European Pine Marten (Martes martes) as Natural Definitive Host of Sarcocystis Species in Latvia: Microscopic and Molecular Analysis
Petras Prakas, Rasa Vaitkevičiūtė, Naglis Gudiškis, Emilija Grigaliūnaitė, Evelina Juozaitytė-Ngugu, Jolanta Stankevičiūtė, Dalius Butkauskas

TL;DR
European pine martens in Latvia host multiple Sarcocystis species, some of which can infect livestock and cause food poisoning.
Contribution
First molecular identification of Sarcocystis species in European pine martens from Latvia, including a novel species.
Findings
Nine Sarcocystis species were identified in pine marten intestinal samples, including a new species.
Several Sarcocystis species found can infect livestock or cervids and may cause food poisoning.
Co-infections with multiple Sarcocystis species were detected in 30% of the samples.
Abstract
The pine marten (Martes martes) is a species of the family Mustelidae, widely distributed in the Baltic States. This predatory species can harbor a range of pathogens; however, its role in transmitting protozoan Sarcocystis parasites remains understudied. In the current investigation, we aimed to molecularly identify Sarcocystis species from intestinal samples of European pine martens from Latvia, focusing on the diagnosis of zoonotic and pathogenic ones. This is the first study to establish the richness of Sarcocystis species in the intestines of predators from Latvia. Overall, nine Sarcocystis species were identified: eight known ones, using cervids and livestock as their intermediate hosts, and a genetically new species, Sarcocystis sp. 25MmLV, closely related to S. wenzeli, which is known to be pathogenic for chickens. We report the first detection of S. entzeorthi, S. hjorti, S.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
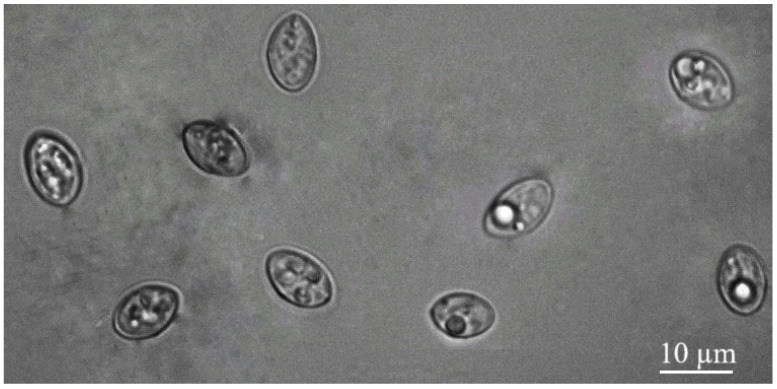 Figure 1
Figure 1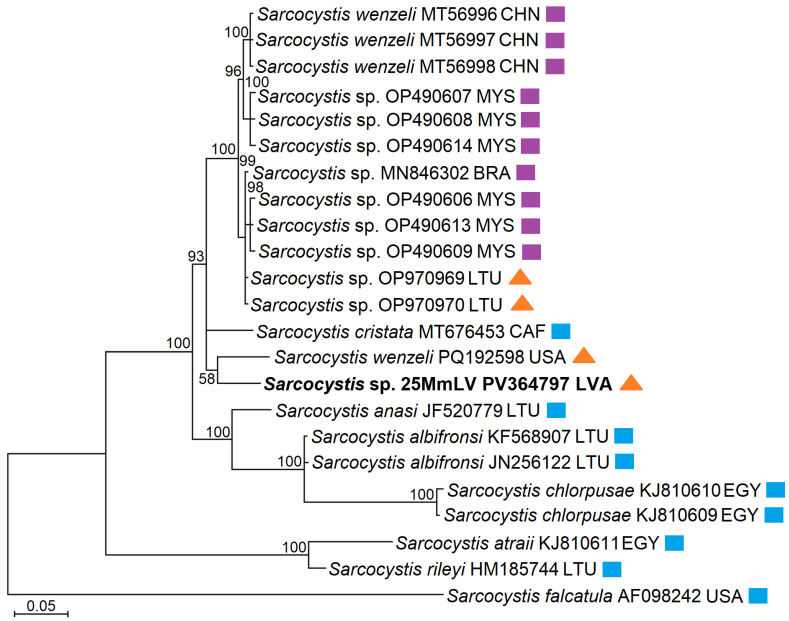 Figure 2
Figure 2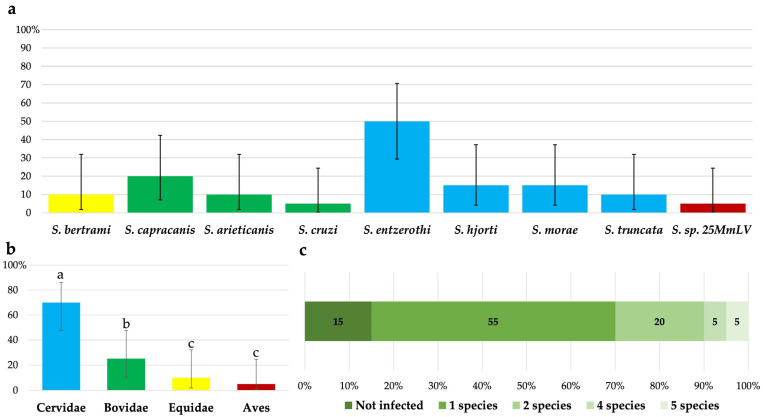 Figure 3
Figure 3Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsToxoplasma gondii Research Studies · Yersinia bacterium, plague, ectoparasites research · Rabies epidemiology and control
1. Introduction
The Mustelidae family represents the most extensive and taxonomically diverse clade within the order Carnivora, encompassing five subfamilies and approximately sixty-seven extant species of terrestrial carnivorous or piscivorous mammals [1]. Mustelids exhibit remarkable ecological adaptability, thriving across diverse climates and seasonal environments. Their tendency to occupy large territories contributes to lower population densities than other carnivores [1,2]. Within the Mustelidae family, food habits vary widely among species, reflecting their adaptability to different environments. Many species hunt in burrows and crevices, while others have evolved specialized behaviors, such as climbing trees (e.g., martens) to pursue their diverse prey [1,3,4]. In Latvia, a total of nine species belonging to the Mustelidae family are found. These include weasels (Mustela