Cytogenetics of insects in the era of chromosome-level genome assemblies
V.A. Lukhtanov, E.A. Pazhenkova

TL;DR
New genomic tools have transformed insect cytogenetics, revealing detailed chromosome structures and evolutionary patterns previously unknown.
Contribution
High-quality genome assemblies enable precise analysis of chromosomal rearrangements and evolution in insects.
Findings
Chromosomal number changes in Lepidoptera occur via simple fusions and fissions at reused breakpoints.
Chromosome fissions correlate with interstitial telomeres inside chromosomes.
Telomeric DNA in insects includes long sequences with retrotransposon interruptions and diverse motifs.
Abstract
Over the past few years, a revolution has occurred in cytogenetics, driven by the emergence and spread of methods for obtaining high-quality chromosome-level genome assemblies. In fact, this has led to a new tool for studying chromosomes and chromosomal rearrangements, and this tool is thousands of times more powerful than light microscopy. This tool has revolutionized the cytogenetics of many groups of insects for which previously karyotype information, if available at all, was limited to the chromosome number. Even more impressive are the achievements of the genomic approach for studying the general patterns of chromosome organization and evolution in insects. Thus, it has been shown that rapid transformations of chromosomal numbers, which are often found in the order Lepidoptera, are most often carried out in the most parsimonious way, as a result of simple fusions and fissions of…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
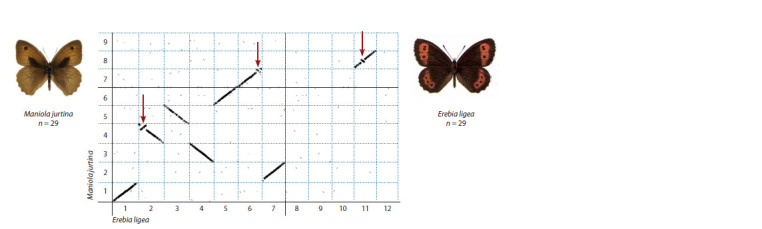 Fig. 1
Fig. 1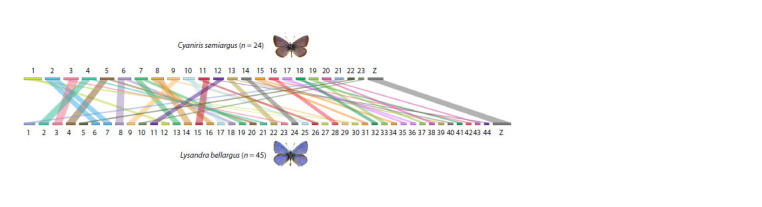 Fig. 2
Fig. 2 Fig. 3
Fig. 3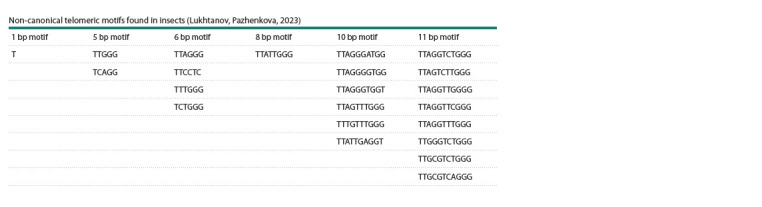 Table 1
Table 1 Fig. 4
Fig. 4Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsChromosomal and Genetic Variations · Genomics and Phylogenetic Studies · Insect Resistance and Genetics
Introduction
Progress in science is often driven by new research methods. In the field of genetics, one such fundamentally new approach, which has given a powerful impetus to the development of the entire discipline, is the DNA sequencing procedure (Heather, Chain, 2016). High-throughput sequencing methods, along with advances in bioinformatics and the development of the Hi-C DNA analysis protocol (Lieberman-Aiden et al., 2009), have recently led to a breakthrough technology for obtaining genome assemblies at the chromosome level (Dudchenko et al., 2017). This methodology has revolutionized comparative cytogenetics, stimulating the emergence of a large number of works devoted to the structure of chromosomes, patterns of chromosomal changes in evolution and role of these changes in speciation. In fact, this new technique has switched the attention of many biologists, especially bioinformaticians, from the analysis of nucleotide substitutions to the analysis of structural changes in DNA, enhancing and complementing the work previously done with a microscope.
The aim of this brief review is to analyze and summarize the advances made in comparative and evolutionary cytogenetics of insects using bioinformatic analysis of chromosome level genome assemblies.
