | 80 | 2.06 | 76–81 |
| PaScGed | 41 | 37 | 46 | 40 | 3.24 | 37–46 |
| GL | 136 | 146 | 137 | 141 | 3.94 | 136–146 |
| Ta I (H) | 18 | 19 | 19 | 19 | 0.43 | 18–19 |
| Ta I (L) | 159 | 154 | 147 | 157 | 4.55 | 147–159 |
| Ti I | 167 | 166 | 163 | 166 | 1.50 | 163–167 |
| Ge I | 135 | 133 | 131 | 135 | 1.66 | 131–135 |
| TFe I | 83 | 82 | 78 | 78 | 2.28 | 78–83 |
| BFe I | 89 | 90 | 89 | 91 | 0.83 | 89–91 |
| Tr I | 50 | 51 | 56 | 51 | 2.35 | 50–56 |
| Cx I | 73 | 66 | 67 | 63 | 3.63 | 63–73 |
| Ta II (H) | 19 | 16 | 18 | 18 | 1.09 | 16–19 |
| Ta II (L) | 150 | 147 | 143 | 152 | 3.39 | 143–152 |
| Ti II | 178 | 169 | 165 | 174 | 4.92 | 165–178 |
| Ge II | 132 | 130 | 127 | 132 | 2.05 | 127–132 |
| TFe II | 74 | 74 | 72 | 79 | 2.59 | 72–79 |
| BFe II | 91 | 86 | 88 | 89 | 1.80 | 86–91 |
| Tr II | 60 | 56 | 57 | 57 | 1.50 | 56–60 |
| Cx II | 81 | 86 | 78 | 79 | 3.08 | 78–86 |
| Ta III (H) | 17 | 16 | 17 | 19 | 1.09 | 16–19 |
| Ta III (L) | 165 | 164 | 154 | 163 | 4.39 | 154–165 |
| Ti III | 252 | 251 | 246 | 254 | 2.95 | 246–254 |
| Ge III | 150 | 144 | 139 | 147 | 4.06 | 139–150 |
| TFe III | 115 | 117 | 118 | 111 | 2.68 | 111–118 |
| BFe III | 117 | 120 | 121 | 114 | 2.74 | 114–121 |
| Tr III | 67 | 61 | 56 | 54 | 5.02 | 54–67 |
| Cx III | 80 | 86 | 86 | 77 | 3.90 | 77–86 |
| Leg I | 756 | 742 | 731 | 741 | 8.90 | 731–756 |
| Leg II | 766 | 748 | 730 | 762 | 14.10 | 730–766 |
| Leg III | 946 | 943 | 920 | 920 | 12.30 | 920–946 |
| IP | 2468 | 2433 | 2381 | 2423 | 31.01 | 2381–2468 |
- —National Natural Science Foundation of China 501100001809 https://ror.org/01h0zpd94 http://doi.org/10.13039/501100001809
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsStudy of Mite Species · Insect-Plant Interactions and Control · Parasite Biology and Host Interactions
Introduction
Southwestern China comprises three provinces (Sichuan, Guizhou, and Yunnan), one municipality (Chongqing), and one autonomous region (Xizang (Tibet)). This region is characterized by diverse topography with significant variations in elevation and numerous separated basins (Shi et al. 2018), including the Yunnan-Guizhou Plateau, the Qinghai-Tibetan Plateau, the Hengduan Mountains, and the Sichuan Basin. Simultaneously, the region exhibits diverse climatic conditions, including a subtropical monsoon climate, plateau mountain climate, mountain climate, tropical monsoon climate, dry-hot valley climate and a temperate monsoon climate. The variety of climates provides a wealth of environmental conditions, which play a pivotal role in fostering species diversity in the ecosystem. Therefore, Southwestern China is a key area for fauna-flora biodiversity research in China (Han et al. 2008; Shi et al. 2018).
