Mechanistic Insights into Drug-Induced Guillain–Barré Syndrome: A Large-Cohort Analysis of the FAERS Database
Jianxiong Gui, Xiao Li, Hongyuan Chu, Junjiao Zhang, Meiyu Dong, Fan Zhang, Renqiuguo Li, Huaxia Luo, Kai Gao, Yuwu Jiang

TL;DR
This study uses a large database to identify drugs and biological pathways linked to drug-induced Guillain–Barré Syndrome, offering insights into its mechanisms.
Contribution
The study provides novel mechanistic insights by linking drug targets to GBS susceptibility proteins using GWAS, MR, and PPI network analysis.
Findings
Nelarabine showed the strongest correlation with drug-induced GBS.
TNF and PDCD1LG2 were identified as key GBS-susceptible genes.
Pyrimidine metabolism and IL6/JAK/STAT3 pathways contribute to drug-induced GBS.
Abstract
Background/Objectives: Drug-induced Guillain–Barré Syndrome (GBS) is a severe complication of pharmacotherapy. Previous research has established a connection between certain medications and higher GBS risk. However, a large-cohort analysis is crucial to reveal underlying biological mechanisms of drug-induced GBS. This study aimed to evaluate the association between GBS and various drugs currently accessible in the Food and Drug Administration Adverse Event Reporting System (FAERS) database and explore the mechanisms underlying drug-induced GBS. Methods: