Effect of controlled human Plasmodium falciparum infection on B cell subsets in individuals with different levels of malaria immunity
Pilar Requena, Gloria Patricia Gómez-Pérez, Matthew B. B. McCall, Diana Barrios, Ruth Aguilar, Julia Fernández-Morata, Marta Vidal, Joseph J. Campo, Carla Sanchez, Maria Yazdabankhsh, B. Kim Lee Sim, Stephen L. Hoffman, Peter Kremsner, Bertrand Lell, Benjamin Mordmüller

TL;DR
The study shows how malaria infection affects different types of B cells in people with varying levels of immunity, including those who are vaccinated or naturally exposed.
Contribution
The study reveals how malaria immunity and vaccination influence B cell subset frequencies and how CHMI modulates these effects.
Findings
Semi-immune individuals had higher baseline frequencies of immature and active B cell subsets compared to malaria-naïve individuals.
CHMI caused an expansion of resting classical memory B cells in all groups during early infection.
Vaccination altered the effect of CHMI on B cells, with specific associations observed for resting classical memory B cells and active naïve B cells.
Abstract
Continuous exposure to Plasmodium falciparum (Pf) has been associated with alterations in B cells. We investigated the effect of controlled human malaria infection (CHMI) on B cell phenotypes in individuals with different Pf immunity status: malaria-naïve, immunized with PfSPZ-CVac and semi-immune (lifelong-exposed) volunteers. Compared to naïve, semi-immune but not vaccinated individuals, had increased baseline frequencies of immature B cells (CD19+CD10+), active naive (IgD+CD27−CD21−) B cells, active atypical (IgD−CD27−CD21−) memory B cells (MBCs), active classical (IgD−CD27+CD21−) MBCs and CD1c+-B cells but lower frequencies of some IgG+-B cells. The frequencies of CD1c+ active atypical MBCs correlated positively with anti-Pf antibodies and negatively with circulating eotaxin levels, while the opposite was observed for IgG+ resting atypical MBCs. During early blood-stage infection…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
