An Isospora Species (Apicomplexa: Eimeriidae) Identified From a Black‐Faced Cuckoo‐Shrike (Coracina novaehollandiae) (Gmelin, 1789) (Passeriformes: Campephagidae) in Western Australia
Yinhua Chen, Belinda Brice, Bruno P. Berto, Qiong Li, Rongchang Yang

TL;DR
Scientists discovered a new species of Isospora parasite in a black-faced cuckoo-shrike in Western Australia using both physical and genetic analysis.
Contribution
The discovery of the first coccidian species from the Campephagidae family in Australia.
Findings
The new Isospora species has distinct morphological features including bi-layered oocysts and specific sporocyst characteristics.
Genetic analysis shows high similarity to I. serinuse across 18S rRNA, 28S rRNA, and COI loci.
Phylogenetic analysis confirms the new species is closely related to I. serinuse.
Abstract
We describe and characterize a new Isospora species from the black‐faced cuckoo‐shrike ( Coracina novaehollandiae ) in Western Australia, using both morphological and molecular approaches. Microscopic analysis of a fecal sample revealed 20 ellipsoidal oocysts, which are subspheroidal to ovoid, measuring 40–43 × 39–41 μm (mean 41.4 × 39.6 μm) with a length/width (L/W) ratio of 1.0–1.1 (mean 1.04). The oocyst wall is bi‐layered (~1.5 μm thick), with a smooth outer layer constituting approximately two thirds of the total thickness. A micropyle is present, characterized by a slight invagination of the inner layer (~6.0 μm wide), but no micropyle cap is observed. Both the oocyst residuum and polar granule are absent. Sporocysts (n = 20) are ellipsoidal, measuring 23–24 × 13–14 μm (mean 23.3 × 13.4 μm) with an L/W ratio of 1.7–1.8 (mean 1.74). A flattened to knob‐like Stieda body (~1.5 × 3.0…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
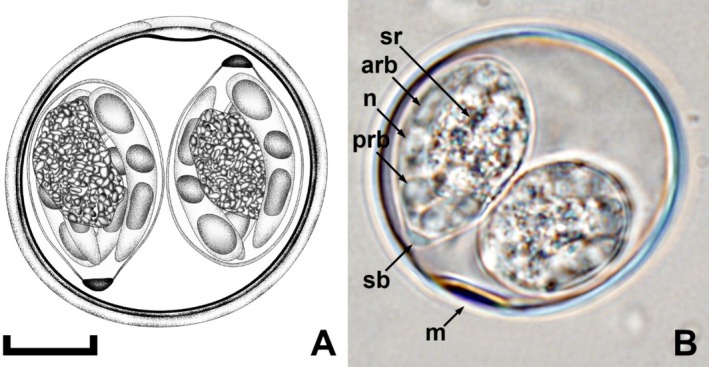 FIGURE 1
FIGURE 1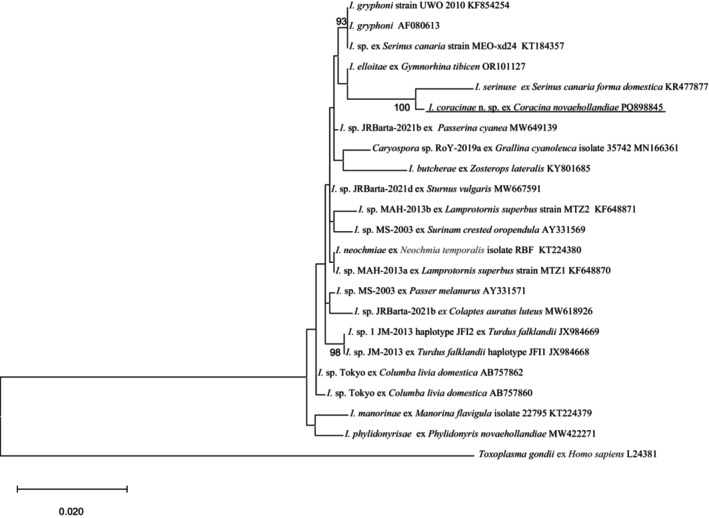 FIGURE 2
FIGURE 2 FIGURE 3
FIGURE 3 FIGURE 4
FIGURE 4|
| Hosts | References | Oocysts | Sporocysts | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Shape | Measurements (um) | Shape index | Wall (um) | Polar granule | Oocyst residuum | Shape | Measurements | Stieda body | Substieda body | Residuum | |||
|
|
| Yang et al. ( | Subspherical | 23.4 × 20.7 (20.0–26.0 × 19.0–22.0) | 1.1 | Bi‐layered c. 0.8 | − | − | O | 14.5 × 10.1 (11.0–17.0 × 9.0–11.0) | Hemi‐dome | Rectangular‐shaped | Compact |
|
|