To date, 13 species in eight genera (Balaustium von Heyden, 1826, Caeculisoma Berlese, 1888, Charletonia Oudemans, 1910, Erythraeus Latreille, 1806, Grandjeanella Southcott, 1961, Leptus Latreille, 1796, Marantelophus Haitlinger, 2011, and Neoabrolophus Khot, 1965) of five subfamilies (Abrolophinae Witte, 1995, Balaustiinae Grandjean, 1947, Callidosomatinae Southcott, 1957, Erythraeinae Robineau-Desovidy, 1828, and Leptinae Billberg, 1820) of Erythraeidae have been documented in Southwestern China (Xu et al. 2017, 2019a, b, 2020, 2022a, 2022b, 2023, 2025). Of them, three species were described based on post-larval forms, one species was recorded based on both larval and post-larval stages, and the remaining species were known only by larvae (Table 1).
Species of the genus Abrolophus Berlese, 1891 from China are distributed in the Macao Special Administrative Region, Hainan Province, Zhejiang Province, Shandong Province, Guangxi Zhuang Autonomous Region, Guangdong Province, and Hunan Province (Zheng 2002a; Haitlinger 2006; Xu et al. 2021, 2022a). However, there has been no report of Abrolophus in Southwestern China.
A total of 27 species of Caeculisoma have been reported worldwide, of which 14 were based on the post-larval stage, 12 were based on the larval stage, and only one species was based on both larval and post-larval instars (Mąkol and Wohltmann 2012; Xu et al. 2019a, 2019b, 2020; Saboori et al. 2023; Kohansal et al. 2024; Noei et al. 2024). Among the 27 known Caeculisoma species, four were recorded in China based on the larval stage, with two (C.penlineatus Xu & Jin, 2019, C.semispinus Xu & Jin, 2019) collected from Southwestern China (Zheng 2002b; Xu et al. 2019a, 2019b, 2020).
Hitherto, only two species of the genus Iguatonia have been reported based on their larval stage worldwide (Haitlinger 2004; Xu et al. 2020; Noei et al. 2024): I.barbillae Haitlinger, 2004 from Brazil and I.xinfengi Xu & Jin, 2020 from Hainan Province (Island), China.
In this study, two new species, C.taianensis sp. nov. and I.barboproxima sp. nov., collected from Yunnan Province and Chongqing Municipality, respectively, are described and illustrated based on larvae. Additionally, new data for A.quadrapexicis Xu & Jin, 2022 is provided.
Material and methods
Erythraeid larvae were collected along with their insect hosts using 200 mesh insect nets, and subsequently preserved in small vials containing absolute ethanol. The larvae on the hosts were detached using a fine brush under a stereomicroscope (Nikon SMZ745) in the lab. Then, all larval specimens were cleared in Oudemans’ fluid for about 12 h at 25 °C and slide-mounted in Hoyer’s medium (Walter and Krantz 2009). Figures were drawn with the aid of a drawing tube attached to a Nikon Eclipse Ni-E compound microscope. Genus identification is based on the key to world genera of larval Callidosomatinae of Noei et al. (2024). Terminology and abbreviations are adapted from Noei et al. (2024), Wohltmann et al. (2007) and Xu et al. (2020). Measurements are expressed in micrometers (μm). The standard deviations (SD) are provided with two decimal places. All specimens were deposited at the Institute of Entomology, Guizhou University, Guiyang, China (GUGC).
Results
Abrolophinae Witte, 1995
Abrolophus Berlese, 1891
Abrolophus
quadrapexicis
Taxon classificationAnimaliaTrombidiformesErythraeidae
Xu & Jin, 2022
A26E19A0-AF53-5C5F-961E-59122D327E44
Material examined.
China • one larva (2000–1600-GZ-yj); Guizhou Province, Fanjingshan National Nature Reserve; 27°59'27″N, 108°33'30″E; 673 m; 30 Jun. 2023; Si-Yuan Xu leg.; on an unidentified Psyllidae (Hemiptera).
Distribution.
Guizhou Province (new distribution), Shandong Province, Zhejiang Province.
Note.
This species was collected from plants without a host record in Zhejiang and Shandong (Xu et al. 2022b).
Callidosomatinae Southcott, 1957
Caeculisoma Berlese, 1888
Caeculisoma
taianensis
sp. nov.
Taxon classificationAnimaliaTrombidiformesErythraeidae
F0A460C9-EA62-55AC-BE81-E3CE2C1A1F06
https://zoobank.org/F5C1A34A-2366-423A-B816-969D2202B854
Diagnosis (larva).