We analyzed drug-induced GBS adverse event reports in the FAERS database to identify strongly associated drugs. We then investigated GBS susceptibility proteins through GWAS meta-analysis and Mendelian Randomization (MR) based on plasma proteomics, complemented by protein–protein interaction (PPI) network analysis to explore underlying…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
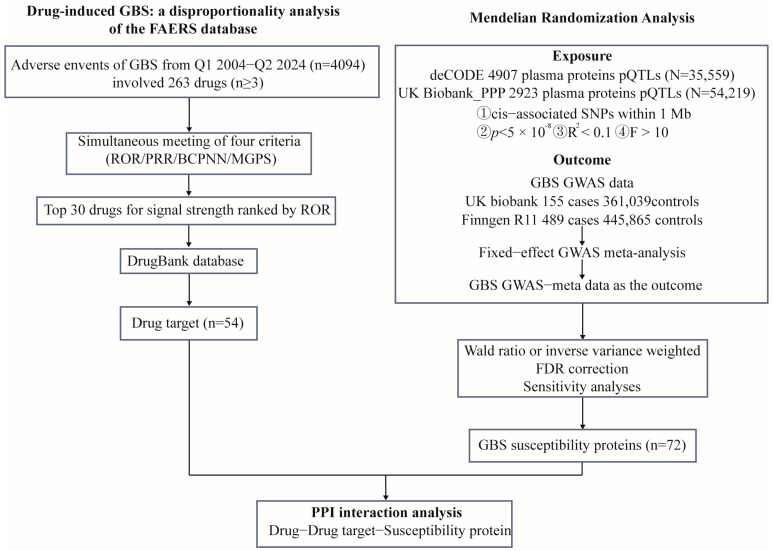 Figure 1
Figure 1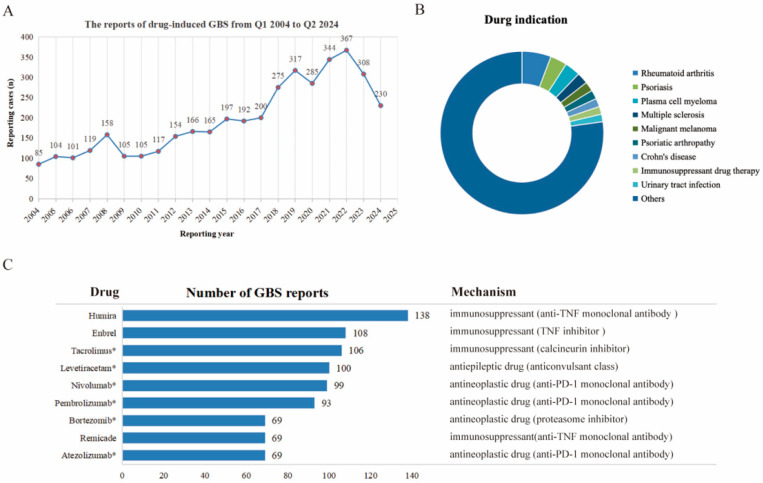 Figure 2
Figure 2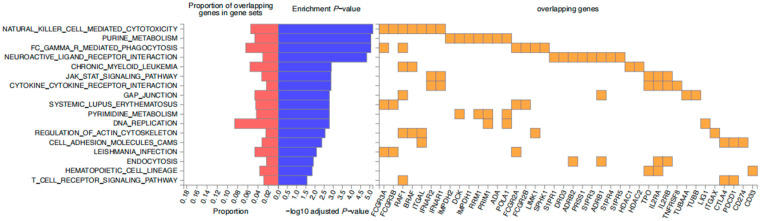 Figure 3
Figure 3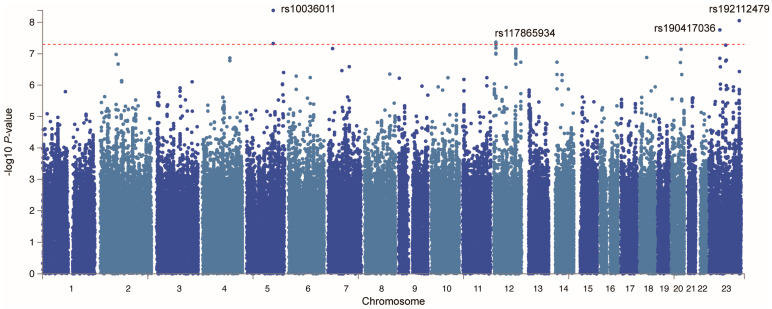 Figure 4
Figure 4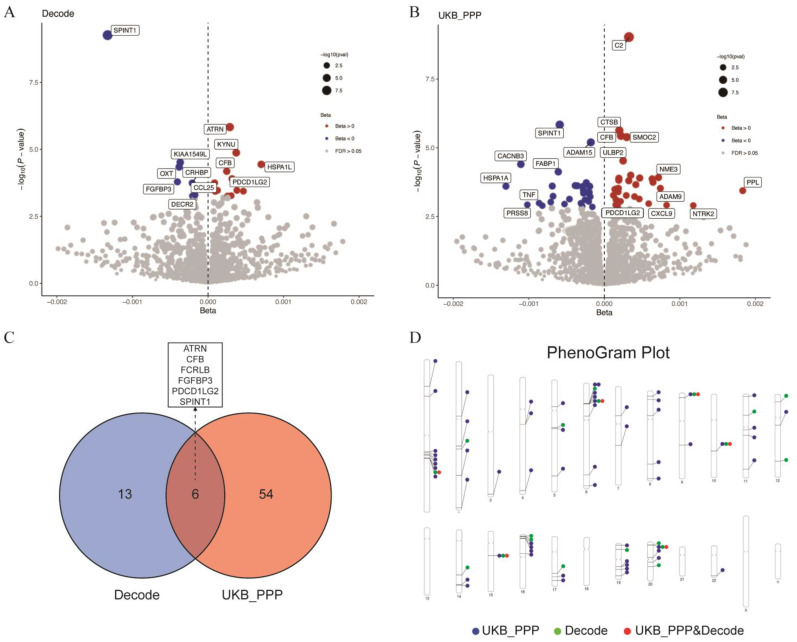 Figure 5
Figure 5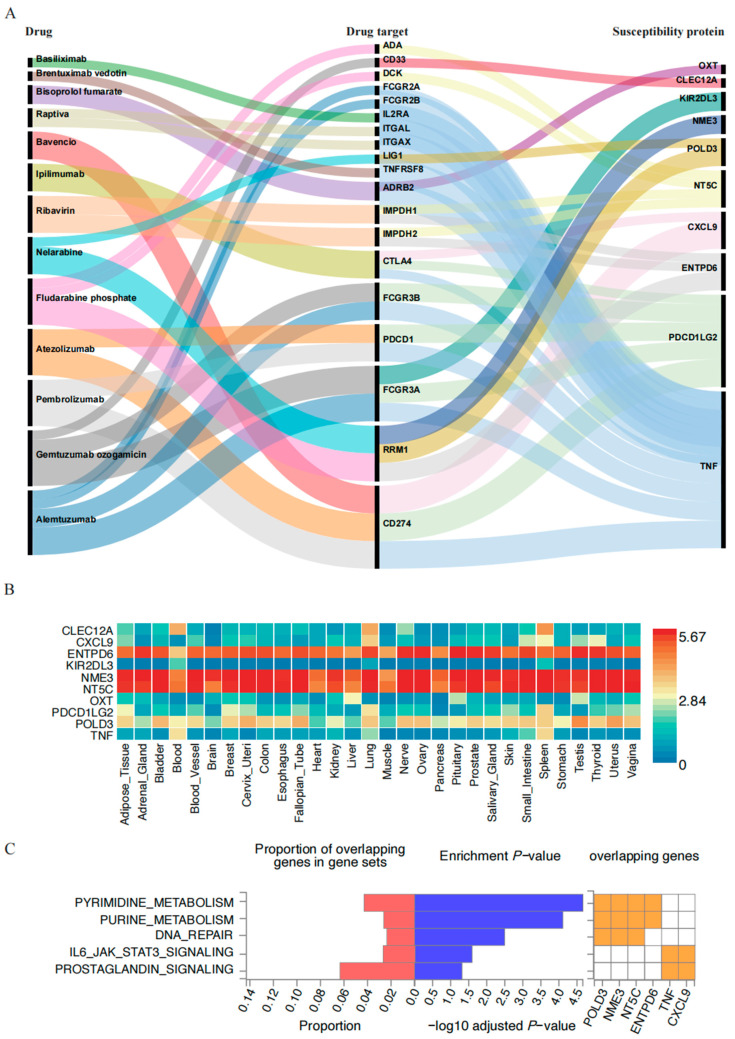 Figure 6
Figure 6- —National Natural Science Foundation of China
- —Fundamental Research Funds for the Central Universities
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPeripheral Neuropathies and Disorders · Computational Drug Discovery Methods · Chronic Lymphocytic Leukemia Research
1. Introduction
Guillain–Barré Syndrome (GBS) is an acute inflammatory polyradiculoneuropathy affecting the peripheral nervous system, marked by progressive limb weakness, sensory deficits, cranial nerve involvement, and cerebrospinal fluid (CSF) albuminocytological dissociation. It is the leading cause of acute flaccid paralysis globally [1]. The worldwide incidence of GBS ranges from 0.16 to 3.0 cases per 100,000 person-years [2,3]. Drug-induced GBS is a life-threatening condition initially identified in individuals who received the influenza vaccine [4]. Associated drugs include allopurinol, gold therapy, gangliosides, TNF-alpha antagonists, second-generation antipsychotics like risperidone, and some antibiotics [5,6,7,8]. Most reports on drug-induced GBS are sporadic, with only a few medications occasionally documented in literature reviews and summaries regarding their potential to trigger GBS. For instance, it has been reported that the frequency of demyelinating diseases in patients receiving anti-TNF-α therapy ranges from approximately 0.02% to 0.2% [9]. Moreover, a study on neuroimmunological adverse events associated with immune checkpoint inhibitors (ICIs) reported that the proportion of GBS in patients treated with ICIs was nearly eight times higher than in the control group [10]. GBS with potentially life-threatening consequences occurs in approximately 0.1–0.2% of patients treated with ICIs [11].Currently, the specific details and underlying mechanisms are unclear, necessitating a comprehensive analysis of large patient samples to elucidate the characteristics of these commonly used drugs and mechanisms in drug-induced GBS.