| Trachta et al. ( | Spherical to subspherical | 17.8 × 16.9 (16–19 × 16–18) | 1 | One‐layered c. 1.0 | − | − | E | 13.2 × 10.8 (12–14 × 9–12) | Tiny | Absent | Scattered granules |
|
|
| Box ( | Subspherical to ellipsoidal | 24.6 × 21.8 (17–30 × 17–30) | 1.1 | Tri‐layered c. 1.2 | + | − | Lemon | 18.1 × 11.5 (17.0–22.0 × 1.00–13.0) | Nipple‐like | 2.0 × 3.0 | Compact |
|
|
| This study | Subspheroidal to ovoidal | 41.4 × 39.6 (40–43 × 39–41) | 1.04 | Bi‐layerd c. 1.5 | − | − | E | 23.3 × 13.4 (23–24 × 13–14) | Knob‐like | Absent | Scattered granules |
|
|
| Trachta et al. ( | Spherical to subspherical | 24.6 × 23.6 (22–26 × 22–25) | 1 | Bi‐layerd c. 1.5 | − | − | O | 13.2 × 10.9 (15–17 × 10–13) | Small | Absent | Scattered granules |
|
|
| McQuistion ( | Ellipsoidal | 27.3 × 23.6 (22–30 × 20–27) | 1.2 | Bi‐layered c. 1.5 | + | − | O | 15.2 × 10.2 (15.0–16.0 × 9.0–11.0) | Nipple‐like | Small | Scattered granules |
|
|
| Yang et al. ( | Subspherical | 20.7 × 18.7 (19.8–21.6 × 18–19.6) | 1.1 | Bi‐layered c. 1.5 | + | − | O | 12.6 × 9.7 (11.9–13.2 × 8.9–10.8) | Flattened to half‐moon | Indistinct | Compact |
|
|
| McQuistion and Wilson ( | Subspheroidal | 20.4 × 20.1 (20–23 × 18–23) | 1 | One‐layered | − | − | O | 14 × 9.5 (13–15 × 8–10) | Small | Small | Irregular‐shaped |
|
|
| McQuistion and Wilson ( | Subspheroidal | 25.3 × 24.2 (24–27 × 23–25) | 1 | One‐layered | + | − | Piriform | 15.4 × 11.5 (14–17 × 11–12) | Knob‐like | Prominent | Irregular‐shaped |
|
|
| Olson et al. ( | Spherical | 29.2 × 30.7 (25.0–33.0 × 28.0–34.0) | 1 | Bi‐layered c. 0.8 | + | − | O | 22.2 × 13.4 (15–25.0 × 12.0–14.5) | Small | Indistinct | Prominent |
|
|
| Yang et al. ( | Subspherical to ellipsoidal | 24.8 × 22.2 (24.0–25.0 × 21.0–23.0) | 1.12 | Bi‐layered c. 1.0 | + | − | O | 15.7 × 10.8 (15.0–16.0 × 10.0–11.0) | Knob‐like | Rounded to trapezoidal | Compact |
|
|
| Yang et al. ( | Subspheroidal | 30.6 × 29.4 (27–34 × 26–31) | 1.04 | Bi‐layered c. 1.0 | + | − | O | 18.3 × 10.7 (17–19 × 10–12) | Flattened to rounded | Rounded to rectangular | Compact |
|
|
| Yang et al. ( | Spherical to subspherical | 22.8 × 18.3 (20.3–23.8 × 17.7–18.7) | 1.2 | Bi‐layered c. 1.3 | + | − | Lemon | 15.53 × 9.7 (14.6–15.73 × 9.5–9.7) | Hemi‐dome | Rectangular‐shaped | Compact |
|
|
| Yang et al. ( | Spherical | 18.3 × 18.2 (18.2–18.9 × 18.2–18.6) | 1 | Bi‐layered c. 1.2 | + | − | O | 13.3 × 8.6 (9.5–16.4 × 6.8–10.0) | Indistinct | Absent | Compact |
|
|
| Trachta et al. ( | Subspherical to broadly ellipsoidal | 24.3 × 19.8 (22–26 × 18–22) | 1.2 | One‐layered c. 1.5 | + | − | O | 15.7 × 10.1 (14–18 × 8–12) | Distinct | Distinct | Spherical |