ASE nude and posterior to the level of ML, closer to ML than PL; PSE with barbs on the distal one-third; gnathosoma with two pairs of nude hypostomalae; ISD 56–63; Ti I 188–207; Ti III 264–287.
Description.
Dorsum. Idiosoma lateral cuticle of holotype used for drawing broken slide preparation, almost oval, with 30 (fD = 28–30 in paratypes) barbed setae, a pair of setae located between scutum and eyes (Fig. 1A). Scutum outline pentagonal with rounded angles, length somewhat longer than width, anterior margin slightly concave, anterolateral and posterolateral margins slightly sinuous, posterior margin with small concavity between bases of PSE (Figs 1A, 2A, 3). Three pairs of normal setae (AL, ML and PL), and two pairs of sensilla (ASE and PSE) placed on scutum. AL, ML and PL completely barbed, ASE nude, PSE with fine barbs in distal about one-third. ASE placed between ML and PL, and closer to ML than PL, PSE near posterior margin of scutum. PSE much longer than ASE, ML slightly longer than AL and PL, AL slightly longer than PL, one paratype (c) AL equal to PL (Table 2).
Caeculisomataianensis sp. nov., larva A dorsal view of idiosoma B ventral view of idiosoma C dorsal view of coxa I, showing supracoxal seta.
Caeculisomataianensis sp. nov., larva A scutum B ventral view of gnathosoma C dorsal view of the cheliceral bases.
Caeculisomataianensis sp. nov., larva photograph. Scutum, showing the shape of scutum, ASE and PSE.
Venter. All ventral setae, including coxalae, barbed and with pointed ends (Fig. 1B). Three pairs of intercoxal setae (1a, 2a and 3a), 2a longer than 1a and 3a, 3a slightly longer than 1a (Table 2), 14 setae behind coxae III (fV = 12–14 in paratypes). Five pairs of coxalae (1b, 2b_1_, 2b_2_, 3b_1_ and 3b_2_), 1b much longer than the other coxalae, 2b_1_ subequal 3b_1_, 2b_2_ and 3b_2_ subequal, 2b_1_ and 3b_1_ longer than 2b_2_ and 3b_2_, respectively (Table 2). Dorsum of coxa I with a peg-like supracoxal seta (elc I) (Fig. 1C).
Gnathosoma (Fig. 2B, C). Dorsal view of the cheliceral base punctated. One pair of galealae (cs) and two pairs of hypostomalae (as and bs) nude; bs longer than cs and much longer than as (Table 2). Hypostomal lip with fimbriation. Palpfemur and palpgenu each with one barbed, pointed dorsal seta (PaScFed and PaScGed). Palptibia with three barbed setae, one on ventral surface, odontus bifid. Palptarsus with seven setae, five nude, one solenidion (ω) and one eupathidium (ζ). fPp = 0-B-B-3B_2_-5Nωζ. Palpal supracoxal seta (elcp) peg-like.
Legs (Figs 1B, 3). With seven segments (femora divided). IP = 2533–2692 (Holotype and six paratypes). Claws hook-like and posterior claw with few ciliations, and empodium claw-like. Normal setae on legs barbed and pointed. Leg setal formula: leg I: Cx—1n; Tr—1n; Bfe—4n; Tfe—5n; Ge—1σ, 1κ, 12n; Ti—2φ, 1κ, 1Cp, 18n; Ta—1ω, 1ε, 2ζ, 1Cp, 27n. leg II: Cx—2n; Tr—1n; Bfe—4n; Tfe—5n; Ge—1κ, 12n; Ti—2φ, 19n; Ta—1ω, 1ζ, 28n. leg III: Cx—2n; Tr—1n; Bfe—2n; Tfe—5n; Ge—12n; Ti—1φ, 19n; Ta—1ζ, 28n. The morphometric data of the legs is listed in Table 2.
Etymology.
The new species’ name is derived from Taian Town, where the holotype and paratype were collected.
Material examined.