The Food and Drug Administration Adverse Event Reporting System (FAERS) is a voluntary system that collects information on drug-related adverse events reported by healthcare professionals, patients, and manufacturers. It provides valuable insights into drug safety and serves as an essential resource for monitoring adverse reactions to pharmaceuticals [12]. However, this database has not yet been fully utilized to investigate drug-induced GBS. To explore the underlying mechanisms and gain a comprehensive understanding of drug-induced GBS, we analyzed all reports of drug-induced GBS currently available in the FAERS database. Mendelian Randomization (MR) and genome-wide association study (GWAS) meta-analysis based on plasma proteomics were employed for drug target identification and the identification of GBS susceptibility proteins. To further investigate the association between drug use and GBS, protein–protein interaction (PPI) network analysis was also utilized. By integrating GWAS, MR, and PPI network analysis, this study aimed to improve our understanding of how specific drugs may trigger this serious condition.
2. Results
The schematic presentation of the overall study design is shown in Figure 1.
2.1. Descriptive Analysis
Clinical characteristics of reports with drug-induced GBS from the FAERS database (2004 Q1–2024 Q2).
After the removal of duplicates and subsequent filtering, a total of 4094 reports of drug-induced GBS were identified. The clinical characteristics associated with these reports from the FAERS database were presented in Table 1. Notably, the proportion of males (49.98%) was significantly higher than that of females (38.86%). Regarding age distribution, although the ages of 27.58% of patients remained unknown, most reported cases occurred within the middle-aged and elderly demographic (41–64 years old), which accounted for 31.31%. The incidence rate of drug-induced GBS among pediatric patients (≤18 years old) was 3.86%. Among all reported data, the United States represented the largest share at 24.94%. In terms of clear indications, most patients (82.49%) had well-documented records indicating specific drugs responsible for causing GBS.
Further analysis of drugs associated with GBS revealed their primary uses in treating conditions such as rheumatoid arthritis, psoriasis, plasma cell myeloma, multiple sclerosis, malignant melanoma, psoriatic arthropathy, Crohn’s disease, immunosuppressant therapy, and urinary tract infections (Figure 2B). Figure 2C presented the drugs with the highest number of reported cases, ranked as follows: Humira (n = 138), Enbrel (n = 108), Tacrolimus (n = 106), Levetiracetam (n = 100), Nivolumab (n = 99), Pembrolizumab (n = 93), Bortezomib (n = 69), Remicade (n = 69), and Atezolizumab (n = 69). Apart from Levetiracetam, all the listed drugs are classified as either immunosuppressants or antineoplastic agents.
2.2. Signal Strength Detection
If we consider only the number of cases, it would not provide a comprehensive view. A higher number of reports linking a particular drug to drug-induced GBS might simply reflect its widespread use, rather than indicating the strongest association. Therefore, we employed four well-established drug association analysis formulas to identify the drugs with the strongest correlation to GBS. Table 2 presents the top 30 drugs with the highest signal strength, ranked by ROR. The top five drugs were Nelarabine (n = 16, ROR 138.91, PRR 134.71, IC 7.07, EBGM 134.19), Roferon-a (n = 3, ROR 60.62, PRR 59.81, IC 5.90, EBGM 59.77), Zerit (n = 3, ROR 30.17, PRR 29.97, IC 4.90, EBGM 29.95), Methimazole (n = 9, ROR 22.43, PRR 22.32, IC 4.48, EBGM 22.28), and Pravastatin sodium (n = 4, ROR 22.25; PRR 22.15; IC 4.47; EBGM 22.13).
Moreover, the drug targets and mechanism for these medications are presented in Supplementary Table S1, based on the Drug Bank database. To further elucidate the relevance between the drug targets of medicines inducing GBS, KEGG (Kyoto Encyclopedia of Genes and Genomes) pathway analysis was performed. This analysis identified several pivotal pathways, with the top five being natural killer cell-mediated cytotoxicity, purine metabolism, Fc gamma R-mediated phagocytosis, neuroactive ligand-receptor interaction, and the chronic myeloid leukemia signaling pathway (Figure 3).