erminea), stone martens (Martes foina), European minks (Mustela lutreola), European pine martens (Martes martes), least weasels (Mustela nivalis), badgers (Meles meles), polecats (Mustela putorius), American minks (Mustela vison), and otters (Lutra lutra) [5]. Historically, these animals have played an important role for humans, particularly in the fur trade and in controlling rodent populations [6,7]. The European pine marten is a key fur-bearing species found across the Baltic States. Although data from the Official Statistics of Latvia show a slight decline in the number of hunted animals from 2018 to 2025 (decreasing from 23 thousand to 22 thousand) [8], the precise estimated population of the European pine marten in Latvia remains uncertain. This uncertainty is compounded by the fact that, despite the overall decline in hunted animals, the population of European pine martens has been growing, likely due to reduced hunting pressure, which is largely attributed to a sharp decline in fur prices [9].
The European pine marten is known for its generalist and opportunistic feeding behavior, adapting its diet based on seasonal and local food availability. As a generalist omnivore, it likely occupies the most expansive dietary niche among species in the Mustelidae family [10]. Its diet shows considerable geographic and seasonal variation but commonly includes a variety of mammals, birds, fruits, invertebrates, amphibians, and reptiles [11,12,13]. The European pine marten interacts with a wide range of food sources, which exposes it to various parasites across different ecological niches. Given its diverse diet and ecological role, there is evidence suggesting that the European pine marten, although not extensively studied, may serve as a vector and potential reservoir for several pathogens [14]. To date, there have been documented cases of the European pine marten harboring a range of pathogens, including bacteria (e.g., Leptospira spp., Yersinia spp., Salmonella spp., Helicobacter spp.), protozoa (e.g., Toxoplasma gondii, Neospora caninum, Hepatozoon spp., Sarcocystis spp.), ectoparasites (e.g., Demodex spp., Ixodes spp.), and helminths such as trematodes (Euryhelmis squamula), cestodes (Taenia martis), and nematodes (Crenosoma spp., Toxocara spp., Trichinella spp., Baylisascaris columnaris) [9,15,16,17]. Bacterial pathogens like Leptospira spp., Salmonella spp., and Yersinia spp. are known to cause severe gastrointestinal, systemic, or vector-borne illnesses in humans and wildlife [18,19,20]. Also, the European pine marten contributes to the spread and circulation of important zoonotic protozoa and helminths, such as Toxoplasma gondii, Eucoleus aerophilus, Capillaria hepatica, Angiostrongylus vasorum, Sarcocystis spp., and Trichinella spp., in the environment [9,14]. Furthermore, zoonotic viruses such as SARS-CoV-2, Louping ill virus (LIV), and rotaviruses found in the European pine marten pose risks of cross-species transmission, potentially contributing to emerging infectious diseases [14].