Holotype. China • a larva (1087–0275-YN-yl); Yunnan Province, Yulong County, Taian Town; 26°37'8"N, 100°02'9"E; 2502 m; 8 Aug. 2021; Yan Jiang leg.; from Atkinsoniella sp. (Hemiptera, Cicadellidae). Paratypes China • one larva (1088–0275-YN-yl), same data as the holotype. China • three larvae (1082–0271-YN-yl, 1083–0271-YN-yl, 1084–0271-YN-yl); Yunnan Province, Yulong County, Taian Town; 26°37'8"N, 100°02'9"E; 2502 m; 8 Aug. 2021; Yan Jiang leg.; from unidentified Atkinsoniella (Hemiptera: Cicadellidae). China • one larva (1085–0272-YN-yl); Yunnan Province, Yulong County, Taian Town; 26°37'8"N, 100°02'9"E; 2502 m; 8 Aug. 2021; Yan Jiang leg.; from an unknown nymph of Cicadellidae. China • one larva (1086–0273-YN-yl); Yunnan Province, Yulong County, Taian Town; 26°37'8"N, 100°02'9"E; 2502 m; 8 Aug. 2021; Yan Jiang leg.; from an unidentified nymph of Cicadellidae.
The holotype and paratypes are deposited in the Institute of Entomology, Guizhou University, Guiyang, China (GUGC).
Remarks.
Based on the description of larvae, the genus Caeculisoma includes 13 species so far; of them, four species are from Australia and four from China, two species are from Brazil, while the remaining three species are found in Iran, the Republic of South Africa and New Zealand, respectively (Mąkol and Wohltmann 2012; Xu et al. 2020; Kohansal et al. 2024; Noei et al. 2024).
Similar species to C.taianensis sp. nov. are currently known as C.darwiniense Southcott, 1961, C.mouldsi Southcott, 1988, C.pouyani Noei & Kohansal, 2024 and C.sparnoni Southcott, 1972 based on the key to species of Caeculisoma in Kohansal et al. (2024).
The new species differs from C.darwiniense by the shape of ASE (nude vs barbed), posterior hypostomalae (bs) (nude vs barbed), galealae (cs) (nude vs barbed), the number of normal setae on fn Ge I–III (12-12-12 vs 11-12-13), the number of normal setae on fn Ti I–III (18-19-19 vs 17-18-20), longer L (103–114 vs 87–88), Ti I (188–207 vs 102), and Ti III (264–287 vs 143); differs from C.mouldsi by the shape of ASE (nude vs barbed), shape of posterior hypostomalae (nude vs barbed), gnathosoma with two pairs hypostomalae (vs one pair hypostomalae), the number of normal setae on fn Ge I–III (12-12-12 vs 12-11-12), the number of normal setae on fn Ti I–III (18-19-19 vs 18-19-18), longer 1b (64–74 vs 37–58), PaScFed (73–85 vs 42), leg II (800–855 vs 750), leg III (952–1010 vs 915), IP (2533–2692 vs 2455); differs from C.pouyani by the shape of ASE (nude vs barbed), galealae (nude vs barbed), hypostomalae (nude vs barbed), palp tarsus with five nude normal setae (vs with five barbed normal setae), longer W (97–108 vs 75–87), 1b (64–74 vs 40–46), Ti I (188–207 vs 85–95), Ti II (194–216 vs 85–92), Ti III (264–287 vs 115–130), IP (2533–2692 vs 1360–1505) and differs from C.sparnoni by shape of ASE (nude vs barbed), cheliceral bases without striations (vs with lengthwise striations), longer L (103–114 vs 83), W (97–108 vs 85), 1b (64–74 vs 28), Ta I (150–160 vs 77), Ti I (188–207 vs 83), Ta III (165–176 vs 79), Ti III (264–287 vs 100), IP (2533–2692 vs 1220).