|
|
| Yang et al. ( | Subspheroidal | 29.8 × 29.4 (29–32 × 28–31) | 1.01 | Bi‐layered c. 1.5 | + | − | O | 18.4 × 12.3 (18–19 × 12–14) | Flatted | Rounded | Scattered granules |
|
|
| McQuistion and Wilson ( | Subspheroidal | 20.9 × 20.8 (20–24 × 19–23) | 1 | One‐layered | + | − | O | 15 × 9.7 (13–16 × 9–10) | Knob‐like | Prominent | Round |
|
|
| Box ( | Spherical to subspherical | 20.1 × 19.2 (13.0–23.03 × 13.0–23.0) | 1 | Tri‐layered c. 1.2 | − | − | E | 15.2 × 9.4 (13.0–16.0 × 8.0–11.0) | 2.0 × 0.6 | 5.0 × 3.0 | Scattered granules |
|
|
| Yang et al. ( | Spherical to subspherical | 25.5 × 23.5 (24.4–27.0 × 22.0–24.8) | 1.09 | Bi‐layered c. 1.2 | + | − | Lemon | 18.9 × 11.8 (17.8–20.2 × 10.6–13.0) | Small | Indistinct | Compact |
|
|
| Yang et al. ( | Spherical | 23.8 × 22.5 (22–24.5 × 21.8 × 24.5) | 1.06 | Bi‐layered c. 1.0 | − | + | O | 14.4 × 11.2 (11.5–15.8) × (10.4–12.5) | Hemi‐dome | Rectangular‐shaped | Compact |
|
|
| McQuistion and Wilson ( | Subspheroidal | 25.4 × 21.1 (21–30 × 17–23) | 1.2 | One‐layered | + | − | Piriform | 15 × 10 (14–15 × 9–11) | Knob‐like | Prominent | Round |
|
|
| Madani et al. ( | Spherical to subspherical | 23.3 × 22.3 (18.5–30 × 18.1–29.3) | 1.05 | Bi‐layered c. 1.3 | − | − | O | 13.9 × 9.3 (10.2–17.5 × 6.5–12.2) | Flatted | Rounded | Compact |
|
|
| Chen et al. ( | Ellipsoidal | 23.4 × 18.7 (21–25 × 18–20) | 1.25 | Bi‐layered c.1.0 | + | − | E | 14.1 × 8.7 (14–15 × 8–9) | Flatted | Rounded | Compact |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCoccidia and coccidiosis research · Bird parasitology and diseases · Parasitic Infections and Diagnostics
Introduction
1
The black‐faced cuckoo‐shrike ( Coracina novaehollandiae Gmelin, 1789) is a passerine bird that is widespread in Australia and Tasmania (Taylor and Bonan 2020). The adult bird has a black face and throat, dove‐gray head, white and dove‐gray plumage on its back and wings, and a black tail with a white tip. A hooked black beak is used to feed on a wide variety of invertebrates and insects, fruits, and seeds. It is a member of the Campephagidae family and has been assigned to the Coracina genus along with 21 other species (Gill and Donsker 2019). These slender birds often shuffle their wings when they land, which has given rise to them also being called “shufflewings.” They are also known as Gray jays or Cherry hawks (Pizzey and Knight 1997).