The main stages of insect cytogenetics
Although the karyotypes of some model insect species, such as the midge Chironomus plumosus and the silkworm Bombyx mori, have been studied in great detail (Kiknadze et al., 1991, 2016; Yoshido et al., 2005), for non-model species, information on karyotypes, if available at all, is often limited to the estimation of the diploid (or haploid) number of chromosomes, an approximate description of the size characteristics of individual chromosomes and, less often, individual chromosome arms in the form of a centromeric index (Peruzzi, Eroğlu, 2013). It should be noted that obtaining the latter characteristic is, in principle, impossible for representatives of many insect orders, for example, for butterflies (Lepidoptera) and bugs (Hemiptera), since they have holocentric chromosomes, that is, they do not have a localized centromere (Mandrioli, Manicardi, 2020). Such a low average level of insect cytogenetics is largely due to the objective difficulties of studying chromosomes using a microscope: the sizes of chromosomes are often at the limit of the resolving power of light microscopy
It is therefore not surprising that in the history of cytogenetics, beginning with its inception in the 19th century, attempts have been made to increase the resolving power of cytogenetic analysis. The first stage in the history of cytogenetics can be called the era of chromosome numbers. It arose in the second half of the 19th century, when the first descriptions and images of karyotypes containing the correct determination of the number of chromosomes appeared (e. g., Henking, 1890). The heyday of this era came in the first half of the 20th century, when the study of karyotypes became a mass phenomenon (Beliajeff, 1930; White, 1973).
Significant progress in cytogenetic research was associated with the emergence and widespread use in the second half of the 20th century of methods of differential staining of chromosomes, such as C-banding (Pardue, Gall, 1970) and G-banding (Seabright, 1971). Cytogenetics entered the era of chromosome banding. Almost simultaneously, even more powerful methods of cytogenetic analysis appeared and were developed in parallel, based on the use of the FISH method (Gall, Pardue, 1969; Langer-Safer et al., 1982) and its modifications, such as BAC-FISH (BAC Resource Consortium 2001; Yoshido et al., 2005) and chromosome painting (Schrock et al., 1996; Speicher et al., 1996). This has led to stunning advances in cytogenetics of many groups of organisms, especially vertebrates (Ferguson-Smith, Trifonov, 2007; Graphodatsky et al., 2011). As for insects, with the exception of some model species (Yoshido et al., 2005), this progress has affected them to a lesser extent. Of course, light microscopes have become much better, and the resulting images of karyotypes have become much clearer compared to what they were 100 years ago. In addition, the GISH method has made it possible to effectively detect sex chromosomes (Fukova et al., 2005; Šíchová et al., 2015). Despite this, the cytogenetics of many insect groups, for example, most families of Lepidoptera, is still at the stage of elementary counting of chromosome numbers (Pazhenkova, Lukhtanov, 2023a).
Chromosome level genome assemblies:
a new tool for studying karyotypes
A revolution in the field of karyotype studies has occurred over the past six-eight years. Modern approaches to genome analysis based on obtaining long reads and using Hi-C technology (Dudchenko et al., 2017) make it possible to obtain chromosome level genome assemblies, in which all or at least most of the chromosomes are read from telomere to telomere (Miga et al., 2020; The Darwin Tree…, 2022; Zhang et al., 2023).
Currently, high-quality chromosome level genome assemblies have been obtained for representatives of most insect orders: fleas (Siphonaptera) (Driscoll et al., 2020), stoneflies (Plecoptera) (Dixon et al., 2023), dipterans (Diptera) (Zamyatin et al., 2021; Reinhardt et al., 2023), beetles (Coleoptera) (Van Dam et al., 2021; Huang et al., 2022), springtails (Collembola) (Jin et al., 2023), stick insects (Phasmatodea) (Lavanchy et al., 2024), Hymenoptera (Sun et al., 2021), mayflies (Ephemeroptera) (Farr et al., 2023), Hemiptera (Biello et al., 2021; Mathers et al., 2021; Chen H. et al., 2022; Wang et al., 2024), Orthoptera (Li R. et al., 2024), Trichoptera (Ge et al., 2024), Psocoptera (Feng et al., 2022), Neuroptera (Wang et al., 2022), Odonata (Patterson et al., 2024), Thysanoptera (Yingning et al., 2024), Dermaptera (https://www.ncbi.nlm. nih.gov/datasets/genome/GCA_963082975.1/) and a large number of Lepidoptera species (Mackintosh et al., 2022a; Gauthier et al., 2023; Wright et al., 2024). These assemblies contain information on the haploid number of chromosomes and the size of each chromosome, measured in the number of base pairs. Almost always, there is also information on the presence and size of the sex chromosome X (Z for Lepidoptera and caddisflies, in which females are the heterogametic sex) and, less often, the sex chromosome Y (W for Lepidoptera).
The use of chromosome level genome assemblies has actually led to the emergence of a new methodology and a new tool for studying chromosomes and chromosomal rearrangements. The resolution power of this tool significantly exceeds the capabilities of light microscopy. The basis of the methodology is to obtain pairwise or multiple alignments of chromosome assemblies of different species. These alignments are usually presented in the form of circular plots (Krzywinski et al., 2009). Another analysis option is to obtain pairwise comparisons presented in the form of dot plots (Li H., 2018), in which the nucleotide sequences of individual chromosomes are plotted along the abscissa and ordinate axes, starting with the first, largest chromosome (Fig. 1). Such graphs clearly demonstrate macrosyntenic regions and identify chromosome fusions/fissions, as well as chromosomal inversions. The latter are visible on the graph as segments that are perpendicular to the main diagonals (Fig. 1).