The apicomplexan parasites of the genus Sarcocystis have zoonotic potential and can cause significant health issues in both livestock and wildlife, resulting in economic losses [21]. Sarcocystis spp. exhibit an obligate two-host life cycle, characterized by a prey–predator dynamic. The definitive host (DH), typically a carnivore or omnivore, becomes infected through the ingestion of mature sporocysts present in the muscle tissue of infected prey. Within the DH, Sarcocystis undergoes sexual reproduction, resulting in the release of sporocysts into the environment through fecal excretion, which subsequently contaminates food and water sources. These contaminated resources are consumed by animals serving as intermediate hosts (IHs). Upon ingestion, the sporocysts or oocysts are internalized by the IH, where Sarcocystis undergoes asexual reproduction, leading to the formation of sarcocysts mainly within the muscle tissue of the IH [22]. In recent years, the molecular characterization of Sarcocystis species has advanced significantly through the use of various genetic markers. Among these, the most commonly employed genetic regions in DH studies of Sarcocystis parasites include 18S rRNA, 28S rRNA, ITS1, and cox1 [23,24,25,26]. Notably, the cox1 gene has been shown to provide the highest resolution for differentiating Sarcocystis species infecting ungulates as IHs [27], whereas the ITS1 region has proven to be the most informative marker for species identification when birds serve as IHs [28,29]. To date, over 200 distinct Sarcocystis species have been identified. Humans can become the DH of several Sarcocystis species by consuming beef or pork/wild boar meat [22,30,31]. Additionally, food poisoning has been linked to potential Sarcocystis spp. infections, particularly following the consumption of raw venison or horse meat, resulting in symptoms such as abdominal pain, vomiting, watery diarrhea, loss of appetite, and nausea [32,33]. Sarcocystis spp. primarily affect herbivorous IHs, causing significant tissue damage, increased mortality, and economic losses, while infections in carnivorous or omnivorous DHs are usually mild or asymptomatic, with occasional mild to chronic diarrhea. As such, Sarcocystis parasites pose a greater threat to IHs than to DHs [34,35].
Sarcocystis species exhibit varying degrees of pathogenicity across livestock, with significant implications for animal health and productivity. The pathogenicity of Sarcocystis depends on the species of parasite, the localization in the host, the dose of infection, and the immune status of the host [22]. Sarcocystis species transmitted by canids tend to be more pathogenic compared to those transmitted by other DHs [36]. Sarcocystis cruzi is the most pathogenic species in cattle, causing a range of clinical signs, including fever, anorexia, anemia, neuromuscular dysfunction, abortion, and mortality, with severity influenced by the infectious dose. Conversely, Sarcocystis hirsuta transmitted via felids is considered mildly pathogenic [37]. Typically, Sarcocystis gigantea results in few clinical signs in sheep, with economic losses mainly due to carcass condemnation [38]. However, occasionally, Sarcocystis arieticanis and Sarcocystis tenella can cause miscarriage or acute disease early in the infection, followed by chronic effects like reduced productivity [22]. In goats, Sarcocystis capracanis is the most pathogenic species, leading to fever, weight loss, miscarriage, and death, with survivors remaining unthrifty and prone to secondary infections [39]. Sarcocystis bertrami, which parasitizes horses, generally causes mild or no clinical signs [40]. In pigs, Sarcocystis miescheriana can induce severe symptoms, including weight loss, skin purpura, muscle tremors, miscarriage, and death, depending on the sporocyst load [41,42]. Sarcocystis wenzeli is potentially implicated as the etiological agent of meningoencephalitis in chickens [43,44]. Research on the pathogenicity of Sarcocystis spp. in wild ungulates of the Cervidae family are limited. Nevertheless, a study in Switzerland suggested Sarcocystis hjorti as the etiological agent of eosinophilic fasciitis in red deer [45], emphasizing the need for additional investigations to elucidate the pathogenic impact of various Sarcocystis species on wildlife health.
Recent studies employing molecular techniques have increasingly identified various mustelid species as natural DHs for several Sarcocystis spp. in Lithuania [46,47,48,49]. However, previous investigations tested a limited number of Sarcocystis species, emphasizing the diagnosis of parasite species that have certain IHs, such as cattle [49], cervids [47], birds [48], or livestock and cervids [46]. In the present study, we aimed to molecularly identify Sarcocystis spp. characterized by different IHs and DHs in the intestines of European pine marten collected in Latvia. It should be noted that we have focused on the detection of zoonotic Sarcocystis spp. and species pathogenic to farm animals and wildlife.
2. Materials and Methods
2.1. Sample Collection and Isolation of Sarcocystis spp. Sporocyst
Twenty European pine martens were legally hunted in the northern part of Latvia (main location: 57°20′33.1″ N, 25°25′21.4″ E) in February of 2023. Mustelids were collected in accordance with national and institutional guidelines from licensed third parties. The Cabinet of Ministers’ Hunting Law, Regulation No 421 (22 July 2014, Riga) classifies the European pine marten as an unlimited game animal and allows hunting from 1 October to 31 March. This study did not involve the purposeful hunting of animals. Intestinal samples from European pine martens were collected in collaboration with hunters and delivered to the Laboratory of Molecular Ecology at the Nature Research Centre (Vilnius, Lithuania). The material was preserved at –20 °C and subsequently used for the molecular examination of Sarcocystis species.
Fecal matter was expressed from each intestinal sample, after which the intestine was longitudinally incised. The mucosal surface was gently scraped with a scalpel, and the resulting material was suspended in 100 mL of distilled water. Sporocysts of Sarcocystis species were isolated from the full intestinal scrapings of each European pine marten following the protocol described by Verma et al. 2017 [50], with modifications as detailed in Šukytė et al. 2024 [51].