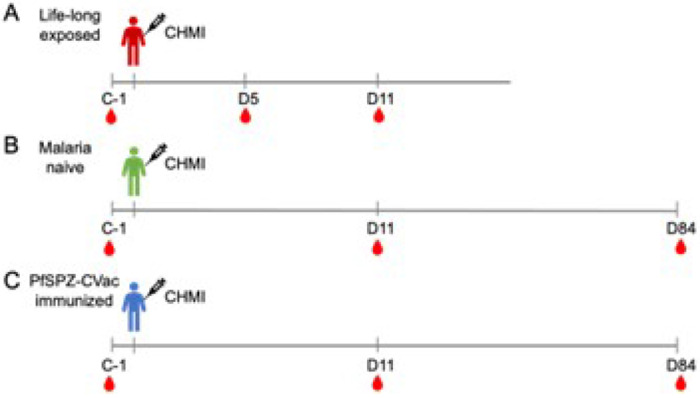 Figure 1
Figure 1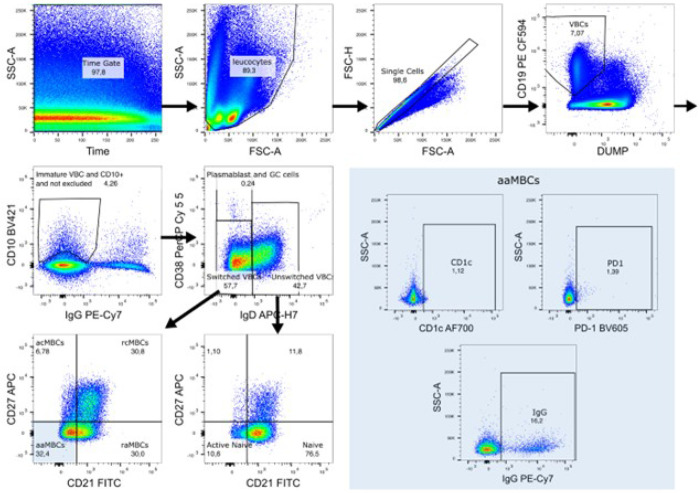 Figure 2
Figure 2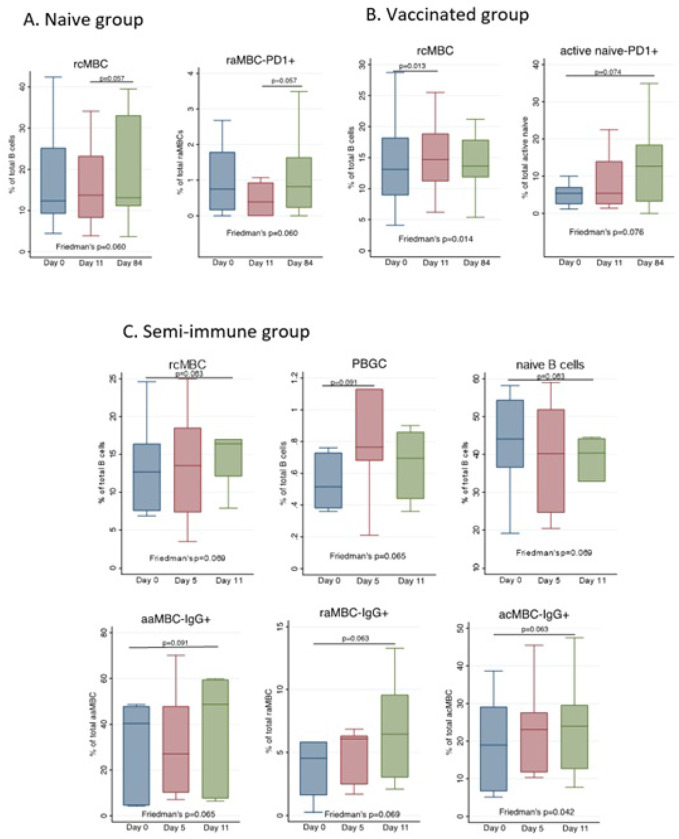 Figure 3
Figure 3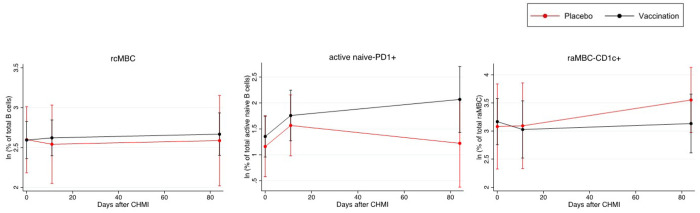 Figure 4
Figure 4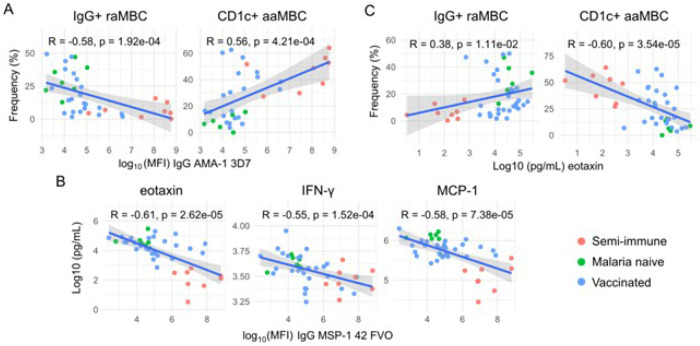 Figure 5
Figure 5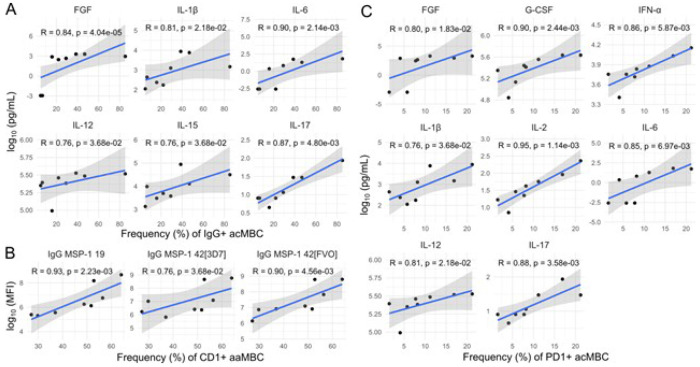 Figure 6
Figure 6Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMalaria Research and Control · Mosquito-borne diseases and control · Viral Infections and Vectors
INTRODUCTION
Malaria is one of the most serious health problems worldwide, with 263 million cases and 597,000 deaths estimated in 2023 [1]. Among Plasmodium species causing human malaria, P. falciparum (Pf) is the major responsible for the high rates of mortality. Different efforts are ongoing to fight malaria disease, and one of the strategies is vaccination. Two malaria vaccines, RTS,S/AS01E (Mosquirix^®^) and R21/Matrix-M^™^, are recommended by the World Health Organization for widespread use among children living in regions with moderate to high Pf malaria transmission [1]. However, since duration and level of protection are not optimal [2, 3], next-generation vaccines are being developed. One candidate with promising results is the Sanaria PfSPZ-CVac vaccine, which consists of intravenous (IV) injection of Pf sporozoites (PfSPZs) together with chloroquine as a chemoattenuating drug. In a clinical trial conducted in Tübingen (Germany) PfSPZ-CVac immunization resulted in full protection against controlled human malaria infection (CHMI) 8–10 weeks after the last vaccine dose [4], a finding that could be reproduced in subsequent trials [5, 6].
CHMI, consisting of administrating drug-sensitive Pf and closely monitoring parasitemia to administer antimalarial drugs at the first sign of infection, is a powerful tool for evaluation of the efficacy of novel malaria vaccines and drugs (reviewed in [7]). CHMI provides also a unique opportunity to study in depth the early immune response to Pf infection.