Comparison of the genomes of the butterflies Maniola jurtina and Erebia ligea based on chromosome assemblies.The butterflies M. jurtina and E. ligea have 29 chromosomes in the haploid set. The first 12 chromosomes of E. ligea are mapped on the abscissa axis. The first nine chromosomes of M. jurtina are mapped on the ordinate axis. The diagonals on the graph show the regions of macrosynteny. Inversions are marked with red arrows. It is seen that chromosome 1 of M. jurtina is homologous to chromosome 1 of E. ligea, chromosome 2 of M. jurtina is homologous to chromosome 7 of E. ligea, etc. (according to: Pazhenkova, Lukhtanov, 2023b).
The use of this tool has revolutionized cytogenetics of many insect groups, for which karyotype information, if any, was previously limited to elementary counts of mitotic or meiotic chromosomes. However, even more impressive are the achievements of the genomic approach to the study of general patterns of chromosome organization and karyotype evolution, including the analysis of chromosomal rearrangements and chromosomal syntenies (Biello et al., 2021; Mathers et al., 2021; Sun et al., 2021; Van Dam et al., 2021; Höök et al., 2023; Hundsdoerfer et al., 2023; Wright et al., 2024), as well as the reconstruction of ancestral karyotypes (Chen X. et al., 2023; Wright et al., 2024). Chromosome level genome assemblies have been used to study meiotic drive (Reinhardt et al., 2023; Boman et al., 2024), sex chromosome evolution (Mackintosh et al., 2022b; Berner et al., 2023; Höök et al., 2023, 2024), interspecific transfer of chromosomal inversion during interspecific hybridization (Seixas et al., 2021), the role of chromosomal rearrangements in the evolution of recombination frequency (Näsvall et al., 2023), and to identify genomic coordinates for breakpoints that give rise to chromosomal rearrangements (Zamyatin et al., 2021).
Chromosomal conservatism
and rapid karyotypic evolution
In our work (Pazhenkova, Lukhtanov, 2023a), we used the analysis of chromosome level genome assemblies to solve one of the mysteries of evolutionary cytogenetics. It is known that the chromosome numbers of many insects are conservative and remain unchanged or with minimal changes for tens and hundreds of millions of years (White, 1973). For example, in the order Lepidoptera (butterflies and moths), the ancestral haploid chromosome number n = 31 has been preserved for 200 million years, although n = 30 is often found in some species along with n = 31. In the blue butterflies (the family Lycaenidae), the haploid number n = 24 predominates, although n = 23 is often found (Robinson, 1971). This suggests that large chromosomal rearrangements are rare in the evolution of the order Lepidoptera. At the same time, in some genera of butterflies, there are explosions of karyotypic variability,and chromosome numbers change dramatically in a very short time, for example, during the divergence of two closely related species (White, 1973). The chromosomal mechanisms of such rapid karyotypic evolution were unclear. In addition, it was unclear how real the phenomenon of chromosomal conservatism itself was, since the preservation of the ancestral chromosome number does not exclude intra-chromosomal rearrangements
Analysis of chromosome level genome assemblies in multiple Lepidoptera species showed that in the evolutionary phase of chromosomal conservatism, most autosomes are indeed stable. However, this does not apply to the sex chromosome Z. Fusions of the Z chromosome with one of the autosomes, independently occurring in different evolutionary lineages, lead to multiple variants of the NeoZ chromosome and a decrease in the haploid number by one unit.
As for the explosive karyotypic evolution, the most rapid changes in chromosome numbers are carried out in a parsimonious way: as a result of simple fusions and fissions of chromosomes (Fig. 2). Moreover, these fusions and fissions are not random and can be carried out in different phylogenetic lineages due to the repeated use of the same ancestral chromosomal breakpoints (Pazhenkova, Lukhtanov, 2023a). It should also be noted that the tendency for breaks is correlated with the presence of the so-called interstitial telomeres in chromosomes, i. e. telomere-like structures located not at the ends of chromosomes, but inside them (Fig. 3).
Schematic representation of chromosomes and regions of macrosynteny in the karyotypes of the butterflies Cyaniris semiargus (n = 23+Z) and Lysandra bellargus (n = 44+Z).The karyotype of L. bellargus differs from the karyotype of C. semiargus by chromosome fissions that have occurred in 21 out of 23 autosomes (according to: Pazhenkova, Lukhtanov, 2023a).
Telomere and interstitial telomere structure in chromosome 38 of Lysandra bellargusEach telomere is a long array (CCTAA)n/(TTAGG)n (yellow) interspersed with retrotransposable TRAS (blue) and SART (green) elements. The TRAS and SART elements have long An/Tn and Tn/An tails and are specifically inserted between CCT/ AGG and AA/TT nucleotides of the TTAGG motif (according to: Pazhenkova, Lukhtanov, 2023a).