2.3. Potential Susceptibility Proteins for GBS
To identify potential GBS susceptibility proteins, we first conducted a GWAS meta-analysis by integrating GBS GWAS data. The QQ plot derived from the GWAS meta-analysis is presented in Figure S1, revealing a genomic inflation factor (λ) of 1.015. Typically, a value greater than 1 indicates acceptable stability, providing reliable outcome data for subsequent MR analyses. Our investigation revealed four significant SNP loci (Figure 4, Table S2) with p-values less than 5 × 10^−8^, as depicted in the Manhattan Plot of GWAS meta-analysis, suggesting a statistically significant association with a specific trait or disease. However, as shown in the Manhattan plot based on genetic analysis, we were unable to directly identify genetic risk loci significantly associated with GBS (p < 5 × 10^−8^), which could be attributed to the relatively small sample size of GBS cases (Figure S2).
Subsequently, MR analysis was conducted using plasma proteomic data as exposure and GBS GWAS-meta data as the outcome. In the Decode dataset, 20 proteins associated with GBS risk were identified (Table S3a), while 61 proteins were identified in the UKB_PPP dataset (Table S3b). Following sensitivity and heterogeneity analyses, a total of 73 GBS susceptibility proteins were included, comprising 19 from Decode and 60 from UKB_PPP (Figure 5A,B), with six proteins shared between the two datasets: attractin (ATRN), complement factor B (CFB), Fc receptor-like B (FCRLB), fibroblast growth factor binding protein 3 (FGFBP3), programmed cell death 1 ligand 2 (PDCD1LG2), and serine peptidase inhibitor, Kunitz type 1 (SPINT1) (Figure 5C). Reverse MR analysis, which used GBS GWAS-meta data as the exposure and plasma proteomic data as the outcome, suggested no reverse causal relationship between GBS and the 73 proteins (Table S3c). This finding suggested that the potential reverse causal effects of GBS on these proteins had been thoroughly considered and excluded. Consequently, this reinforced the hypothesis that these proteins were likely involved in susceptibility to GBS. The chromosomal distribution of these genes encoding 73 proteins is illustrated in Figure 5D.
2.4. PPI Network and Enrichment Analysis Results
Additionally, we generated PPI networks to explore the interactions between the 73 GBS susceptibility proteins and previously identified drug targets (Figure S3, Table S4). The PPI network provided a detailed visualization of the potential interactions between drugs and their putative targets, as well as the connections between genes encoding GBS-related proteins and these drug targets. This comprehensive illustration enhanced our understanding of the intricate relationships within the network, highlighting potential avenues for therapeutic intervention. A total of 10 genes (out of 73) encoding GBS-susceptible proteins were found to interact with 19 genes (out of 54) encoding drug targets that were potentially associated with GBS. These 19 drug targets were involved in a total of 13 different drugs (Figure 6A). The top five GBS susceptibility genes with the most association with drug targets were the following: Tumor Necrosis Factor (TNF, 15 interactions), Integrin Subunit Alpha X (ITGAX, nine interactions), Fc gamma receptor IIIa (FCGR3A, six interactions), 5′, 3′-nucleotidase, cytosolic (NT5C, five interactions), and PDCD1LG2 (five interactions). By analyzing PPI networks, we identified key genes associated with drug targets. The GBS susceptibility-related gene TNF had the highest number of interactions, with 15 drug targets. The drugs associated with these targets included Bisoprolol fumarate (cardiovascular drug), Atezolizumab (antineoplastic drug), Alemtuzumab (immunosuppressant), Bavencio (antineoplastic drug), Pembrolizumab (antineoplastic drug), Ipilimumab (antineoplastic drug), Basiliximab (immunosuppressant), and Raptiva (immunosuppressant), with Alemtuzumab (immunosuppressant) being the most closely related. PDCD1LG2 was the second most interactive protein, binding to five drug targets, with Gemtuzumab ozogamicin (antineoplastic drug) being the most closely associated drug.