The most common coccidia to infect passerine birds are the Isospora (Duszynski et al. 1999), with more than 500 species reported (Madani et al. 2018). Isospora are members of the Apicomplexa phylum, a group of parasitic protists, belonging to the Eimeriidae family. Many wild and captive birds shed coccidian oocysts in their feces. Isospora infections are not commonly associated with causing disease in free‐living birds; however, they may result in severe illness and death in some cases. Although there are many Isospora spp. infecting wild birds, only a relatively small number have been both morphologically and genetically characterized (Carreno and Barta 1999; Schrenzel et al. 2005). To date, there has only been one unnamed Isospora species reported in the Campephagidae, that from a Pied triller ( Lalage nigra Pennant, 1781) by Hegner and Chu (1930).
Taxonomic classification of Isospora is becoming more reliant on a combination of both morphological and molecular techniques, including DNA sequencing (Berto et al. 2011).
Isospora species that have been both morphologically and genetically described from wild birds in Australia include Isospora lesouefi from the endangered regent honeyeater ( Xanthomyza phrygia Shaw, 1794), which is endemic to south‐eastern Australia (Morin‐Adeline et al. 2011); Isospora anthochaerae from the red wattlebird ( Anthochaera carunculata Shaw, 1790) (Yang et al. 2014a); Isospora streperae from a gray currawong ( Strepera versicolor Latham, 1801) (Yang et al. 2015a); Isospora manorinae from a yellow‐throated miner ( Manorina flavigula wayensis Gould, 1840) (Yang et al. 2016); Isospora butcherae from a silvereye ( Zosterops lateralis Latham, 1801) (Yang et al. 2018); Isospora coronoides from an Australian raven ( Corvus coronoides Vigors and Horsfield, 1827) (Liu et al. 2019); Isospora lugensae from a Kerguelen petrel ( Lugensa brevirostris Lesson, 1831) (Yang et al. 2021a); Isospora elliotae from an Australian magpie ( Gymnorhina tibicen Latham, 1801) (Yang et al. 2023) and Isospora virescensae from a singing honeyeater (Gavicalis virescens Vieillot, 1817) (Chen et al. 2025).
In this study, we describe a species of Isospora identified in a black‐faced cuckoo‐shrike from Western Australia and characterize it both morphologically and genetically.
Materials and Methods
2
Sample Collection
2.1
An adult, black‐faced cuckoo‐shrike was admitted to the Wattle Grove Veterinary Hospital, Perth after it was found by a member of the public with a broken wing. Surgery was performed to pin the fractured humerus. The bird was subsequently transferred to the Kanyana Wildlife Rehabilitation Centre (KWRC) for further treatment and rehabilitation. A fecal sample was obtained soon after admission to KWRC and screened for intestinal parasites.
Morphological Analysis
2.2
Numerous embryonated nematode eggs and occasional coccidian oocysts were seen on a wet mount. A small portion of feces was placed in a 2% (w/v) potassium dichromate solution (K_2_Cr_2_ O_7_) and emulsified. This emulsion was stored at 4°C until it was transported to Murdoch University for further study. On arrival at Murdoch University (within 24 h), a thin layer of the emulsion was poured into a Petri dish and stored at room temperature (20°C–22°C), in a dark cupboard. The oocysts were screened daily to assess sporulation, using the 100x oil immersion objective of an Olympus CH‐2 binocular microscope, in combination with an ocular micrometer.
The bird was treated with a broad‐spectrum benzimidazole anthelmintic as well as toltrazuril, an anticoccidial agent, and was successfully released near the found location 6 weeks later.
DNA Isolation
2.3
Total DNA was extracted from 200 mg of each fecal sample using the PowerSoil DNA Kit (MolBio, Carlsbad, California) with slight modifications as described by Yang et al. (2016). Specifically, the samples underwent four freeze–thaw cycles, alternating between liquid nitrogen and boiling water, to enhance oocyst lysis before proceeding with the manufacturer's protocol.