Telomeric DNA of insects
It is believed that in most insects, telomeric DNA consists of a canonical five-letter TTAGG motif, which is repeated hundreds and thousands of times at the ends of chromosomes (Kuznetsova et al., 2020). However, the analysis of telomeric DNA in 220 insect species in our studies (Lukhtanov, 2022; Lukhtanov, Pazhenkova, 2023), as well as other works that appeared in parallel (Zhou et al., 2022; Fajkus et al., 2023), showed that in addition to the canonical TTAGG motif, insects contain a large number of other variants of telomeric repeats, the length of which varies from 1 to 11 nucleotides (see the Table).
Non-canonical telomeric motifs found in insects (Lukhtanov, Pazhenkova, 2023)
Even more intriguing is the fact that the vast majority of insects have telomeres with a complex multilayer structure (Lukhtanov, Pazhenkova, 2023). In these telomeres, blocks of short telomeric motifs are regularly interrupted by retrotrans posons that are specifically embedded in repeats of both the canonical TTAGG motif (Fig. 3) and other non-canonical motifs. In our opinion, such a structure indirectly indicates the presence of two parallel mechanisms for maintaining telomere length during cell divisions in insects: the classical telomerase mechanism and a mechanism based on transpositions. In general, insects are characterized by a great diversity in the organization of telomeric DNA, which is summarized in Figure 4.
Types of telomeric DNA organization in insects.A – short repeats; B – main short repeats interspersed with variant short repeats, known in the honey bee Apis mellifera (Wallberg et al., 2019); C – main short repeats with insertions of telomere-specific non-LTR retrotransposons of the TRAS and SART families (found in Lepidoptera); D – main short repeats (5–11 bp) with insertions of telomere-specific non-LTR retrotransposons of the SART family (found in most studied species of Hemiptera, Coleoptera and many Hymenoptera); E – main short repeats with insertions of telomere-specific non-LTR retrotransposons of the TRAS family (found in Trichoptera); F – long (173–381 bp) repeats (found in Diptera); G – telomere-specific non-LTR retrotransposons of the HeT-A, TAHRE and TART families (found in Drosophila melanogaster) (Biessmann et al., 2000; Casacuberta, Pardue, 2003).
Prospects for the use of chromosome level
genome assemblies in cytogenetics
The number of high-quality chromosome level genome assemblies available in the GenBank database is growing exponentially due to the activities of various laboratories, and primarily The Wellcome Sanger Institute in the UK (The Darwin Tree…, 2022). Currently, GenBank contains information on chromosome assemblies of genomes (including determination of the haploid number of chromosomes) for 1,118 insect species (https://www.ncbi.nlm.nih.gov/datasets/ genome/?taxon=50557, dated May 15, 2024). Thus, it can already be stated that over the past three-four years, the number of new insect karyotypes, obtained using bioinformatic analysis of genomes, is comparable to or even exceeds the number of karyotypes studied using routine cytogenetic analysis. In addition, chromosomal assemblies of genomes carry several orders of magnitude more information about the obtained karyotypes.
The successes of the genomic approach discussed above do not mean that classical cytogenetics, which provides information on the real spatial configurations of chromosomes, should be discounted. Classical cytogenetics is also needed to validate chromosome level genome assemblies, in particular to confirm the number of chromosomes (Pazhenkova, Lukhtanov, 2023a) and the structure of telomeric DNA motifs (Dalla Benetta et al., 2020; Stoianova et al., 2024). Examples of such validation reinforce the conclusion that chromosome level genome assemblies are a reliable source of information on karyotypes. For instance, for butterflies and moths (order Lepidoptera), information on chromosome assemblies, including determination of the haploid number of chromosomes, is available for 452 species (https://www.ncbi. nlm.nih.gov/datasets/genome/?taxon=7088, from May 15, 2024). For more than half of them, there are data on chromosome numbers obtained using light microscopy methods. Having compared these data, we found complete agreement in the haploid chromosome number calculations made using the bioinformatics approach and light microscopy methods (Pazhenkova, Lukhtanov, 2023a).
Thus, the exceptional prospects of using chromosome level genome assemblies for karyotype analysis are beyond doubt.
Conflict of interest
The authors declare no conflict of interest.