Immunity to clinical malaria is slowly acquired over time through repeated natural infections; however, sterile immunity to Pf infection is rarely developed. As a result, in highly endemic areas adults frequently have asymptomatic and submicroscopic infections, i.e. low parasitemias only detected by qPCR [8]. Moreover, naturally acquired immunity to clinical malaria may diminish when frequent exposure to Plasmodium spp. is discontinued, with several studies reporting declines of Pf-specific antibodies within months and even weeks after an initial potent response [9–12] in contrast to the antibody responses against many viruses and non-replicating antigens used in vaccines that last decades [9, 13]. In this regard, we and others have demonstrated clear associations between high levels of exposure to Plasmodium spp. and alterations in peripheral B lymphocyte populations that are responsible for antibody production. An expansion of CD10^+^ immature B cells has been seen in children with acute episodes of Pf malaria, which reached up to 25% of the CD19^+^ B lymphocyte subset [14]. The expansion of immature B cells has also been described in HIV [15] and other human immunodeficiency states [16] implying they are associated with chronic immune stimulation. In addition, a subset of CD21^−^CD27^−^ memory B cells (MBCs), termed as “exhausted” or “atypical” MBC are expanded in malaria, HIV and other infections [17–23]. It is not clear whether these malaria-related atypical MBCs are dysfunctional compared to classical MBCs. On the one hand, atypical MBCs have a transcriptional profile distinguishable from the classical ones [24], with a reduced expression of genes involved in BCR signaling [25] and antigen recognition [26], and lower surface IgG levels [23, 25, 27]. On the other hand, signs of proliferative activity and secretion of antibodies have been described in atypical MBCs [27]. It also remains uncertain if a single or only repeated exposure to malaria parasites is sufficient to induce these alterations in B cell subsets. Most studies have been performed in malaria endemic regions, but one study reported an expansion of atypical MBCs in malaria-naïve Dutch individuals after a single CHMI [28]. In addition, the levels of chemokine CCL11 (eotaxin) have been negatively correlated with atypical MBC [23], as well as P. vivax infection and Pf parasitemia [29, 30]. Furthermore, frequencies of atypical MBCs had a positive correlation with serum concentrations of the proinflammatory cytokines tumor necrosis factor (TNF) and interleukin (IL)-8 in a cohort with a lifelong exposure to malaria [23].
The objectives of this work were 1) to compare the B cell profiles of individuals with different Pf immunity status: malaria-naïve, PfSPZ-CVac-immunized and malaria semi-immune individuals with lifelong exposure to Pf in Africa and 2) to evaluate the effect of a single Pf infection via CHMI on these groups.
MATERIALS AND METHODS
Study participants
This study was performed in the context of three CHMI trials (Fig.1) that differed in the level of malaria exposure of the participants: malaria lifelong exposed individuals (LACHMI-001 trial), malaria naïve individuals (TÜCHMI-001 trial) and malaria naïve PfSPZ-CVac vaccinated individuals (TÜCHMI-002 trial). The protocols have been published previously [4,31,32].
The LACHMI-001 clinical trial (NCT02237586) [31] was designed to study the effect of lifelong malaria exposure as well as sickle cell trait on healthy adults aged 18–30 years following experimental infection with 3,200 PfSPZs (strain NF54) administered by direct venous inoculation (DVI) at CERMEL (Gabon) from July 2014 to February 2016. Volunteers with no history of antimalarial drug intake were allocated into three groups: non-immunes (N=5) minimally exposed to malaria (Europeans living temporally in Gabon), and semi-immunes with a lifelong malaria exposure (Gabonese individuals from Lambaréné), further segregated depending on the presence (hemoglobin [Hb] AS, N=9) or absence (HbAA, N=11) of sickle cell trait. Peripheral blood was collected one day before (C-1) and five (D5) and 11 (D11) days after the CHMI, and peripheral blood mononuclear cells (PBMCs) were cryopreserved. Non-immune participants were treated when parasitemia was detected, and semi-immunes when parasitemia and clinical symptoms were detected or at D28 if they were not treated before [31]. In this study eight semi-immune subjects with sufficient PBMCs were analyzed (HbAS, n=5; HbAA, n=3).
The TÜCHMI-001 clinical trial (NCT01624961) [32] aimed to determine the best dose of PfSPZs administered by intravenous injection (IV) through an indwelling catheter for a CHMI regimen. The study was performed at the Eberhard Karls University of Tübingen (Germany) from June to December 2012, with a verification group with the highest dose of parasites administrated by DVI at the Hospital Clinic (Barcelona, Spain) from December 2012 to July 2013 (NCT01771848) [33]. Participants were healthy individuals (N=36) age 18-45 years with no history of malaria. Dose-escalation of PfSPZ (NF54) started with 50 and ended with a maximal dose of 3,200 PfSPZ. Peripheral blood was collected one day before the PfSPZ inoculation (C-1), 11 (D11), and 84 (D84) days after, and PBMCs were cryopreserved [32]. The number of individuals analyzed here was seven, all from the maximal dose group (3,200 PfSPZ) from TÜCHMI-001 clinical trial.
The TÜCHMI-002 clinical trial (NCT02115516) [4] aimed at establishing a safe and well-tolerated vaccine regimen using three different doses by DVI (3.2x10^3^, 1.28x10^4^ and 5.12x10^4^) of purified, cryopreserved PfSPZs administered to malaria-naïve healthy adult volunteers taking chloroquine (PfSPZ-CVac vaccine). To assess vaccine efficacy, placebo and PfSPZ-CVac immunized individuals underwent CHMI 8–10 weeks later with 3200 PfSPZ (NF54) by DVI. The trial was carried out in Tübingen (Germany) from May to July of 2014. Peripheral blood was collected at C-1, D11, D84, and PBMCs were cryopreserved [4]. The number of subjects analyzed here was 33 (placebo, n=12; PfSPZ-CVac vaccinated, n=21).