The differences between the new species and the four present species of Caeculisoma found in China are as follows: C.taianensis sp. nov. differs from C.allopenlineatus by the positions of ASE (ASE closer to ML than PL vs ASE closer to PL than ML), shape of scutum (pentagonal vs oval), ASE (nude vs barbed), the longer Ti I (188–207 vs 164–170), Ti II (194–216 vs 151–161), Ti III (264–287 vs 227–239), IP (2533–2692 vs 2061–2115), the shorter ML (46–51 vs 114–120); differs from C.hunanica by the number of solenidia on Ti II (2 vs 1), the number of normal setae on TFe III (5 vs 4), the number of normal setae on fn Ti I–III (18-19-19 vs 16-16-18), longer leg I (781–834 vs 655), leg II (800–855 vs 621), leg III (952–1010 vs 728), IP (2533–2692 vs 2004); differs from C.penlineatus by ASE base location (closer to ML than PL vs in line with the level of PL), BFe I and II with four barbed setae (vs with three barbed setae and one nude seta), palptarsus with one eupathidium (vs with two eupathidia), longer Ti I (188–207 vs 143–167), Ti II (194–216 vs 150–179), Ti III (264–287 vs 213–239), leg I (781–834 vs 627–709), leg II (800–855 vs 645–724), leg III (952–1010 vs 768–878), IP (2533–2692 vs 2060–2298), data based on Xu et al. (2019b, 2020) and differs from C.semispinus by the shape of ASE (nude vs with barbs on distal halves), palptibia with three barbed setae (vs with one barbed seta and two nude setae), palptarsus with five nude setae (vs with two barbed setae and three nude setae), longer L (103–114 vs 78–84), W (97–108 vs 71–80), ISD (56–63 vs 41–44), Ti I (188–207 vs 149–163), Ti III (264–287 vs 213–227), IP (2533–2692 vs 2041–2100).
Caeculisomataianensis sp. nov., larva. Leg I A trochanter—genu B tibia—tarsus; leg II C trochanter—genu D tibia—tarsus; leg III E trochanter—genu F tibia—tarsus.
Iguatonia Haitlinger, 2004
Iguatonia
barboproxima
sp. nov.
Taxon classificationAnimaliaTrombidiformesErythraeidae
BB63528E-EA2F-5BBD-B8AD-E7FFC9B27022
https://zoobank.org/B7AA6D5B-ADD7-4F98-9245-1789A345EFD3
Diagnosis (larva).
ASE and PL located in posterior half of scutum; ASE and PSE with fine barbs on distal halves; two pairs hypostomalae barbed; ISD 30–42.
Description.
Idiosoma almost oval, with 32 (fD = 32–34 in paratypes) barbed setae, a pair of setae located between scutum and eyes at level with PSE bases (Fig. 5A). Scutum about trapezoid outline with rounded angles, wider than long, anterior margin concave, lateral margins arcuate obviously, posterior margin convex in median and with small concave between bases of PSE (Figs 6A, 7). Scutum with three pairs of normal setae (AL, ML and PL) and two pairs of sensilla (ASE and PSE). AL, ML and PL completely barbed, AL slightly shorter than PL, and ML longer than both, PW > MW > AW (Table 3). ASE and PSE with setules in distal half, ASE bases posterior to PL bases, PL placed in posterior half of scutum, PSE near posterior border of scutum and longer than ASE (Fig. 6A, 7).
Iguatoniabarboproxima sp. nov., larva A dorsal view of idiosoma B ventral view of idiosoma C dorsal view of coxa I, showing supracoxal seta.
Iguatoniabarboproxima sp. nov., larva A scutum B ventral view of gnathosoma C dorsal view of the cheliceral bases.
Iguatoniabarboproxima sp. nov., larva photograph. Scutum, showing the shape of scutum, ASE and PSE.
Venter. All ventral setae, including coxalae, barbed and with pointed ends (Fig. 5B). Dorsum of coxa I with a peg-like supracoxal seta (elc I) (Fig. 5C). Three pairs of intercoxal setae (1a, 2a and 3a), 1a posterior to level of posterior edge of coxae I, 2a between coxae II, and 3a at a line with anterior edges of coxae III. 2a and 3a subequal and both slightly longer than 1a (Table 3). Five pairs of coxalae (1b, 2b_1_, 2b_2_, 3b_1_ and 3b_2_), 1b longest, 2b_1_, 3b_1_, and 2b_2_ subequal and all slightly longer than 3b_2_ (Table 3). 12 setae behind coxae III (fV = 12 in paratypes).
Gnathosoma (Fig. 6B) with a pair of nude galealae (cs), two barbed anterior hypostomalae (as) and two posterior hypostomalae with barbs on proximal half, bs slightly longer than cs, and both longer than as (Table 3). Hypostomal lip fimbriated. Cheliceral bases punctate on the dorsal surface (Fig. 6C). Palpfemur and palpgenu, each with one barbed, pointed dorsal seta. Palptibia with one nude ventral seta, one barbed ventral seta, and one barbed dorsal seta, odontus bifid. Palptarsus with seven setae, three barbed, two nude, one solenidion and one eupathidium. fPp = 0-B-B-2BN_2_-3B2Nωζ. Palpal supracoxal seta (elcp) peg-like.