References
BAC Resource Consortium. Integration of cytogenetic landmarks into the draft sequence of the human genome. Nature. 2001;409:953- 958. doi 10.1038/35057192
Beliajeff N.K. Die Chromosomenkomplexe und ihre Beziehung zur Phylogenie bei den Lepidopteren. Z Vererbungsl. 1930;14:369-399. doi 10.1007/BF01848966
Berner D., Ruffener S., Blattner L.A. Chromosome-level assemblies of the Pieris mannii butterfly genome suggest Z-origin and rapid evolution of the W chromosome. Genome Biol Evol. 2023;15(6):evad111. doi 10.1093/gbe/evad111
Biello R., Singh A., Godfrey C.J., Fernández F.F., Mugford S.T., Powell G., Hogenhout S.A., Mathers T.C. A chromosome-level genome assembly of the woolly apple aphid, Eriosoma lanigerum Hausmann (Hemiptera: Aphididae). Mol Ecol Resour. 2021;21(1):316-326. doi 10.1111/1755-0998.13258
Biessmann H., Zurovcova M., Yao J.G., Lozovskaya E., Walter M.F. A telomeric satellite in Drosophila virilis and its sibling species. Chromosoma. 2000;109:372-380. doi 10.1007/s004120000094
Boman J., Wiklund C., Vila R., Backström N. Meiotic drive against chromosome fusions in butterfly hybrids. Chromosome Res. 2024; 32(2):7. doi 10.1007/s10577-024-09752-0
Casacuberta E., Pardue M.L. Transposon telomeres are widely distributed in the Drosophila genus: TART elements in the virilis group. Proc Natl Acad Sci USA. 2003;100:3363-3368. doi 10.1073/pnas. 0230353100
Chen H., Qiao G., Liang A. Chromosome-level genome assembly of Callitettix versicolor (rice spittlebug). Genome Biol Evol. 2022; 14(9):evac130. doi 10.1093/gbe/evac130
Chen X., Wang Z., Zhang C., Hu J., Lu Y., Zhou H., Mei Y., Cong Y., Guo F., Wang Y., He K., Liu Y., Li F. Unraveling the complex evolutionary history of lepidopteran chromosomes through ancestral chromosome reconstruction and novel chromosome nomenclature. BMC Biol. 2023;21:265. doi 10.1186/s12915-023-01762-4
Dalla Benetta E., Antoshechkin I., Yang T., Nguyen H.Q.M., Ferree P.M., Akbari O.S. Genome elimination mediated by gene expression from a selfish chromosome. Sci Adv. 2020;6:eaaz9808. doi 10.1126/sciadv.aaz9808
Dixon A., Macadam C.R., Natural History Museum Genome Acquisition Lab, Darwin Tree of Life Barcoding collective, Wellcome Sanger Institute Tree of Life programme, Wellcome Sanger Institute Scientific Operations: DNA Pipelines collective, Tree of Life Core Informatics collective, Darwin Tree of Life Consortium. The genome sequence of the spring stonefly, Protonemura montana (Kimmins, 1941) [version 1; peer review: awaiting peer review]. Wellcome Open Res. 2023;8:333. doi 10.12688/wellcomeopenres.19790.1
Driscoll T.P., Verhoeve V.I., Gillespie J.J., Johnston J.S., Guillotte M.L., Rennoll-Bankert K.E., Rahman M.S., Hagen D., Elsik C.G., Macaluso K.R., Azad A.F. A chromosome-level assembly of the cat flea genome uncovers rampant gene duplication and genome size plasticity. BMC Biol. 2020;18(1):70. doi 10.1186/s12915-020-00802-7
Dudchenko O., Batra S.S., Omer A.D., Nyquist S.K., Hoeger M., Durand N.C., Shamim M.S., Machol I., Lander E.S., Aiden A.P., Aiden E.L. De novo assembly of the Aedes aegypti genome using Hi-C yields chromosome-length scaffolds. Science. 2017;356(6333): 92-95. doi 10.1126/science.aal3327
Fajkus P., Adámik M., Nelson A.D.L., Kilar A.M., Franek M., Bubeník M., Frydrychová R.Č., Votavová A., Sýkorová E., Fajkus J., Peška V. Telomerase RNA in Hymenoptera (Insecta) switched to plant/ciliate-like biogenesis. Nucleic Acids Res. 2023;51(1):420- 433. doi 10.1093/nar/gkac1202
Farr A., Macadam C.R., Natural History Museum Genome Acquisition Lab; Darwin Tree of Life Barcoding collective; Wellcome Sanger Institute Tree of Life programme; Wellcome Sanger Institute Scientific Operations: DNA Pipelines collective; Tree of Life Core Informatics collective; Darwin Tree of Life Consortium. The genome sequence of the Northern Summer Mayfly, Siphlonurus alternatus (Say, 1824) [version 1; peer review: 2 approved, 3 approved with reservations]. Wellcome Open Res. 2023,8:488. doi 10.12688/ wellcomeopenres.20172.1
Feng S., Opit G., Deng W., Stejskal V., Li Z. A chromosome-level genome of the booklouse, Liposcelis brunnea, provides insight into louse evolution and environmental stress adaptation. GigaScience. 2022;11:giac062. doi 10.1093/gigascience/giac062