Ethical approval
For all studies, written informed consent was obtained from all participants. The TÜCHMI-001 and TÜCHMI-002 studies were approved by the ethics committee of the University Clinic and the Medical Faculty of the University of Tübingen, and the Hospital Clinic and the Hospital de la Santa Creu i Sant Pau ethics committees in Barcelona. The LACHMI-001 study received approval by the Gabonese National Ethics Committee (Comité National d’Ethique de la Recherche) and was conducted under the USA FDA Investigational New Drug application. All these studies strictly followed the principles of the Declaration of Helsinki in its sixth revision as well as the Good Clinical Practice guidelines.
Detection of Pf
Pf infection was diagnosed by thick blood smear (TBS) microscopy and by 18S qPCR. From day 5 until antimalarial treatment, blood samples were withdrawn daily. Quantitative TBS were prepared as described elsewhere [34] and qPCR was performed after all the samples were collected, using published protocols [4,31,32].
Isolation of plasma and PBMCs
Plasma was separated from the cellular fraction of heparinized peripheral whole blood by centrifugation at 600 g for 10 min at room temperature within 2–4 h of collection, aliquoted and stored at −80°C. A density gradient medium (Lymphoprep 4X250mL, PALEX, Cat 1114545) was used to obtain PBMCs that were frozen in fetal bovine serum with 10% of dimethyl sulfoxide and stored in liquid nitrogen until analysis.
Immunophenotyping and gating strategy
PBMC samples were thawed and processed at ISGlobal (Spain) in three periods of time for logistic reasons: i) all LACHMI-001 and 11 TÜCHMI-002 samples, ii) 2 TÜCHMI-001 and 7 TÜCHMI-002 samples and iii) 5 TÜCHMI-001 and 15 TÜCHMI-002 samples. Samples were thawed by adding RPMI 1640 medium supplemented with 10% fetal bovine serum (Life Technologies) and 0.5μL/mL benzonase (Novagen-Merck, Cat 70664-3), and were centrifuged 500xg for 7 min at RT. After thawing, PBMC viability was measured on a Guava^®^ Cytometer (PC550IG-C4C / 0746-2747) using ViaCount Reagent (Merck-Millipore, Cat 4000-0041). Samples with viabilitites below 70% were excluded.
Between 5 × 10^5^ and 1 x 10^6^ PBMCs per sample were used for B cell staining. Cell suspensions were stained with LIVE/DEAD^®^ Fixable Aqua Dead Cell Stain Kit (Invitrogen, Cat L34957), washed and blocked with bovine serum albumin 0.5% for 15 min. After washing, cells were stained with an 11-color panel. Supplementary Table 1 summarizes the antibodies used. For compensation control, BD Comp Beads (BD, Cat 552843) were used. To establish the gates for positive events, Fluorescence Minus One (FMO) controls were performed which consisted on staining samples with all the fluorophores used in the panel except one of them. The gating strategy is summarized in Figure 1.
First, leukocytes were gated using a time event gate and selecting singlets. Viable B cells (VBCs) were gated by CD19-expression and by excluding CD3^+^, CD14^+^, CD16^+^, dead and apoptotic cells. Immature VBCs were gated as CD10^+^ within CD19^+^ live cells, whereas mature VBCs were CD10^−^. Mature VBC were divided into switched (IgD^−^), unswitched (IgD^+^ CD38^−/low^), plasmablast cells and germinal center cells (PCGCs) (IgD^−^ CD38^++^) populations. Of note, PCGCs can also include pre-germinal cells and recent germinal cells. Switched and unswitched populations were further segregated by their expression of CD21 and CD27. Within the switched population, MBCs were classified as active classical (acMBCs) (CD27^+^ CD21^−^), resting classical (rcMBCs) (CD27^+^ CD21^+^), active atypical (aaMBCs) (CD27^−^ CD21^−^) and resting atypical (raMBCs) (CD27^−^ CD21^+^). The unswitched (IgD^+^) population was classified as resting naïve (CD27^−^ CD21^+^) and active naïve (CD27^−^ CD21^−^) B cells. B cell subpopulations are reported here as percentage of total VBCs. CD1c^+^, IgG^+^ and PD1^+^ cells were gated within each B cell subset. Cell acquisition was performed on a BD LSR II Fortessa cytometer and the data analysis was performed on FlowJo software version v10.