Legs (Figs 5B, 8) with seven segments (femora divided). IP = 2381–2468 (Holotype and three paratypes) (Table 3). Anterior and posterior claws hook-like, subequal in length, and anterior claw with few ciliations. Claw-like empodium falciform, longer and slenderer than lateral claws. Normal setae on legs barbed and pointed. Leg setal formula: Leg I: Cx—1n; Tr—1n; Bfe—4n; Tfe—5n; Ge—1σ, 1κ, 12n; Ti—2φ, 1κ, 1Cp, 18n; Ta—1ω, 1ε, 2ζ, 1Cp, 29n. leg II: Cx—2n; Tr—1n; Bfe—4n; Tfe—5n; Ge—1κ, 12n; Ti—2φ, 19n; Ta—1ω, 1ζ, 30n. leg III: Cx—2n; Tr—1n; Bfe—2n; Tfe—5n; Ge—12n; Ti—1φ, 19n; Ta—1ζ, 30n.
Iguatoniabarboproxima sp. nov., larva. Leg I A trochanter—genu B tibia—tarsus; leg II C trochanter—gen D tibia—tarsus; leg III E trochanter—genu F tibia—tarsus.
Etymology.
The specific epithet of the new species refers to the posterior hypostomalae, which exhibit fine barbs on their proximal half.
Material examined.
Holotype China • a larva (2023–1767-CQ-wx); Chongqing Municipality, Wuxi County, Shuangyang Town; 31°31'29"N, 109°50'12"E; 1151 m; 30 Jun. 2022; Yan Jiang leg.; from an unidentified nymph of Cicadellidae (Hemiptera). Paratypes China • two larvae (2024–1767-CQ-wx, 2025–1767-CQ-wx), the same data as the holotype. China • one larva (2026–1768-CQ-wx); Chongqing Municipality, Wuxi County, Shuangyang Town; 31°29'28"N, 109°49'44"E; 1132 m; 30 Jun. 2022; Xiao-Li Xu leg.; from an unidentified nymph of Cicadellidae (Hemiptera).
The holotype and paratypes are deposited in the Institute of Entomology, Guizhou University, Guiyang, China (GUGC).
Remarks.
To date, two species of Iguatonia have been documented based on larvae, one from Brazil and another one from China (Haitlinger 2004; Noei et al. 2024; Xu et al. 2020).
Iguatoniabarboproxima sp. nov. differs from I.barbillae by the shape of scutum (about trapezoid vs quadrilateral), PL location (in posterior half of scutum vs in anterior half of scutum), ASE location (closer to PL than PSE vs far from PL and near PSE), longer ISD (30–42 vs 10), Ti I (163–167 vs 76–78), and Ti III (246–254 vs 116–126) and differs from I.xinfengi by the shape of scutum (about trapezoid vs sub-rounded), shape of hypostomalae (barbed vs nude), positions of ASE (far from PL and closer to PL than PSE vs almost at the same line with PL), longer ISD (30–42 vs 20–21), Ti I (163–167 vs 114–117), and Ti III (246–254 vs 194–197).