Subsequently, we performed an organ system distribution analysis of these 10 GBS-related proteins. Figure 6B illustrated the expression levels of specific proteins in different tissues, aiding in understanding their functions and distributions across various tissues. Notably, TNF, DNA polymerase delta 3 (POLD3), PDCD1LG2, Killer cell immunoglobulin like receptor, two Ig domains and long cytoplasmic tail 3 (KIR2DL3), Ectonucleoside triphosphate diphosphohydrolase 6 (ENTPD6), C-X-C motif chemokine ligand 9 (CXCL9), and C-type lectin domain family 12 member A (CLEC12A) exhibited high expression in immune-related tissues (such as blood and spleen), suggesting their potential involvement in immune responses and inflammatory processes.
To explore the possible biological mechanisms of GBS-related genes identified by MR, we conducted enrichment analysis on these 10 genes, filtering the results to include only pathways with a corrected p < 0.05 for significance. We identified a total of five pathways associated with GBS (Figure 6C). These genes were primarily involved in the following processes: pyrimidine metabolism, purine metabolism, DNA repair, IL6/JAK/STAT3 signaling, and prostaglandin signaling.
3. Discussion
GBS is an immune-mediated polyneuropathy characterized by areflexia and ascending paresthesia, which can progress to respiratory failure [1]. The use of certain drugs, particularly some ICIs employed in immunotherapy for melanoma and refractory cancers, may elevate the risk of developing GBS [11,13]. However, the possible mechanisms and triggers of GBS in patients receiving specific medications remain unknown. This study focused on occurrences of GBS associated with drugs use as recorded in the FAERS database and explored potential mechanisms through MR analysis and PPI analysis.
In this study, the number of reported cases of drug-induced GBS has increased steadily from 2019 to 2022, reaching its peak in 2022. Recent reports found that the COVID-19 virus and its vaccines can increase the incidence of GBS by two to five times, leading to more severe cases of GBS in patients [14,15]. Furthermore, the first report of GBS caused by COVID-19 and its vaccines were in 2019 and 2021, respectively [15,16]. Therefore, we speculate that the increased incidence of drug-induced GBS during this period may be related to the immune system disorders caused by COVID-19 and its vaccines [16].
Our findings indicated that drug-induced GBS was predominantly associated with medications utilized in the treatment of immune system disorders and cancers. Among the 13 drugs associated with the 10 GBS-susceptible genes, eight were antineoplastic drugs, three were immunosuppressant drugs, one was a cardiovascular drug, and one was an antiviral drug. Among these antineoplastic drugs, Nelarabine exhibited the strongest correlation with GBS. Nelarabine is a purine nucleoside analog used in the management of T-cell acute lymphoblastic leukemia (T-ALL) and T-cell lymphoblastic lymphoma (T-LBL) [17]. Its primary mechanism involves disrupting DNA synthesis by inhibiting Ribonucleotide Reductase M1 (RRM1), thereby hindering the proliferation of cancer cells. Additionally, Nelarabine may induce apoptosis in cells with compromised LIG1 function [18]. This study validated that the drug targets of Nelarabine were RRM1 and LIG1 (DNA ligase 1); however, the mechanisms underlying its side effects that lead to GBS remain unclear. PPI analysis in this study suggested that Nelarabine-induced GBS may be mediated via the RRM1/LIG1-POLD3 or RRM1-NME3 pathway. POLD3 is essential for DNA repair and maintaining immune cell function [19]. Previous studies have indicated that LIG1 level can influence the expression of POLD3 [20,21]. Additionally, NME3 (Nucleoside Metabolism Enzymes 3) plays a significant role in repairing both single- and double-stranded breaks in DNA, contributing to genomic instability and promoting malignant tumor progression [22,23]. This study proposed that the drug target RRM1 may induce GBS through POLD3 and NME3 proteins. However, the biological significance of the results requires further experimental verification.