Ferguson-Smith M.A., Trifonov V. Mammalian karyotype evolution. Nat Rev Genet. 2007;8(12):950-962. doi 10.1038/nrg2199
Fuková I., Nguyen P., Marec F. Codling moth cytogenetics: karyotype, chromosomal location of rDNA, and molecular differentiation of sex chromosomes. Genome. 2005;48(6):1083-1092. doi 10.1139/g05-063
Gall J.G., Pardue M.L. Formation and detection of RNA-DNA hybrid molecules in cytological preparations. Proc Natl Acad Sci USA. 1969;63(2):378-383. doi 10.1073/pnas.63.2.378
Gauthier J., Meier J., Legeai F., McClure M., Whibley A., Bretaudeau A., Boulain H., Parrinello H., Mugford S.T., Durbin R., Zhou C., McCarthy S., Wheat C.W., Piron-Prunier F., Monsempes C., François M.C., Jay P., Noûs C., Persyn E., Jacquin-Joly E., Meslin C., Montagné N., Lemaitre C., Elias M. First chromosome scale genomes of ithomiine butterflies (Nymphalidae: Ithomiini): comparative models for mimicry genetic studies. Mol Ecol Resour. 2023;23(4):872-885. doi 10.1111/1755-0998.13749
Ge X., Peng L., Deng Z., Du J., Sun C., Wang B. Chromosome-scale genome assemblies of Himalopsyche anomala and Eubasilissa splendida (Insecta: Trichoptera). Sci Data. 2024;11(1):267. doi 10.1038/s41597-024-03097-3
Graphodatsky A.S., Trifonov V.A., Stanyon R. The genome diversity and karyotype evolution of mammals. Mol Cytogenet. 2011;4:22. doi 10.1186/1755-8166-4-22
Heather J.M., Chain B. The sequence of sequencers: the history of sequencing DNA. Genomics. 2016;107(1):1-8. doi 10.1016/j.ygeno. 2015.11.003
Henking H. Untersuchung über die erster Entwicklungsvorgänge in den Eien der Insekten. I. Das Ei von Pieris brassicae L. nebst Bemerkungen über Samen und Samenbildung. Z Wiss Zool. 1890;49:503-564
Höök L., Näsvall K., Vila R., Wiklund C., Backström N. High-density linkage maps and chromosome level genome assemblies unveil direction and frequency of extensive structural rearrangements in wood white butterflies (Leptidea spp.). Chromosome Res. 2023;31(1):2. doi 10.1007/s10577-023-09713-z
Höök L., Vila R., Wiklund C., Backström N. Temporal dynamics of faster Neo-Z evolution in butterflies. Evolution. 2024;78:1554-1567. doi 10.1093/evolut/qpae082
Huang Q., Sim S.B., Geib S.M., Childers A., Liu J., Wei X., Han W., Posada-Florez F., Xue A.Z., Li Z., Evans J.D. Identification of sex chromosomes and primary sex ratio in the small hive beetle, a worldwide parasite of honey bees. GigaScience. 2022;12:giad056. doi 10.1093/gigascience/giad056
Hundsdoerfer A.K., Schell T., Patzold F., Wright C.J., Yoshido A., Marec F., Daneck H., Winkler S., Greve C., Podsiadlowski L., Hiller M., Pippel M. High-quality haploid genomes corroborate 29 chromosomes and highly conserved synteny of genes in Hyles hawkmoths (Lepidoptera: Sphingidae). BMC Genomics. 2023;24(1):443. doi 10.1186/s12864-023-09506-y
Jin J., Zhao Y., Zhang G., Pan Z., Zhang F. The first chromosome-level genome assembly of Entomobrya proxima Folsom, 1924 (Collembola: Entomobryidae). Sci Data. 2023;10(1):541. doi 10.1038/ s41597-023-02456-w
Kiknadze I.I., Shilova A.I., Kerkis I.E., Shobanov N.A., Zelentsov N.I., Grebenyuk L.P., Istomina A.G., Prasolov V.A. Karyotypes and Morphology of Larvae of the Tribe Chironomini. An Atlas. Novosibirsk: Nauka Publ., 1991 (in Russian)
Kiknadze I., Istomina A., Golygina V., Gunderina L. Karyotypes of Palearctic and Holarctic Species of the Genus Chironomus. Novosibirsk: Academic Publishing House “Geo”, 2016
Krzywinski M., Schein J., Birol I., Connors J., Gascoyne R., Horsman D., Jones S.J., Marra M.A. Circos: an information aesthetic for comparative genomics. Genome Res. 2009;19:1639-1645. doi 10.1101/gr.092759.109
Kuznetsova V., Grozeva S., Gokhman V. Telomere structure in insects: a review. J Zool Syst Evol Res. 2020;58:127-158. doi 10.1111/jzs. 12332
Langer-Safer P.R., Levine M., Ward D.C. Immunological method for mapping genes on drosophila polytene chromosomes. Proc Natl Acad Sci USA. 1982;79(14):4381-4385. doi 10.1073/pnas.79.14.4381