PCR Amplification
2.4
A nested PCR approach was used to amplify the 18S rRNA gene, 28S rRNA locus, and partial COI gene sequence. For the 18S rRNA gene, primers EiGTF1 (5′‐TTC ACA GGA CCC TCC GAT C) and EiGTR1 (5′‐AAC CAT GGT AAT TCT ATG G) were employed for external amplification, yielding a product of ~1510 bp, while primers EiGTF2 (5′‐TTA CGC CTA CTA GGC ATT CC) and EiGTR2 (5′‐TGA CCT ATC AGC TTT CGA CG) were used for the internal reaction. The PCR reaction mix included 2.5 μL of 10× Kapa PCR buffer, 2 μL of 25 mM MgCl2, 1.0 μL of 10 mM dNTPs, 10 pM of each primer, 1 unit of KapaTaq (Geneworks, Adelaide, SA), 1 μL of DNA (~50 ng) for the external reaction (or 1 μL of external PCR product for the internal reaction), and 16.4 μL of H_2_O. The cycling conditions consisted of an initial denaturation at 94°C for 3 min, followed by 40 cycles of 94°C for 30 s, 55°C for 30 s, and 72°C for 2 min, with a final extension at 72°C for 5 min.
The 28S rRNA locus was amplified using nested PCR with external primers 28SExF (5′‐TAC CCG CTG AAC TTA AGC) and 28SExR (5′‐CMA CCA AGA TCT GCA CTA G), producing a 1362 bp product (Schrenzel et al. 2005). Internal primers 28SInF (5′‐ACT ATG TTC CCT AGT AAC G) and 28SInR (5′‐AAC GCT TCG CCA CGA TCC) generated an amplicon of 1420 bp (Yang et al. 2014a). The reaction mix contained 2.5 μL of 10× Kapa PCR buffer, 2 μL of 25 mM MgCl2, 1 μL of 10 mM dNTPs, 10 pM of each primer, 1 unit of KapaTaq, 1 μL of DNA (~50 ng), and 16.9 μL of H_2_O. Both primary and secondary reactions followed identical cycling conditions: 94°C for 3 min, followed by 35 cycles of 94°C for 30 s, 60°C for 30 s, and 72°C for 90 s, with a final extension at 72°C for 5 min.
The partial COI gene sequence (723 bp) was also amplified using nested PCR with external primers COIF1 (Ogedengbe et al. 2011) and COXR1 (Dolnik et al. 2009) and internal primers COIF2 (Yang et al. 2013) and COXR2 (Dolnik et al. 2009). The reaction mix contained 2.5 μL of 10× Kapa PCR buffer, 2 μL of 25 mM MgCl2, 1.0 μL of 10 mM dNTPs, 10 pM of each primer, 1 unit of KapaTaq, 1 μL of DNA (~50 ng), and 13.4 μL of H_2_O. PCR cycling conditions for both external and internal reactions involved an initial denaturation at 94°C for 3 min, followed by 40 cycles of 94°C for 30 s, 58°C for 30 s, and 72°C for 1 min, concluding with a final extension at 72°C for 5 min.
Sequence Analysisss
2.5
Amplicons from the second‐round PCRs were gel‐purified using an in‐house filter tip method as outlined by Yang et al. (2013). The purified PCR products were sequenced in both forward and reverse directions, with duplicate sequencing performed using amplicons from independent PCR runs. Sanger sequencing was conducted using an ABI Prism Dye Terminator Cycle Sequencing Kit (Applied Biosystems, Foster City, California) in accordance with the manufacturer's guidelines.
The sequencing data were analyzed and edited using FinchTV v1.4.0 (http://www.geospiza.com/Products/finchtv.shtml). The resulting sequences were compared to Isospora and other coccidian parasite sequences available in GenBank through BLAST searches and aligned with reference sequences using BioEditor (http://bioeditor.sdsc.edu/download.shtml).