2.2. Molecular Analysis of Sarcocystis Species
DNA was extracted using the GeneJET Genomic DNA Purification Kit (Thermo Fisher Scientific Baltics, Vilnius, Lithuania), adhering to the provided protocol. The resulting purified DNA was stored at −20 °C for future molecular analyses.
Sarcocystis spp. were detected by amplifying partial cox1 or ITS1 sequences using nested PCR (nPCR). The ITS1 region was used as a genetic marker for Sarcocystis species with avian IHs, while the cox1 gene was employed for species infecting ungulates. The set of genus-specific (I step) and species-specific (II step) primer pairs used in this study is listed in Table 1. It is worth noting that the two most well-studied zoonotic Sarcocystis species (S. hominis and S. suihominis) were tested using species-specific primers in both rounds of nPCR. A total of 18 Sarcocystis species were examined. Of these, seven use members of the family Canidae as their DHs, including S. arieticanis (IH: wild and domestic sheep), S. bertrami (IH: horse), S. capracanis (IH: wild and domestic goats), S. cruzi (IH: cattle), S. hjorti (IH: Cervidae), S. miescheriana (IH: wild boars and pigs), and Sarcocystis morae (IH: Cervidae) [52,53,54,55,56,57,58]. Four species utilize placental predatory mammals as their DHs: Sarcocystis anasi (IH: ducks), Sarcocystis albifronsi (IH: geese), Sarcocystis wenzeli (IH: chicken), and Sarcocystis rileyi (IH: ducks) [59,60]. Meanwhile, Felidae serve as the DHs for S. gigantea (IH: wild and domestic goats) and S. hirsuta (IH: cattle) [57]. Two species employ Mustelidae as DHs, such as Sarcocystis entzerothi (IH: Cervidae) and Sarcocystis truncata (IH: Cervidae) [61,62]. The DHs of Sarcocystis bovifelis (IH: cattle) could be either Felidae or Mustelidae [63,64]. Additionally, zoonotic Sarcocystis hominis (IH: cattle) and Sarcocystis suihominis (IH: pigs and wild boars) use humans as their DHs [30,31]. The specificity of primers used was mainly confirmed in previous studies, except for hjor3/hjor4, which were tested in this study using S. hjorti DNA isolated from red deer (Table 1).
The first round of nPCR was conducted in a total reaction volume of 25 μL, comprising 12.5 μL of DreamTaq PCR Master Mix (Thermo Fisher Scientific, Vilnius, Lithuania), 0.5 μM of each primer (forward and reverse), 4 μL of extracted gDNA, and nuclease-free water to adjust the final volume. The PCR thermal cycling protocol commenced with an initial denaturation at 95 °C for 5 min, followed by 35 cycles consisting of denaturation at 94 °C for 35 s, annealing at a 55–69 °C (determined by the primer pair) for 45 s, and elongation at 72 °C for 55 s, concluding with a final extension step at 72 °C for 5 min. The second round of amplification was carried out in a 25 μL reaction mixture containing 12.5 μL of DreamTaq PCR Master Mix, 0.5 μM of each primer, 2 μL of the PCR product from the initial nPCR round, and nuclease-free water to adjust the final volume. The thermal cycling conditions were maintained identical to those in the initial round. To ensure the reliability of the amplification process, both positive and negative controls were included. Positive controls were prepared using genomic DNA extracted from Sarcocystis spp. sarcocysts identified in previous studies, while negative controls consisted of nuclease-free water instead of DNA.
The quality of the amplified nPCR products was checked using 1% agarose gel electrophoresis. Positive amplicons were purified with ExoI and FastAP (Thermo Fisher Scientific Baltics, Vilnius, Lithuania), following the manufacturer’s instructions. The purified DNA fragments were then sequenced using the same forward and reverse primers as in the nPCR. Sequencing reactions were carried out with the BigDye^®^ Terminator v3.1 Cycle Sequencing Kit and analyzed on a 3500 Genetic Analyzer (Applied Biosystems, Foster City, CA, USA), following the manufacturer’s guidelines. The resulting sequences were manually examined to ensure accuracy, ensuring there were no double peaks or poly signals.
2.3. Phylogentic Analysis
The ITS1 and cox1 sequences obtained in our work were deposited in GenBank with accession numbers PV364797, PV388221−PV388247, respectively. We used the nBLAST sequence similarity search algorithm [70] to compare sequences generated in the present work with those available in NCBI GenBank. BLAST analysis of the ITS1 sequence obtained in this study revealed no significant similarity to any previously described Sarcocystis species, suggesting the presence of a genetically new species. Consequently, phylogenetic analysis was conducted to elucidate the phylogenetic relationships of this genetically new species. A multiple-sequence alignment was performed using a ClustalW algorithm incorporated into a MEGA11 v. 11.0.13 software [71]. The final alignment consisted of 27 sequences and 506 bp including gaps. The selection of nucleotide substitution model best fitting for a resulted alignment and a construction of a phylogenetic tree under a Bayesian inference was carried out using TOPALi v. 2.5 software [72]. The SYM nucleotide substitution model was chosen, and the tree was rooted on Sarcocystis falcatula. Bayesian analysis was performed in two runs, using half a million generations with a sample frequency of 10 and 25% burn-in value. We have used Microsoft Office suite for data visualization.