Lavanchy G., Brandt A., Bastardot M., Dumas Z., Labédan M., Massy M., Toubiana W., Tran Van P., Luchetti A., Scali V., Mantovani B., Schwander T. Evolution of alternative reproductive systems in Bacillus stick insects. Evolution. 2024;78(6):1109-1120. doi 10.1093/evolut/qpae045
Li H. Minimap2: pairwise alignment for nucleotide sequences. Bioinformatics. 2018;34:3094-3100
Li R., Qin Y., Rong W., Deng W., Li X. Chromosome-level genome assembly of the pygmy grasshopper Eucriotettix oculatus (Orthoptera: Tetrigoidea). Sci Data. 2024;11:431. doi 10.1038/s41597- 024-03276-2
Lieberman-Aiden E., van Berkum N.L., Williams L., Imakaev M., Ragoczy T., Telling A., Amit I., Lajoie B.R., Sabo P.J., Dorschner M.O., Sandstrom R., Bernstein B., Bender M.A., Groudine M., Gnirke A., Stamatoyannopoulos J., Mirny L.A., Lander E.S., Dekker J. Comprehensive mapping of long-range interactions reveals folding principles of the human genome. Science. 2009;326(5950):289-293. doi 10.1126/science.1181369
Lukhtanov V.A. Diversity and evolution of telomere and subtelomere DNA sequences in insects. bioRxiv. 2022. doi 10.1101/2022.04.08. 487650
Lukhtanov V.A., Pazhenkova E.A. Diversity and evolution of telomeric motifs and telomere DNA organization in insects. Biol J Linn Soc. 2023;140:536-555. doi 10.1093/biolinnean/blad068
Mackintosh A., Laetsch D.R., Baril T., Ebdon S., Jay P., Vila R., Hayward A., Lohse K. The genome sequence of the scarce swallowtail, Iphiclides podalirius. G3 (Bethesda). 2022a;12(9):jkac193. doi 10.1093/g3journal/jkac193
Mackintosh A., Laetsch D.R., Baril T., Foster R.G., Dincă V., Vila R., Hayward A., Lohse K. The genome sequence of the lesser marbled fritillary, Brenthis ino, and evidence for a segregating neo-Z chromosome. G3 (Bethesda). 2022b;12(6):jkac069. doi 10.1093/ g3journal/jkac069
Mandrioli M., Manicardi G.C. Holocentric chromosomes. PLoS Genet. 2020;16(7):e1008918. doi 10.1371/journal.pgen.1008918
Mathers T.C., Wouters R.H.M., Mugford S.T., Swarbreck D., van Oosterhout C., Hogenhout S.A. Chromosome-scale genome assemblies of aphids reveal extensively rearranged autosomes and long-term conservation of the X chromosome. Mol Biol Evol. 2021;38(3):856- 875. doi 10.1093/molbev/msaa246
Miga K.H., Koren S., Rhie A., Vollger M.R., Gershman A., Bzikadze A., Brooks S., … Pevzner P.A., Gerton J.L., Sullivan B.A., Eichler E.E., Phillippy A.M. Telomere-to-telomere assembly of a complete human X chromosome. Nature. 2020;585(7823):79-84. doi 10.1038/s41586-020-2547-7
Näsvall K., Boman J., Höök L., Vila R., Wiklund C., Backström N. Nascent evolution of recombination rate differences as a consequence of chromosomal rearrangements. PLoS Genet. 2023;19(8): e1010717. doi 10.1371/journal.pgen.1010717
Pardue M.L., Gall J.G. Chromosomal localization of mouse satellite DNA. Science. 1970;168(3937):1356-1358. doi 10.1126/science.168. 3937.1356
Patterson C.W., Bonillas-Monge E., Brennan A., Grether G.F., Mendoza- Cuenca L., Tucker R., Vega-Sánchez Y.M., Drury J. A chromosome- level genome assembly for the smoky rubyspot damselfly (Hetaerina titia). J Hered. 2024;115(1):103-111. doi 10.1093/ jhered/esad070
Pazhenkova E.A., Lukhtanov V.A. Сhromosomal conservatism vs chromosomal megaevolution: enigma of karyotypic evolution in Lepidoptera. Chromosome Res. 2023a;31:16. doi 10.1007/s10577- 023-09725-9
Pazhenkova E.A., Lukhtanov V.A. Whole-genome analysis reveals the dynamic evolution of holocentric chromosomes in satyrine butterflies. Genes. 2023b;14(2):437. doi 10.3390/genes14020437
Peruzzi L., Eroğlu H.E. Karyotype asymmetry: again, how to measure and what to measure? Comp Cytogenet. 2013;7(1):1-9. doi 10.3897/ CompCytogen.v7i1.4431
Reinhardt J.A., Baker R.H., Zimin A.V., Ladias C., Paczolt K.A., Werren J.H., Hayashi C.Y., Wilkinson G.S. Impacts of sex ratio meiotic drive on genome structure and function in a stalk-eyed fly. Genome Biol Evol. 2023;15(7):evad118. doi 10.1093/gbe/evad118
Robinson R. Lepidoptera Genetics. Oxford: Pergamon Press, 1971
Schrock E., du Manoir S., Veldman T., Schoell B., Wienberg J., Ferguson- Smith M.A., Ning Y., Ledbetter D.H., Bar-Am I., Soenksen D., Garini Y., Ried T. Multicolor spectral karyotyping of human chromosomes. Science. 1996;273(5274):494-497. doi 10.1126/science. 273.5274.494