Phylogenetic Analysis
2.6
Phylogenetic trees for Isospora spp. were constructed using partial 18S rDNA, 28S rDNA, and COI gene sequences, aligned with additional isolates retrieved from GenBank. Distance analyses and phylogenetic inferences were performed using MEGA‐X (Barry 2013). Sanger sequencing chromatogram files were imported into MEGA‐X, where the nucleotide sequences for each gene were curated, analyzed, and aligned with reference sequences from GenBank using Clustal W (http://www.clustalw.genome.jp).
Maximum likelihood (ML) trees were generated after determining the most appropriate nucleotide substitution models: TN93 + G + I for 18S and 28S, and GTR + G + I for the COI gene. Bootstrap support values were calculated based on 1000 replicates. Genetic similarities among sequences were also computed using MEGA‐X.
Line Drawing
2.7
The line drawings underwent editing using two software applications from CorelDRAW (Corel Draw Graphics Suite, Version 2020, Corel Corporation, Canada), specifically Corel DRAW and Corel PHOTO‐PAINT (Yang et al. 2021a).
Results
3
Description of the Oocyst
3.1
Oocysts (n = 20) subspheroidal to ovoidal, 40–43 × 39–41 (41.4 × 39.6); length/width (L/W) ratio 1.0–1.1 (1.04). Wall bi‐layered, ~1.5 thick, outer layer smooth, c. 2/3 of total thickness. Micropyle present with a slight invagination of the inner layer, ~6.0 wide. Micropyle cap absent. Oocyst residuum and polar granule absent. Sporocysts (n = 20) ellipsoidal, 23–24 × 13–14 (23.3 × 13.4); length/width (L/W) ratio 1.7–1.8 (1.74). Stieda body present, flattened to knob‐like, ~1.5 × 3.0; sub‐Stieda body and para‐Stieda body absent; sporocyst residuum present, composed of granules clustered by a membrane, in an irregular shape, ~12.0 × 8.0. Sporozoites with anterior and posterior refractile bodies and nucleus (Figures 1 and 2).
Composite line drawing and photomicrograph of a sporulated oocyst of I. coracinae n. sp. from a black‐faced cuckoo‐shrike ( Coracina novaehollandiae ) in Western Australia. Note the anterior (arb) and posterior (prb) refractile bodies, micropyle (m), nucleus (n), sporocyst residuum (sr), and Stieda (sb) body. Scale bar: 10 μm.
Type host: Black‐faced cuckoo‐shrike ( Coracina novaehollandiae Gmelin, 1789).
Type locality: Perth (−31.9522°S, 115.8589°E), Western Australia.
Prevalence: 1/1 (100%).
Prepatent period: Unknown.
Patent period: Unknown.
Site of infection: Unknown.
Sporulation time: 48–72 h.
Phylogenetic Analysis
3.2
18S rRNA
3.2.1
Isospora coracinae n. sp. exhibited 98.8% sequence similarity with I. serinuse (GenBank accession: KR477877.2), followed by 98.7% similarity with I. elliotae (OR101127), based on pairwise comparisons. As illustrated in Figure 2, I. coracinae n. sp. clustered with two Isospora species from Western Australia, namely I. serinuse and I. elliotae, forming a distinct subgroup with I. serinuse (Yang et al. 2015b). Pairwise genetic distance analysis between I. coracinae n. sp. and 22 reference species/isolates revealed the shortest genetic distance with I. serinuse (0.0117), followed by I. elliotae (0.0133) (Table S1).
Evolutionary relationships of I. coracinae n. sp. inferred by maximum likelihood analysis (ML) of 18S rDNA sequences (1212 bp). Percentage support (> 70%) from 1000 pseudoreplicates from the ML analysis is indicated at the left of the support nodes.