3. Results
3.1. Microscopical Examination of Sarcocystis spp. Sporocysts
Light microscopy (LM) analysis of intestinal scraping samples detected Sarcocystis spp. infection in 14 out of 20 (70.0%) European pine martens. Under LM, the detected free sporocysts were 8.0–12.9 × 6.1–8.6 µm (mean 10.6 × 7.1 µm; n = 70) in size (Figure 1). No sporulated oocysts or/and oocysts were detected in the intestinal samples analyzed.
3.2. Molecular Identification of Sarcocystis Species
Based on species-specific primers amplifying fragments of cox1, we identified eight known Sarcocystis species: S. arieticanis, S. bertrami, S. capracanis, S. cruzi, S. entzerothi, S. hjorti, S. morae, and S. truncata (Table 2). Other Sarcocystis species tested, including S. anasi, S. albifronsi, S. bovifelis, S. giganta, S. hirsuta, S. hominis, S. rileyi, S. suihominis, and S. wenzeli, were not detected. The comparison of sequences of the same species obtained in the current study showed minor differences (≤0.7%). By contrast, comparing our generated sequences with sequences of the same species available in GenBank, a significantly higher intraspecific variation was obtained, and sequence similarity values were in the range of 96.9–100% for S. bertrami, S. capracanis, S. entzerothi, S. hjorti, S. morae, and S. truncata. Meanwhile, our sequence of S. cruzi demonstrated 95.4–99.5% genetic similarity with those of S. cruzi and ≤90.4% genetic similarity with those of other Sarcocystis spp., and finally, our sequences of S. arieticanis shared 91.3–100% genetic similarity with S. arieticanis and ≤86.3% genetic similarity with those of other Sarcocystis spp. Thus, the calculated intraspecific and interspecific genetic variability values for all detected Sarcocystis species did not overlap, confirming the reliability of the species identification.
Furthermore, a genetically new species, named Sarcocystis sp. 25MmLV, was detected in the intestines of a single European pine marten by using internal SU1F/5.8SR2 and external AZVF1/AZVR1 primers (Table 1). The 461 bp ITS1 sequence of Sarcocystis sp. 25MmLV displayed 92.3% genetic similarity to those of Sarcocystis sp. LT-2022 (OP970969–OP970970) isolated from the intestines of a European pine marten and American mink (Neogale vison) in Lithuania and to that of Sarcocystis sp. isolate Chicken-2016-DF-BR (MN846302) isolated from the brain tissues of chickens in Brazil; 91.2–92.1% genetic similarity to those of S. wenzeli (PQ192598, MT756994–MT756998) obtained from muscle tissues of chickens in China and from the feces of gray wolf (Canis lupus) from the USA; 91.4% genetic similarity to that of Sarcocystis cristata (MT676453) from muscle tissues of a great blue turaco (Corythaeola cristata) in the Central African Republic; and 90.9–91.3% genetic similarity to that of Sarcocystis sp. (OP490606–OP490609, OP490613–OP490614) isolated from the brains and muscles of chickens in Malaysia.
Based on the 461 bp ITS1 sequence analyzed, Sarcocystis sp. 25MmLV was placed, with maximum support, in a cluster consisting of S. wenzeli, S. cristata, S. anasi, S. albifronsi, S. sp., and S. chlorpopusae (Figure 2), which form sarcocysts in the muscle or brain of various birds and are transmitted via placental predatory mammals of the order Carnivora [73]. Sarcocystis sp. 25MmLV was included in a different clade from that composed of S. anasi, S. albifronsi, and Sarcocystis chloropusae. A posterior probability support of 100% was given to group sequences of S. wenzeli (MT756996–MT756998) and Sarcocystis sp. isolated from chickens in China, Malaysia, and Brazil (MN846302, OP490606–OP490609, OP490613–OP490614) and sequences of Sarcocystis sp. isolated from the intestines of a European pine marten and an American mink from Lithuania (OP970969–OP970970). The fragment analyzed was not sufficiently phylogenetically informative to resolve the phylogenetic relationships between Sarcocystis sp. 25MmLV, S. cristata, and Sarcocystis species named S. wenzeli and isolated from the feces of gray wolf (PQ192598). However, based on BLAST analysis, the latter sequence (PQ192598) showed ≤91.9% genetic similarity compared to other available sequences of Sarcocystis spp. In summary, phylogenetic analysis confirmed that the ITS1 sequence obtained in this work belongs to a genetically new Sarcocystis species.