Antibody and cytokine analyses
Plasma samples were collected 1–2 days before CHMI (C-1), and on the following days after: D7, D11-13, D19 (only in the LACHMI-001 cohort), D28 (only in the TÜCHMI-001 and LACHMI-001 cohorts), and D84 (only for TÜCHMI-001 and TÜCHMI-002). Anti-IgG to 21 Pf antigens (Supplementary Table 2) were measured by quantitative suspension array technology using the xMAP^™^ platform (Luminex Corp., Austin, Texas) and their levels were expressed as median fluorescence intensity, as described [35,36] & [Gómez-Pérez et al, submitted]. Plasma cytokines were measured by means of the xMAP^™^ technology (Luminex Corp., Austin, Texas) at time points C-1, D7, D13, D19 and D28 using the Cytokine Human Magnetic 30-Plex Panel from Life Technologies^™^ as described before [30]. The kit included the following proteins representing major cytokine families (Th1, Th2, Th17), chemokines (proinflammatory and regulatory) and growth factors: epidermal growth factor, fibroblast growth factor, granulocyte colony-stimulating factor, granulocyte-macrophage colony-stimulating factor, hepatocyte growth factor, vascular endothelial growth factor, TNF, interferon (IFN)-α, IFN-γ, IL-1 receptor agonist (RA), IL-1β, IL-2, IL-2R, IL-4, IL-5, IL-6, IL-7, IL-8, IL-10, IL-12(p40/p70), IL-13, IL-15, IL-17, IFN-γ induced protein (IP-10), monocyte chemoattractant protein (MCP-1), monokine induced by IFN-γ (MIG), macrophage inflammatory protein (MIP)-1α, MIP-1β and regulated on activation normal T cell expressed and secreted (RANTES) and CCL11 (eotaxin). Concentrations in pg/mL were log_10_ transformed for analysis.
Statistical analysis
Cell population frequencies were calculated with respect to the VBCs, therefore, alterations in one population were detected as changes on the other cell subsets. To improve statistical power, all malaria-naïve individuals (TÜCHMI-001 and placebo TÜCHMI-002) were grouped and termed “malaria naïve”.
The distributions of quantitative variables were analyzed using the skewness and kurtosis tests for normality. Most frequencies of B cells did not follow a Gaussian distribution (data not shown) and therefore non-parametric statistical tests were chosen for the analysis. Baseline frequencies of B cell subsets were compared between the three exposure groups (naïve, vaccinated and semi-immune) by Kruskal-Wallis followed by Dunn’s test, corrected with the Bonferroni method for multiple comparisons. The effect of the CHMI was evaluated independently in each exposure group by means of the Friedman’s test, followed by two-by-two comparisons with Bonferroni’s correction. To assess whether previous vaccination affected the B cell changes induced by CHMI, we performed generalized estimated equations and explored the interaction between time and group allocation (placebo vs. vaccine) in the TÜCHMI-2 cohort. The same analysis was performed to evaluate the interaction between time and sickle cell trait (HbAA vs HbAS) in the LACHMI-001 cohort. To analyze the association between the B cell frequencies and other population variables and infection after CHMI, individual or multivariable logistic regressions were estimated with the samples from LACHMI and the PfSPZ-CVac-vaccinated group, estimating the odds ratios (OR) and 95% confidence intervals (CI).
The correlations between B cell frequencies and antibody levels or cytokine concentrations were assessed by Spearman’s test at baseline with the naïve, semi-immune and vaccinated donors data together and separately.
Statistical significance was defined at p<0.05, and at p<0.1 a trend was considered. All the statistical analyses and graphs were performed using Stata v17 (Stata Corp., College Station, TX, USA, 2017), SPSS version 28.0.1.0 (IBM Corp., Armonk, N.Y., USA) and R (corrplot and tidyverse packages)[37,38].
RESULTS
Population
The clinical characteristics of study participants are summarized in Table 1. There were more males than females and the median body mass index was in the normal range. After CHMI, all the individuals in the TÜCHMI-001 and placebo-TÜCHMI-002 groups became parasitaemic, while only 29% of the vaccinated-TÜCHMI-002 did. In the LACHMI cohort (semi-immune), 75% of the individuals got a positive smear and 88% were positive by qPCR. In addition, all the individuals with HbAA developed microscopic parasitaemia, while among the participants with HbAS, 40% and 80% got positive smears and qPCR, respectively.
Baseline B cell frequency differences between exposure groups
Compared to naïve, semi-immune participants had higher percentages of immature B cells, aaMBC, acMBC and active naïve B cells, and lower percentages of PCGC at baseline (Table 2). However, some of these differences were not statistically significant after adjusting for multiple testing. In addition, for all B cell subsets, semi-immune donors had higher frequencies of CD1c-expressing cells than naïve individuals (Table 2). Similar results were observed when semi-immune were compared to vaccinated individuals. In contrast to CD1c-expressing cells, semi-immune individuals had lower percentages of IgG^+^ cells for many B cell subsets compared to naïve and vaccinated donors, except aaMBCs, for which they had a higher percentage (Table 2). With regards PD1-expressing cells, only aaMBCs showed different frequencies among exposure groups, with semi-immune donors having more PD1^+^ aaMBC cells than naïve individuals. No differences in the frequencies of B cell subsets were observed between naïve and vaccinated individuals, with the exception of a trend towards lower IgG^+^ aMBC (both resting and active) in vaccinated than naïve individuals (Table 2).
In summary, cumulative but not a single (vaccine) malaria exposure was associated with increased frequencies of many B cell subsets, with higher and lower percentages of CD1c and IgG expressing-cells respectively. The aaMBC subset presented a different behavior, as semi-immune donors had more IgG^+^ aaMBC and PD1^+^aaMBC cells than vaccinated and naïve individuals, respectively.