Seabright M. A rapid banding technique for human chromosomes. Lancet. 1971;2(7731):971-972. doi 10.1016/s0140-6736(71)90287-x
Seixas F.A., Edelman N.B., Mallet J. Synteny-based genome assembly for 16 species of Heliconius butterflies, and an assessment of structural variation across the genus. Genome Biol Evol. 2021;13(7): evab069. doi 10.1093/gbe/evab069
Šíchová J., Voleníková A., Dincă V., Nguyen P., Vila R., Sahara K., Marec F. Dynamic karyotype evolution and unique sex determination systems in Leptidea wood white butterflies. BMC Evol Biol. 2015;15:89. doi 10.1186/s12862-015-0375-4
Speicher M.R., Ballard S.G., Ward D.C. Karyotyping human chromosomes by combinatorial multi-fluor FISH. Nat Genet. 1996;12:368- 375. doi 10.1038/ng0496-368
Stoianova D., Grozeva S., Golub N.V., Anokhin B.A., Kuznetsova V.G. The first FISH-confirmed non-canonical telomeric motif in Heteroptera: Cimex lectularius Linnaeus, 1758 and C. hemipterus (Fabricius, 1803) (Hemiptera, Cimicidae) have a 10 bp motif (TTAGGGATGG) n. Genes. 2024;15(8):1026. doi 10.3390/genes 15081026
Sun C., Huang J., Wang Y., Zhao X., Su L., Thomas G.W.C., Zhao M., … Schaack S., Barribeau S.M., Williams P.H., Waterhouse R.M., Mueller R.L. Genus-wide characterization of bumblebee genomes provides insights into their evolution and variation in ecological and behavioral traits. Mol Biol Evol. 2021;38(2):486-501. doi 10.1093/ molbev/msaa240
The Darwin Tree of Life Project Consortium. Sequence locally, think globally: The Darwin Tree of Life Project. Proc Natl Acad Sci USA. 2022;119:e2115642118. doi 10.1073/pnas.2115642118
Van Dam M.H., Cabras A.A., Henderson J.B., Rominger A.J., Pérez Estrada C., Omer A.D., Dudchenko O., Lieberman Aiden E., Lam A.W. The Easter Egg Weevil (Pachyrhynchus) genome reveals syntenic patterns in Coleoptera across 200 million years of evolution. PLoS Genet. 2021;17(8):e1009745. doi 10.1371/journal.pgen.1009745
Wallberg A., Bunikis I., Pettersson O.V., Mosbech M.B., Childers A.K., Evans J.D., Mikheyev A.S., Robertson H.M., Robinson G.E., Webster M.T. A hybrid de novo genome assembly of the honeybee, Apis mellifera, with chromosome length scaffolds. BMC Genomics. 2019;20:275. doi 10.1186/s12864-019-5642-0
Wang Y., Zhang R., Wang M., Zhang L., Shi C.M., Li J., Fan F., Geng S., Liu X., Yang D. The first chromosome-level genome assembly of a green lacewing Chrysopa pallens and its implication for biological control. Mol Ecol Resour. 2022;22(2):755-767. doi 10.1111/1755-0998.13503
Wang Y., Luo Y., Ge Y., Liu S., Liang W., Wu C., Wei S., Zhu J. Chromosome- level genome assembly of the predatory stink bug Arma custos. Sci Data. 2024;11:417. doi 10.1038/s41597-024-03270-8
White M.J.D. Animal Cytology and Evolution. Cambridge: Cambridge University Press, 1973
Wright C.J., Stevens L., Mackintosh A., Lawniczak M., Blaxter M. Comparative genomics reveals the dynamics of chromosome evolution in Lepidoptera. Nat Ecol Evol. 2024;8(4):777-790. doi 10.1038/ s41559-024-02329-4
Yingning L., Shuhua W., Wenting D., Miao M., Ying W., Rong Z., Liping B. Chromosome-level genome assembly of Odontothrips loti Haliday (Thysanoptera: Thripidae). Sci Data. 2024;11(1):451. doi 10.1038/s41597-024-03289-x
Yoshido A., Bando H., Yasukochi Y., Sahara K. The Bombyx mori karyotype and the assignment of linkage groups. Genetics. 2005; 170:675-685. doi 10.1534/genetics.104.040352
Zamyatin A., Avdeyev P., Liang J., Sharma A., Chen C., Lukyanchikova V., Alexeev N., Tu Z., Alekseyev M.A., Sharakhov I.V. Chromosome- level genome assemblies of the malaria vectors Anopheles coluzzii and Anopheles arabiensis. GigaScience. 2021;10(3): giab017. doi 10.1093/gigascience/giab017
Zhang T., Xing W., Wang A., Zhang N., Jia L., Ma S., Xia Q. Comparison of long-read methods for sequencing and assembly of lepidopteran pest genomes. Int J Mol Sci. 2023;24(1):649. doi 10.3390/ ijms24010649
Zhou Y., Wang Y., Xiong X., Appel A.G., Zhang C., Wang X. Profiles of telomeric repeats in Insecta reveal diverse forms of telomeric motifs in Hymenopterans. Life Sci Alliance. 2022;5:e202101163. doi 10.26508/lsa.202101163