3.3. Distribution of Sarcocystis Species Identified in Intestines of European Pine Martens
Of the 20 European pine martens examined, 13 were positive for Sarcocystis spp. by both microscopical and molecular methods. In two animals, no Sarcocystis spp. were identified; in four European pine martens, Sarcocystis spp. were only established by molecular examination; and finally, in one animal, Sarcocystis spp. sporocysts were observed, but no parasite species was diagnosed by PCR. The most commonly identified Sarcocystis species, S. entzerothi, was found in half of the samples, while the prevalence of other Sarcocystis spp. ranged from 5.0% to 25.0% (Figure 3a). The detection rate of S. entzerothi was significantly higher than compared to that of S. hjorti, S. morae (χ^2^ = 5.58, p < 0.05), S. arieticanis, S. bertrami, S. truncata (χ^2^ = 7.62, p < 0.01), S. cruzi, and Sarcocystis sp. 25MmLV (χ^2^ = 10.16, p < 0.01). In general, significantly more often Sarcocystis spp. with Cervidae as IHs (70.0%) were detected compared to those Sarcocystis spp. using Bovidae (25.0%, χ^2^ = 8.12, p < 0.01), Equidae (10.0%, χ^2^ = 15.00, p < 0.001), or Aves (5.0%, χ^2^ = 26.33, p < 0.001) as their IHs (Figure 3b). Based on molecular examination, 55.0% of the samples were positive for single Sarcocystis species, while co-infections with two or more Sarcocystis species were observed in 30.0% of the animals examined (Figure 3c). Two different Sarcocystis species were identified in 20.0% of European pine martens, and four and five different Sarcocystis species were detected in a single animal each.
4. Discussion
4.1. Identification of Sarcocystis spp. in Mustelidae as DH
Sarcocystis parasites predominantly are found as sarcocysts in the striated muscles of IHs, while in the DHs, they are detected in feces or the intestines as sporocysts or sporulated oocysts [22]. Traditionally, species differentiation has been based on variations in the sarcocyst wall structure within IHs [22,74]. However, this approach is not applicable in DH studies, as sporocysts and oocysts exhibit minimal morphological variation, lack distinct structural features, and barely differ in size, making species-level identification challenging [73,75]. To date, the majority of Sarcocystis species in DHs have been identified through transmission experiments [76,77,78]. However, the application of such experiments is significantly restricted worldwide due to ethical regulations [68]. Consequently, the analysis of fecal matter or intestinal scrapings is emerging as an alternative approach, enabling Sarcocystis identification without the need for live animal experimentation [79,80,81,82]. Using light microscopy, Sarcocystis sporocysts have been observed in 14 out of the 20 (70.0%) European pine martens in our study. Due to the limited data on Sarcocystis infection rates in mustelids, the only comparable data come from studies conducted in Lithuania, which have reported slightly lower infection rates: 47.6% [49], 53.0% [48], and 55.9% [46]. In other predatory mammals, infection rates range from 38.0% for red foxes [83] to 66.7% for raccoon dogs [68] and 39.4% for Pampas foxes [84].
Molecular techniques play a crucial role in identifying Sarcocystis species in DHs. A variety of genetic markers are used for precise Sarcocystis species differentiation. However, the genetic characterization of many Sarcocystis species remains limited, making it challenging to design species-specific primer pairs. For instance, this limitation is evident with Sarcocystis linearis and Sarcocystis taeniata, both of which employ cervids as IHs [85]. Thus, further molecular studies are necessary to expand the genetic database of Sarcocystis spp. Based on previous transmission experiments, it was confirmed that members of the Mustelidae family can serve as DHs for various Sarcocystis species with rodents as their IHs, including Sarcocystis campestris (IH: Sciuridae), Sarcocystis citellivulpes (IH: Sciuridae), Sarcocystis muris (IH: Muridae), Sarcocystis putorii (IH: Cricetidae), and Sarcocystis undulati (IH: Sciuridae) [22,86]. However, the actual diversity of Sarcocystis species within the Mustelidae family is likely much greater according to recent molecular reports [46,47,48,49], highlighting the need for further investigations. Furthermore, some of the sequence data available in GenBank may be unreliable due to potential misidentification or lack of morphological validation [87,88]. Therefore, caution is warranted when interpreting the results of genetic analyses based on such data.