Effect of CHMI on each malaria-exposure group and B cell subset
In all the cohorts, CHMI was associated with an expansion of rcMBCs, which occurred sooner (D11) in the vaccinated and semi-immune groups (Figures 2A, 2B and 2C). This was accompanied by an expansion of PCGCs and a decrease of naïve B cells in the semi-immune cohort (Figure 2C). With regards to the expression of markers, an expansion of raMBCs-PD1^+^ and active naive-PD1^+^ B cells was observed in the naïve and vaccinated groups, respectively (Figures 2A and 2B). Finally, we observed an increase in the percentage of several MBCs producing IgG^+^ from day 0 to day 11 in the semi-immune group (Figure 2C). Non-significant results are not shown.
Next, the effect of the interaction of previous vaccination with the average B cell changes during follow-up was assessed in the TÜCHMI-002 cohort. There was a significant (or borderline significant) and positive effect of vaccination for rcMBCs (b=0.190, 95%CI 0.011–0.368, p=0.037) (Figure 3) at D11, and for active naïve-PD1^+^ (b=0.637, 95%CI 0.058–1.217, p=0.031) (Figure 3), acMBC-IgG^+^ (b=0.166, 95%CI −0.025–0.358, p=0.088) and PCGCs (b=0.874, 95%CI −0.137–1.884, p=0.090) at D84. In addition, a negative effect was observed for CD1c^+^raMBC (b=−0.328, 95%CI −0.621–−0.032, p=0.030) (Figure 3) at D84 and CD1c^+^ naive B cells (b=−0.201, 95%CI −0.408–0.006, p=0.058).
Furthermore, the interaction of the sickle cell trait (HbAS) with the average B cell changes after the CHMI was assessed in the LACHMI cohort. During the follow-up, semi-immune individuals with sickle cell trait had more aaMBCs (b=1.244, 95%CI 0.041–2.446, p=0.043) and raMBCs (b=0.752, 95%CI 0.036–1.467, p=0.039) at D5 compared to participants with normal hemoglobin (HbAA). On the contrary, sickle cell trait had a negative effect on acMBC-PD1^+^ (b=−0.853, 95%CI −1.395–−0.310, p=0.002), raMBC-PD1^+^ (b=−0.926, 95%CI −1.445–−0.406, p<0.001) and rcMBC-PD1^+^ (b=−0.962, 95%CI −1.679–−0.245, p=0.009) frequencies. These results were maintained after adjusting for malaria infection post-CHMI (data not shown).
Predictors of infection after CHMI
This was evaluated in the vaccinated and semi-immune groups together. Semi-immune donors had 17x higher risk of having an infection after the CHMI than vaccinated donors, but none of the B cell subsets in their baseline levels were significantly associated with infection after CHMI (Table 3). At D11, aaMBC frequency had a borderline non-significant positive association with infection (Table 3). However, after adjusting for previous malaria exposure, the association was lost (OR=2.45, 95% CI 0.67–8.94, p=0.172). When we performed this analysis separately in the vaccinated and semi-immune groups, no associations of baseline or D11 B cells levels and infection were found (data not shown).
Relationship between B cells, Pf antibody levels, and cytokine concentrations
A negative correlation was found between anti-IgG to Pf antigens that are well known markers of exposure (particularly PfMSP-1_19_, PfMSP-1_42_, PfAMA-1) and raMBC or rcMBC frequencies, in all individuals together, at baseline (rho range −0.61 ⊠ −0.25, raw p<0.05, Figure S1A) and all time points together (rho range −0.44 ⊠ −0.1, raw p<0.05, Figure S1B). In contrast, a positive association was observed with active naïve B cell proportions (baseline rho range 0.4 ⊠ 0.24, all timepoints together rho range 0.41 ⊠ 0.19, raw p<0.05). In addition, a moderate negative correlation was found between Pf antibody levels (mostly PfMSP-1s and PfAMA-1s) and IgG^+^ B cell frequencies, while a positive moderate correlation occurred between such antibodies and CD1c^+^ B cell frequencies (Figure 4A, Figure S1). Similar results were observed at D11 after the CHMI but not at D84 (data not shown).
The antibody markers of Pf exposure PfMSP-1_19_, PfMSP-1_42_, PfAMA-1, PfEXP-1, PfEBA-175 also correlated negatively with the concentrations of eotaxin, MCP-1, IP-10, IFN-g and other cytokines when considering all groups and time points together, but mostly at baseline (Figure 4B, Figure S2). In turn, these cytokines/chemokines (mostly eotaxin) correlated moderately and negatively with the frequencies of CD1c^+^ B cells, and positively with IgG^+^ B cells (Figure 4C, Figure S3).
Collectively, data show that higher Pf exposure, manifested by higher anti-Pf IgG levels (most prominent in the semi-immune), correlated with lower eotaxin and IFN related cytokines, as well as higher frequencies of CD1c^+^ B cells and lower frequencies IgG^+^ B cells.