28S rRNA
3.2.2
Isospora coracinae n. sp. exhibited 93.7% sequence similarity with I. serinuse (KR477878) based on pairwise comparisons. As shown in Figure 3, I. coracinae n. sp. and seven other Isospora species identified from passerine birds in WA—namely I. serinuse (KR477878), I. coronoideae (MK530654), I. anthochaerae (KF766053), I. lunulatae (MW776413), I. phylidonyrisae (MW422270), I. manorinae (KT22438), and I. virescensae (PQ108888)—clustered into distinct groups. Isospora coracinae n. sp. and I. serinuse formed a subgroup within a separate clade, which was parallel to the clade comprising the other six Isospora species from WA. Furthermore, I. coracinae n. sp. showed the shortest genetic distance with I. serinuse (0.0627), followed by an unnamed Isospora species from a Grosbeak starling in the USA (AY283866), with a genetic distance of 0.0718 (Table S2).
Evolutionary relationships of I. coracinae n. sp. inferred by ML of 28S rDNA sequences (1332 bp). Percentage support (> 70%) from 1000 pseudoreplicates from the ML analysis is indicated at the left of the nodes.
COI Gene
3.2.3
Similar to the results from the 18S and 28S rRNA analyses, Isospora coracinae n. sp. exhibited the highest sequence similarity with I. serinuse (KR477878) at 98.9%, based on pairwise comparisons. As shown in Figure 4, I. coracinae n. sp. and I. serinuse formed a subgroup within a distinct clade, which included a pellet subgroup formed by four unnamed Isospora isolates from passerine birds in the USA (OL999112 to OL999115). Furthermore, I. coracinae n. sp. showed the shortest genetic distance with I. serinuse (0.0112), followed by an unnamed Isospora species from a palila ( Loxioides bailleui ) in the USA (OL999112), with a genetic distance of 0.0227 (Table S3).
Evolutionary relationships of I. coracinae n. sp. inferred by ML of partial cox1 gene sequences (633 bp). Percentage support (> 70%) from 1000 pseudoreplicates from the ML analysis is indicated at the left of the nodes.
Discussion
4
Sporulated oocysts of Isospora coracinae n. sp. are morphologically distinct from previously characterized Isospora species recorded from Passeriformes in Oceania (Trachta et al. 2006; Yang et al. 2014a, 2015a, 2015b, 2015c, 2016, 2021a, 2021b, 2023; Chen et al. 2025). As shown in Table 1, the oocyst dimensions of I. coracinae n. sp. (41.4 × 39.6 μm) are larger than those of any previously reported Isospora species listed in Table 1.
The comparison of sequences at three loci revealed that I. coracinae n. sp. consistently exhibited the highest genomic similarities with I. serinuse at the 18S, 28S rRNA, and COI loci, with similarities of 98.8%, 93.7%, and 98.9%, respectively. Phylogenetic analysis also placed I. coracinae n. sp. within the same clade as I. serinuse across all three loci. However, the grouping of I. coracinae n. sp. and I. serinuse varied among the three trees, with other Isospora species present in surrounding clades. For example, at the 18S rRNA locus, I. elloitae, identified in the Australian magpie ( Gymnorhina tibicen ), appeared at the base of the clade containing I. coracinae n. sp. and I. serinuse. Additionally, a subgroup containing three sequences from Canadian birds was observed, including one sequence from Serinus canaria (KT184357), identified in a domestic canary, and two sequences from I. gryphoni (AF080613 and KF854254). At the 28S rRNA locus, the clade containing I. coracinae n. sp. and I. serinuse formed a larger group with six additional Isospora species identified from passerine birds in Western Australia. At the COI locus, the clade of I. coracinae n. sp. and I. serinuse grouped with four unnamed Isospora isolates (OL999112 to OL999115) identified from passerine birds in the USA.
Many coccidian species, including Isospora species, are regarded as being highly host‐specific, typically parasitizing a single host species (Knight et al. 2018). However, exceptions exist, as some coccidian parasites infect multiple host species (Betro et al. 2008; Yang et al. 2014b), even across different genera (McAllister et al. 2024). The host of I. coracinae n. sp., the black‐faced cuckoo‐shrike ( Coracina novaehollandiae ), is a native bird species in Australia. In contrast, I. serinuse parasitizes the domestic canary ( Serinus canaria ), a domesticated form of the wild canary. Although the origins of canaries as cage birds remain somewhat unclear, it is believed that they were initially collected from the Canary Islands (Snow et al. 1997).