The three immunosuppressant drugs Raptiva, Basiliximab and Alemtuzumab were all linked to the TNF gene, which opens new avenues for understanding and treating this debilitating condition. These medications are widely utilized in the treatment of various autoimmune and inflammatory diseases. Several studies indicate that TNF-α produced by T cells plays a crucial pro-inflammatory role in GBS, contributing to demyelination and axonal damage in peripheral nerves [24]. In 1993, one study documented that elevated serum concentrations of TNF-α are detectable in 20% to 50% of patients diagnosed with GBS [25]. The release of TNF-α may exacerbate inflammatory demyelination [26]. However, the MR analysis in Figure 6B shows a negative association between TNF expression and drug-induced GBS. Anti-TNF-α drugs have been linked to an increased incidence and activity of demyelinating diseases like GBS [27]. This paradox may be explained by the dual role of TNF-α: while it promotes inflammation, it also has immunoregulatory functions, suppressing T cell reactivity to autoantigens. Systemic anti-TNF-α therapy could penetrate the peripheral nervous system where the blood–nerve barrier is weak, potentially neutralizing local TNF-α and disrupting its balance [28]. Reduced TNF-α levels could prolong the activation of myelin-specific T cells, raising the risk of immune-mediated neuropathy in susceptible individuals [29]. These findings emphasized that patients undergoing anti-TNF-α therapy should be closely monitored for neurological symptoms indicative of GBS. Additionally, if patients experience GBS-related adverse reactions while taking medications potentially associated with TNF-α in Figure 6A, the possibility of persistently activated myelin-specific T cells due to excessively low TNF-α levels should be considered. Therefore, when evaluating and treating such patients, clinicians should pay attention to the immunomodulatory role of TNF-α and its impact on the nervous system to develop more precise treatment strategies.
Additionally, PDCD1LG2 (PD-L2) emerged as another significant protein that was positively associated with drug-induced GBS, as indicated in Figure 5A,B. The PPI analysis carried out in this study presented potential associations between PDCD1LG2 and the drug targets of Ipilimumab, Gemtuzumab ozogamicin, Atezolizumab, Pembrolizumab, Bavencio and Alemtuzumab. These drugs are all monoclonal antibodies used in cancer treatment [30,31,32,33]. However, there have been no previous reports linking GBS to PDCD1LG2 protein. PDCD1LG2 is an important immunoregulatory molecule primarily expressed in antigen-presenting cells (APCs) [34]. It regulates T cell activation and suppresses immune responses through its interaction with the receptor PD-1 [35]. This study suggests that caution is warranted regarding the occurrence of GBS when using the aforementioned monoclonal antibodies. PDCD1LG2 inhibitors may serve as a novel therapeutic target for preventing drug-induced GBS. Furthermore, investigating the mechanism of PDCD1LG2 in GBS could aid in enhancing the safety of immunotherapy.
Unexpectedly, the use of the cardiovascular drug Bisoprolol fumarate was also associated with the occurrence of GBS, a finding that had never been reported in the literature before. This study revealed that the β2 adrenergic receptor (ADRB2) was the drug target for Bisoprolol fumarate. ADRB2 is a G protein-coupled transmembrane receptor that is widely recognized as a pharmacological target for the treatment of asthma and chronic obstructive pulmonary disorder (COPD) [36]. Previous literature indicated that overactivation of ADRB2 can lead to a decrease in TNF-α levels [36]. Therefore, if neurological symptoms resembling GBS arise during the use of such medications, this mechanism could be taken into consideration.