These correlations, however, were driven mostly by the semi-immune group. In LACHMI-001 semi-immune individuals alone, IgG^+^ acMBC correlated positively with pro-inflammatory cytokines (IL-1, IL-6, IL-12, IL-15, IL-17, FGF; Figure 5A) and negatively with regulatory cytokines (IL-10, IL-13) (Figure S4), while CD1c^+^ aaMBC correlated strongly and positively with anti-PfMSP1_19_ and anti-PfMSP1_42_ IgG levels (Figure 5B). PD1^+^ acMBC also correlated positively with several pro-inflammatory cytokines including IFN-a, IL-1b, IL-6, IL-12, IL-17 and IL-2 in semi-immune participants (Figure 5C). In those individuals, the frequency of aaMBCs correlated negatively with eotaxin levels, but this result did not reach statistical significance (Figure S4).
DISCUSSION
Before the CHMI and compared to naïve individuals, semi-immune but not vaccinated individuals had increased frequencies of immature and active naïve B cells, as well as active classical and atypical MBCs. First acute malaria episodes have been associated with an expansion of plasmablast cells, atypical MBCs and/or naïve B cells [28,39], however the frequencies may decline to baseline levels 35 days after a single malaria infection. This suggests that multiple rather than single malaria exposures are necessary for durable changes in B cell profiles, consistent with our findings. Interestingly, baseline frequencies of CD1c^+^ cells within each B cell subset, were higher in the semi-immune compared to naïve individuals, correlated negatively with eotaxin and positively with anti-Pf antibodies, the latter well-known markers of malaria exposure (reviewed in [40]). CD1c expression in dendritic cells has been positively associated to Pf-malaria exposure and protection [41,42]. However, to the best of our knowledge, the expression of this marker on B cells in relation to malaria has not been previously explored and more studies are necesary to confirm whether it is a marker of exposure. On the other hand, the percentages of IgG^+^ cells were lower in semi-immune than naive donors, for most B cell subsets and, accordingly, a negative correlation of IgG^+^ cells with anti-Pf antibody levels was found. Of note, the decreased frequency of IgG^+^ was not detected for the expanded aaMBC population. This negative association between IgG^+^ cell frequencies and malaria exposure is in contrast with previous results observed in a cohort of pregnant and non-pregnant individuals from Papua New Guinea [23,43], where no association was mostly observed. But pregnancy itself, exposure to both Pf and P. vivax malaria or other population factors rather than malaria exposure may be responsible for that finding.
CHMI resulted in an expansion of rcMBCs in all groups, and of IgG^+^ MBCs at D11 in the semi-immune group, as expected after an infection (reviewed in [44]). Besides, an expansion of certain PD1^+^ B cell populations occurred after the CHMI. We and others have previously demonstrated that malaria exposure/infection is associated with elevated frequencies of PD1^+^ B cells compared to healthy individuals [43,45]. However, in this study no correlation was found between PD1^+^ MBCs and anti-Pf antibody levels. The function of PD1 is not yet much recognized on B cells, whilst it is a well-known inhibitory marker in T cells, associated largely to malaria infection/exposure [46–48], also in NK cells [49]. Although it is classically considered that PD1^+^ T cells are exhausted and associated with impaired parasite control and infection chronicity [50], some studies have reported an active role for leukocytes expressing this marker [49,51]. Indeed, PD1 is expressed in activated cells and is involved in immune-homeostatic mechanisms. In consonance with all these results, previous vaccination with PfSPZ-CVac was associated with increased percentages of rcMBCs and PD1^+^ active naïve B cells over post-CHMI follow-up, compared to placebo treatment. Furthermore, PD1+ acMBC were positively correlated with inflammatory cytokines in semi-immune individuals suggesting PD1 as a marker of immune activation.
One-third of the population estimated to have the sickle cell trait lives in sub-Saharan Africa, and children with HbAS have decreased susceptibility to clinical malaria [52]. Among the potential explanations for this protection are: i) enhanced naturally-acquired immunity to malaria, ii) impaired growth of Pf on HbAS-erythrocytes or increased splenic removal, and iii) altered surface expression of cytoadherence proteins on infected HbAS-erythrocytes [53]. In our CHMI study, an increase of atypical MBC frequencies was observed at D5 on individuals with the HbAS, and a decrease of PD1^+^ MBCs, even after adjusting for current infection status. Antibody analysis including all semi-immune participants (N=20) from the same study (LACHMI-001) showed that sickle cell trait individuals compared with the semi-immune with normal hemoglobin had significantly lower IgM and IgG4 levels, and a trend of higher IgG1 and IgG3 response against certain pre-erythrocytic and blood stage antigens [Gómez-Pérez et al, submitted]. To the best of our knowledge, the B cell phenotype change after malaria (or any other) infection has not been described in carriers of the sickle cell trait, and more studies are necessary to confirm these findings and the associated antibody response, especially considering our restricted sample size.