Conclusion
5
Despite the high genomic similarities at the 18S, 28S rRNA, and COI loci between I. coracinae n. sp. and I. serinuse, their evolutionary paths are entirely distinct. Furthermore, the significant morphological differences in oocysts between the two species support the conclusion that I. coracinae is a new Isospora species.
Author Contributions
Yinhua Chen: investigation (equal), writing – original draft (equal). Belinda Brice: conceptualization (supporting), investigation (equal), methodology (equal), resources (lead), writing – original draft (equal), writing – review and editing (equal). Bruno P. Berto: formal analysis (equal), validation (equal). Qiong Li: methodology (equal), supervision (equal), writing – review and editing (supporting). Rongchang Yang: conceptualization (lead), methodology (equal), supervision (equal), visualization (lead), writing – review and editing (lead).
Conflicts of Interest
The authors declare no conflicts of interest.
Supporting information
Data S1.
Data S2.
Data S3.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Barry, G. H. 2013. “Building Phylogenetic Trees From Molecular Data With MEGA.” Molecular Biology and Evolution 30, no. 5: 1229–1235. 10.1093/molbev/mst 012.23486614 · doi ↗ · pubmed ↗
- 2Berto, B. P. , W. Flausino , D. Mc Intosh , W. L. Teixeira‐Filho , and C. W. Lopes . 2011. “Coccidia of New World Passerine Birds (Aves: Passeriformes): A Review of Eimeria Schneider, 1875 and Isospora Schneider, 1881 (Apicomplexa: Eimeriidae).” Systematic Parasitology 80, no. 3: 159–204. 10.1007/s 11230-011-9317-8.22002022 · doi ↗ · pubmed ↗
- 3Betro, B. P. , L. H. Ribeiro , F. Ildemar , F. Walter , and C. W. Lopes . 2008. “Two new Hosts for Isospora tiesangui Berto, Flausino, Luz, Ferreira & Lopes, 2008 (Apicomplexa: Eimeriidae).” Revista Brasileira de Medicina Veterinária 32, no. 3: 169–171.
- 4Box, E. D. 1975. “Exogenous Stages of Isospora serini (Aragao, 1933) and Isospora canaria sp. n. In the Canary (Serinus canarius Linnaeus).” Journal of Protozoology 22: 165–169.1151906 10.1111/j.1550-7408.1975.tb 05844.x · doi ↗ · pubmed ↗
- 5Carreno, R. , and J. Barta . 1999. “An Eimeriid Origin of Isosporoid Coccidia With Stieda Bodies as Shown by Phylogenetic Analysis of Small Subunit Ribosomal RNA Gene Sequences.” Journal of Parasitology 85, no. 1: 77–83. 10.2307/3285704.10207368 · doi ↗ · pubmed ↗
- 6Chen, Y. , B. Brice , B. Berto , and R. Yang . 2025. “Morphological and Molecular Characterization of a New Isospora (Apicomplexa:Eimeriidae) Species From a Singing Honeyeater (Gavicalis virescens Vieillot, 1817) (Passeriformes:Meliphagidae) in Western Australia.” Ecology and Evolution 15: e 70801. 10.1002/ece 3.70801.39834684 PMC 11742968 · doi ↗ · pubmed ↗
- 7Dolnik, O. V. , V. Palinauskas , and S. Bensch . 2009. “Individual Oocysts of Isospora (Apicomplexa: Coccidia) Parasites From Avian Faeces: From Photo to Sequence.” Journal of Parasitology 95: 169–174.19245285 10.1645/GE-1873.1 · doi ↗ · pubmed ↗
- 8Duszynski, D. W. , S. J. Upton , and L. Couch . 1999. The coccidia of Passeriformes (Isospora spp.). http://biology.unm.edu/biology/coccidia/passer 1.html.