4.2. Importance of European Pine Marten in Transmission of Zoonotic and Potentially Pathogenic Sarcocystis Species
The role of Sarcocystis spp. in zoonotic transmission is increasingly recognized; however, many aspects remain poorly understood. Historically, humans were considered the DH for only two Sarcocystis species, S. hominis and S. suihominis [22]. In 2015, a novel zoonotic species, S. heydorni, was described, though data on its geographical distribution remain scarce [89]. More recently, in 2024, S. sigmoideus was characterized, with its zoonotic potential demonstrated in 2025 [90,91]. Humans can acquire infection through the consumption of raw or undercooked beef containing sarcocysts of S. hominis, S. heydorni, and S. sigmoideus or pork/wild boar meat contaminated with S. suihominis [30,31,89,90]. In this study, no zoonotic Sarcocystis species were identified in the intestinal samples of European pine martens collected in Latvia. Testing for S. hominis and S. suihominis was performed following a prior report of S. hominis DNA detected in the intestines of a single Lithuanian pine marten [49]. These findings suggest that mustelids may act as accidental hosts of zoonotic Sarcocystis, likely acquiring the parasite through scavenging infected cattle meat or exposure to contaminated environments rather than serving as true reservoirs.
During this study, multiple Sarcocystis species were identified, including S. bertrami, S. arieticanis, S. capracanis, and S. cruzi, all of which are known to cause acute infections in farm animals. The aforementioned Sarcocystis species have long been thought to be distributed only by canine predators, but recent studies show that mustelids also contribute to the distribution of these species ([40,41,42,43], PS). However, data on the pathogenicity of Sarcocystis species infecting cervids remain scarce. While S. hjorti has been suggested as a causative agent of eosinophilic fasciitis in red deer [45], the broader impact of these parasites on their IHs is yet to be determined.
In parallel, increasing reports of Sarcocystis-related foodborne illnesses have raised concerns about their potential risk to human health. In Japan, S. truncata was implicated in a food poisoning outbreak linked to venison consumption [33,92,93]. The reported cases exhibited symptoms such as nausea, vomiting, and diarrhea within 24 h of ingestion, consistent with other known foodborne pathogens. However, the extent to which these species contribute to human poisoning remains unclear.
4.3. Composition of Sarcocystis Species in European Pine Marten
This is first study on the identification of Sarcocystis species in the intestines of predators in Latvia. A summarized overview of Sarcocystis species, along with their respective IHs and DHs identified by molecular methods in the intestines of the European pine marten and mustelids in general, is presented in Table 3. Notably, S. entzeorthi, S. hjorti, S. truncata, and Sarcocystis sp. 25MmLV have been identified in European pine marten for the first time worldwide. Additionally, this study provides the first molecular confirmation that a member of the Mustelidae family can serve as the natural DH of S. hjorti. However, in previous studies, S. hjorti was not screened as a potential species transmitted through mustelids using species-specific primers [46,47,48,49].
Sarcocystis species using cattle, sheep, goats, and horses as their IHs and canids as their DHs (S. cruzi, S. arieticanis, S. capracanis, S. bertrami) were previously confirmed in the intestines of the European pine marten in Lithuania [46,49] and in the present study, indicating that these Sarcocystis spp. can use both canids and mustelids for their natural transmission. Similarly, some of the examined Sarcocystis species (S. hjorti, S. linearis, and S. morae), originally characterized by a Cervidae–Canidae (IH-DH) life cycle, can also be transmitted by mustelids ([49], PS). By contrast, S. miescheriana, the type species of the genus Sarcocystis infecting pigs/wild boar [22], was not detected in the small intestines of the European pine marten. Phylogenetic analyses revealed that Sarcocystis species utilizing cervids and bovids as IHs and canids as DHs cluster into a single group. In contrast, species that infect equids and suids as IHs and canids as DHs form distinct phylogenetic lineages [94]. Thus, we propose the hypothesis that alternative transmission via mustelids is a relatively common occurrence within the phylogenetic cluster comprising Sarcocystis species that utilize cervids and bovids as IHs and canids as DHs.
Based on phylogenetic results, the eight Sarcocystis species described in Eurasia and using cervids as IHs—Sarcocystis elongata, S. entzerothi, Sarcocystis japonica, S. matsuoae, Sarcocystis rangiferi, S. silva, Sarcocystis tarandi, and S. truncata—are closely related and do not group with other Sarcocystis spp. using cervids and canids as their IHs and DHs, respectively [95,96]. Of these species, S. elongata, S. entzerothi, S. japonica, S. silva, and S. truncata were confirmed in the intestines of American minks from Lithuania [47]. In this study, two of the aforementioned Sarcocystis species (S. entzerothi and S. truncata) were also identified in European pine martens from Latvia, further supporting the hypothesis that S. elongata, S. entzerothi, S. japonica, S. matsuoae, S. rangiferi, S. silva, S. tarandi, and S. truncata are naturally transmitted by various mustelid species.