The negative correlation of Pf antibodies with eotaxin is in line with the significantly lower eotaxin levels found in lifelong malaria exposed semi-immune individuals compared to the vaccinated or naïve groups (Moncunill et al, in preparation). Other cytokines related to IFN responses, were also negatively correlated with exposure, suggesting a broad impact of cumulative malaria episodes on the immune profile of individuals towards a tolerogenic profile. Consistently these cytokines were negatively associated with B cell subsets expressing CD1c, which increased frequencies were linked to malaria exposure. Ex vivo studies show that human B cells-activation via BCR crosslinking, but not by CD40L alone (T-cell help), upregulate CD1c in naïve, memory and marginal zone-like B cells, and that elevated CD1c enhanced their function as antigen presenting cells [54]. Hence, CD1c expression might be a marker of B cell activation via BCR crosslinking by Pf antigens, and could be related to a regulatory and tolerogenic role given the observed negative correlation with IFN related cytokines. Accordingly, the semi-immune, especially individuals with sickle cell trait, were asymptomatic or had less malaria symptoms than the malaria-naïve [31]. However, we did not find the previously observed negative correlation between eotaxin and atypical MBCs in endemic populations [23]. This may be explained because in this study individuals with long and short exposure to malaria were included, and the semi-immune cohort was actually too small (n=8) to observe significant differences. Nevertheless, in those individuals, frequencies of MBC expressing IgG, diminished by exposure with the exception of aaMBC, were positively correlated with cytokines related to inflammation, Th1 and Th17 responses, suggesting a more tolerogenic and blunt immune status at baseline related with the decreased frequencies of these B cell subsets.
This study is limited by the small sample size of the cohorts and the need to treat individuals at first detection of parasites, which might have influenced the natural immune response. Another limitation is having analyzed the cellular samples in three different periods of time, which may have hampered comparability. Nevertheless, important strengths of our study include its novelty, being the first to analyze the B cell phenotype dynamics as a response to CHMI in individuals with different immunity and hemoglobin status, as well as the analysis of B cell surface markers not previously studied in the context of malaria such as CD1c.
In summary, cumulative but not a single (vaccine) malaria exposure was associated with increased frequencies of many B lymphocyte subsets, with higher and lower percentages of CD1c and IgG expressing cells, respectively, and a decrease of circulating cytokines previously linked to altered MBC phenotypes, mostly eotaxin, overall suggesting a tolerogenic profile. The CHMI was associated with an early expansion of rcMBC in all three cohorts, and this was increased in the vaccine- compared to the placebo-recipients, suggesting boosting of B cell memory in the vaccinated. The effect of CHMI on B cell phenotypes in semi-immune individuals was also modified by the sickle cell trait. The correlation of these B cell subsets with anti-Pf antibodies suggests that this effect is truly driven by the intensity of exposure to Pf parasites and supports further studies to assess the impact of those changes on the humoral response and naturally acquired immunity against malaria.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Geneva: World Health Organization. World malaria report 2024: addressing inequity in the global malaria response. [Internet]. 2024. Available from: https://www.wipo.int/amc/en/mediation/%0Ahttps://www.who.int/teams/global-malaria-programme/reports/world-malaria-report-2023
- 2RTS, S Clinical Trials Partnership*. Efficacy and safety of RTS,S/AS 01 malaria vaccine with or without a booster dose in infants and children in Africa: Final results of a phase 3, individually randomised, controlled trial. The Lancet. 2015;386:31–45.10.1016/S 0140-6736(15)60721-8PMC 562600125913272 · doi ↗ · pubmed ↗
- 3Datoo MS, Natama HM, SoméA, Bellamy D, TraoréO, Rouamba T, Efficacy and immunogenicity of R 21/Matrix-M vaccine against clinical malaria after 2 years’ follow-up in children in Burkina Faso: a phase 1/2b randomised controlled trial. Lancet Infect Dis. 2022;22:1728–36.36087586 10.1016/S 1473-3099(22)00442-X · doi ↗ · pubmed ↗
- 4Mordmüller B, Surat G, Lagler H, Chakravarty S, Ishizuka AS, Lalremruata A, Sterile protection against human malaria by chemoattenuated Pf SPZ vaccine. Nature 2017 542:7642. 2017;542:445–9.10.1038/nature 21060 PMC 1090648028199305 · doi ↗ · pubmed ↗
- 5Mwakingwe-Omari A, Healy SA, Lane J, Cook DM, Kalhori S, Wyatt C, Two chemoattenuated Pf SPZ malaria vaccines induce sterile hepatic immunity. Nature [Internet]. 2021 [cited 2025 Feb 25];595:289–94. Available from: https://pubmed.ncbi.nlm.nih.gov/34194041/10.1038/s 41586-021-03684-z PMC 1112724434194041 · doi ↗ · pubmed ↗
- 6Sulyok Z, Fendel R, Eder B, Lorenz FR, Kc N, Karnahl M, Heterologous protection against malaria by a simple chemoattenuated Pf SPZ vaccine regimen in a randomized trial. Nat Commun [Internet]. 2021 [cited 2025 Feb 25];12. Available from: https://pubmed.ncbi.nlm.nih.gov/33947856/10.1038/s 41467-021-22740-w PMC 809706433947856 · doi ↗ · pubmed ↗
- 7Stanisic DI, Mc Carthy JS, Good MF. Controlled Human Malaria Infection: Applications, Advances, and Challenges. Infect Immun. 2017;86:479–96.10.1128/IAI.00479-17PMC 573679828923897 · doi ↗ · pubmed ↗
- 8Pierce SK, Miller LH. World Malaria Day 2009: what malaria knows about the immune system that immunologists still do not. J Immunol. 2009;182:5171–7.19380759 10.4049/jimmunol.0804153 PMC 2779769 · doi ↗ · pubmed ↗