Discussion
Considering previously published data and the present study, 13 species of Caeculisoma have been documented based on the larval stage and one species was recorded based on both the larval and post-larval instars. Of them, only five species (C.brazilensis Noei & Šundić, 2024; C.carmenae Haitlinger, 2008; C.nestori Haitlinger, 2004; C.hunanica Zheng, 2002; and C.sparnoni Southcott, 1972) are without host records (Southcott 1972; Zheng 2002b; Haitlinger 2004, 2008; Kohansal et al. 2024; Noei et al. 2024). The hosts of Caeculisoma larvae were recorded in Insecta, comprising three orders (Lepidoptera, Orthoptera, Hemiptera), and seven families (Acrididae, Cicadellidae, Cicadidae, Delphacidae, Geometridae, Issidae, Miridae) (Southcott 1961, 1972, 1988; Stroiński et al. 2013, Xu et al. 2019a, b, 2020; Kohansal et al. 2024). Only C.pouyani Noei & Kohansal, 2024 has a host distribution across three families (Acrididae, Cicadellidae, Miridae) in two orders (Hemiptera, Orthoptera); and C.penlineatus Xu & Jin, 2019 has a host distribution across three families (Cicadellidae, Delphacidae and Issidae). And the host of each of the remaining species was only recorded in a single family: three species (C.allopenlineatus Xu & Jin, 2020; C.semispinus Xu & Jin, 2019; C.taianensis sp. nov.) with hosts were recorded in Cicadellidae (Hemiptera), two species (C.cooremani Southcott, 1972; C.darwiniense Southcott, 1961) with hosts were recorded in Acrididae (Orthoptera), C.mouldsi Southcott, 1988 with a host was recorded in Cicadidae (Hemiptera), and C.huxleyi Southcott, 1972 with a host was recorded in Geometridae (Lepidoptera). According to the available data, there is a higher probability of finding larvae of the genus Caeculisoma ectoparasitic on Hemiptera compared to Orthoptera and Lepidoptera.
Iguatoniabarbillae Haitlinger, 2004 from an unidentified Homoptera insect (Hemiptera), I.xinfengi Xu & Jin, 2020 from an unknown Delphacidae (Hemipteran) and I.barboproxima sp. nov. from an unidentified Cicadellidae species (Hemiptera) indicate that the host’s spectrum of Iguatonia species is limited to the order Hemiptera.
Only three families of Hemiptera, including Cicadellidae, Delphacidae and Issidae were documented in China for Caeculisoma hosts. A similar situation occurs in the genus Iguatonia, whose hosts are from two families (Cicadellidae, Delphacidae) of Hemiptera.
According to the present information of these two genera the hosts are limited, which may be related to the locations, times, and methods of collection. Therefore, in future studies of their taxonomy, more attention should be paid to geographical ranges and the use of diverse collection methods at different periods in order to know more about the host.
Supplementary Material
XML Treatment for Abrolophus quadrapexicis
XML Treatment for Caeculisoma taianensis
XML Treatment for Iguatonia barboproxima
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Haitlinger R (2004) Charletoniadomawiti n. sp., Caeculisomanestori n. sp. and Iguatoniabarbillae n. gen. and n. sp. from Brazil (Acari: Prostigmata: Erythraeidae).Genus 15(3): 435–444.
- 2Haitlinger R (2006) Eight new species and new records of mites (Acari: Prostigmata: Erythraeidae, Trombidiidae, Johnstonianidae) from China including Macao.Systematic and Applied Acarology 11: 83–105. 10.11158/saa.11.1.11 · doi ↗
- 3Han B Ouyang ZY Xu WH Xiao J Jiang MK Wang Z Qin WH (2008) The effect of protecting species diversity in Southwest China.Acta Ecologica Sinica 28(9): 4589–4593. [In Chinese with English abstract]
- 4Kohansal M Noei J Ramroodi S Rakhshani E (2024) First record of larval Caeculisoma (Acari: Erythraeidae) from Iran with description a new species.Acarologia 64(4): 1182–1190. 10.24349/j 8kk-8r 4k · doi ↗
- 5Mąkol J Wohltmann A (2012) An annotated checklist of terrestrial Parasitengona (Actinotrichida: Prostigmata) of the world, excluding Trombiculidae and Walchiidae.Annales Zoologici 62(3): 359–562. 10.3161/000345412 X 656671 · doi ↗
- 6Noei JŠundićM Bernardi LFO (2024) A new species of larval Caeculisoma (Trombidiformes: Erythraeidae) from Brazil with a key to species.Systematic and Applied Acarology 29(10): 1437–1448. 10.11158/saa.29.10.10 · doi ↗
- 7Saboori A StarýJ Masoumi H Cakmak I (2023) Two new species of Caeculisoma (Trombidiformes: Erythraeidae) from Madagascar.Acarologia 63(4): 1225–1259. 10.24349/vqaq-42d 1 · doi ↗
- 8Shi XW Zhang L Zhang JJ Ouyang ZY Xiao Y (2018) Priority area of biodiversity conservation in Southwest China.Chinese Journal of Ecology 37(12): 3721–3728. [In Chinese with English abstract]