In the current study, we did not identify any Sarcocystis species characterized by a Bovidae–Felidae (IH-DH) life cycle. This is despite the occasional detection of S. hirsuta and the fact that the prevalence of S. bovifelis in mustelids from Lithuania, a neighboring country to Latvia, has been reported to be as high as 89.0% [46,49]. Further research is required to elucidate this phenomenon, with potential explanations including variations in the prevalence of S. bovifelis across different years or the relatively uncommon presence of this species in Latvia. In general, studies on Sarcocystis species in Latvia are limited and do not typically examine cattle as the IHs or predators as the DHs of these parasites [97]. In summary, European pine martens and other mustelid species play a significant role in the transmission of Sarcocystis species that use Bovidae, Cervidae, and Equidae as their IHs. However, many questions remain unanswered regarding which predators—canids or mustelids (or felids or mustelids in the case of S. bovifelis)—contribute more to the transmission of species such as S. arieticanis, S. bertrami, S. capracanis, S. cruzi, S. hjorti, and S. morae. It is important to note that previous research has primarily focused on the feces or intestines of one or a few predatory mammals within a single family [45,68,83]. Additionally, it should be considered that, in general, Sarcocystis species transmitted by canids cannot be transmitted by felids, and vice versa [22]. Therefore, future studies should focus on investigating predatory mammals from the Canidae and Mustelidae families, as well as from the Felidae and Mustelidae families, as potential DHs of the Sarcocystis species discussed herein, within a single, comprehensive study.
Using in silico primers designed to amplify Sarcocystis spp. infecting birds and transmittable via placental predatory mammals (S. anasi, S. albifronsi, S. rileyi, and S. wenzeli) [48], we identified a genetically novel Sarcocystis species closely related to S. cristata and S. wenzeli in single European pine marten specimen (Figure 2). Our phylogenetic analysis revealed that the Sarcocystis species named as S. wenzeli, isolated from the feces of a gray wolf (PQ192598) [81], does not cluster with other S. wenzeli isolates and may represent a distinct species. Additionally, S. rileyi and Sarcocystis sp. 25MmLV, which phylogenetic results suggest is most likely S. wenzeli, were detected in the intestines of 11.0% of 115 Lithuanian mustelids [48]. Sarcocystis wenzeli is pathogenic to chickens [43,44], while S. rileyi forms macroscopic sarcocysts resembling rice grains in the muscles of ducks, leading to economic losses due to contamination of hunted duck meat [48,59,98]. Thus, mustelids play a significant role in the transmission of important Sarcocystis species that infect birds consumed as human food.
5. Conclusions
In the present study, nine Sarcocystis species, S. arieticanis, S. bertrami, S. capracanis, S. cruzi, S. entzerothi, S. hjorti, S. morae, S. truncata, and Sarcocystis sp. 25MmLV, were identified in the intestinal samples of European pine martens from Latvia using molecular analysis. Some of the detected Sarcocystis species can be pathogenic to livestock or cervids. In addition, S. truncata has been associated with food poisoning due to the consumption of venison. Our study provides evidence that the European pine marten plays an important role as a DH for Sarcocystis species, using cervids and livestock as IHs. Moreover, further research is necessary to elucidate the ecological role of mustelids in the transmission dynamics of Sarcocystis spp.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Kollias G.V. Fernandez-Moran J. Mustelidae Fowler’s Zoo and Wild Animal Medicine Elsevier Amsterdam, The Netherlands 2015 Volume 8476491978-1-4557-7397-8
- 2Yonezawa T. Nikaido M. Kohno N. Fukumoto Y. Okada N. Hasegawa M. Molecular Phylogenetic Study on the Origin and Evolution of Mustelidae Gene 200739611210.1016/j.gene.2006.12.04017449200 · doi ↗ · pubmed ↗
- 3Sato J.J. Hosoda T. Wolsan M. Tsuchiya K. Yamamoto M. Suzuki H. Phylogenetic Relationships and Divergence Times among Mustelids (Mammalia: Carnivora) Based on Nucleotide Sequences of the Nuclear Interphotoreceptor Retinoid Binding Protein and Mitochondrial Cytochrome b Genes Zool. Sci.20032024326410.2108/zsj.20.24312655187 · doi ↗ · pubmed ↗
- 4Wright P.G.R. Croose E. Macpherson J.L. A Global Review of the Conservation Threats and Status of Mustelids Mamm. Rev.20225241042410.1111/mam.12288 · doi ↗
- 5OzoliņšJ. Pilāts V. Distribution and Status of Small and Medium-Sized Carnivores in Latvia Ann. Zool. Fenn.1995322129
- 6Helldin J. Population Trends and Harvest Management of Pine Marten Martes martes in Scandinavia Wildl. Biol.2000611112010.2981/wlb.2000.006 · doi ↗
- 7Bolduc D. Fauteux D. Gauthier G. Legagneux P. Seasonal Role of a Specialist Predator in Rodent Cycles: Ermine–Lemming Interactions in the High Arctic Ecology 2025106 e 451210.1002/ecy.451239838535 PMC 11751380 · doi ↗ · pubmed ↗
- 8Central Statistical Bureau of Latvia Number of Game Animals and Game Animals [Dataset]Available online: https://data.stat.gov.lv/pxweb/en/OSP_PUB/START__ENV__DR__DRM/DRM 010(accessed on 21 March 2025)