To explore additional mechanisms, we conducted KEGG pathway analyses on genes encoding drug targets associated with GBS as well as 10 GBS susceptibility-related genes. The results revealed three common pathways: purine metabolism, IL6-JAK-STAT3 signaling, and pyrimidine metabolism. Previous studies suggest that metabolic factors may also play a role in the onset and progression of the disease [37,38]. Patients with metabolic syndrome exhibiting abnormalities in purine metabolism has been associated with an increased risk of developing GBS. Certain metabolic indicators (such as uric acid levels) in GBS patients may correlate with disease severity or prognosis [37,39]. Additionally, certain drugs that promote pyrimidine degradation may aid in GBS treatment [40]. To clarify this phenomenon, we conducted KEGG and PPI network analyses to identify GBS-related proteins involved in purine and pyrimidine metabolic pathways. We identified four such proteins: POLD3, NME3, NT5C, and ENTPD6. Through drug target analysis, these proteins were associated with three medications—Ribavirin, Nelarabine, and Fludarabine phosphate—all of which are purine or pyrimidine nucleoside analogs [41,42]. The potential mechanism underlying their action may involve the activation and function of immune cells being dependent on metabolic pathways, including purine and pyrimidine metabolism [43]. Metabolic dysregulation could impact immune cell activation and inflammatory responses. This finding offers new insights for clinical treatment strategies.
4. Materials and Methods
4.1. Data Sources and Processing
The data for this study were obtained from drug-induced GBS data in the FAERS database (https://open.fda.gov/data/faers/) (accessed on 28 September 2024) from Q1 2004 to Q2 2024. The reporting odds ratio (ROR) [44], proportional reporting ratio (PRR) [45], Bayesian confidence propagation neural network (BCPNN) [46], and multi-item gamma-Poisson shrinker (MGPS) [47] models were simultaneously applied to calculate signal values to yield more rigorous signals. Detailed criteria and specific calculations are listed in Table 3. The Medical Dictionary for Regulatory Activities (MedDRA) was used to identify drug-induced GBS cases, and the DrugBank database was used to determine drug generic names and drug targets. Additionally, the study explored the causal effect of gene variants linked to protein quantitative trait loci (pQTLs) on GBS risk using MR analysis. This study utilized gene variants linked to 4907 plasma proteins from the deCODE project, which included a sample size of 35,559 Icelanders [48], as well as gene variants related to 2923 plasma proteins from the UK Biobank_PPP project with a sample size of 54,219 participants [49], as exposure factors. Details on protein measurement, data processing, and quality control are available in the original studies.
We performed a fixed-effect GWAS meta-analysis using the METAL package as outcome factors [48,49], which integrated data from the UK Biobank (155 GBS cases and 361,039 controls) and FinnGen R11 (489 GBS cases and 445,865 controls). The quantile–quantile plot was generated using the ‘qqman’ package in R (version 4.3.0) [50]. Regional association plots for the lead SNP at each genome-wide significant locus were generated using the ‘gassocplot’ package in R (version 4.3.0) [51]. The results of the GWAS meta-analysis were annotated using FUMA (https://fuma.ctglab.nl/) (accessed on 4 October 2024) [52].
4.2. Mendelian Randomization
To fulfill the fundamental assumptions of MR—namely relevance, independence, and exclusion restriction—it is imperative to ensure the robustness of MR findings. The relevance assumption stipulates that IVs must exhibit a strong association with exposure. Independence necessitates that IVs are uncorrelated with confounding factors that could distort the exposure–outcome relationship. The exclusion restriction assumption asserts that IVs affect the outcome solely through their influence on the exposure, without any alternative pathways. The criteria for selecting pQTLs were as follows [53]: (1) cis-associated SNPs within 1 Mb of the gene encoding the target protein; (2) genome-wide significance defined by p < 5 × 10^−8^; (3) no significant linkage disequilibrium (LD), indicated by R^2^ < 0.1 in the 1000 Genomes European population; and (4) an F-statistic greater than 10 for each protein’s pQTL.
The MR analysis mainly employed both Wald ratio and inverse variance weighted (IVW) methods to evaluate the causal effect of plasma protein-related gene variants on GBS risk. Depending on the number of SNPs utilized as IVs, either IVW or the Wald ratio method was employed for each protein, using the ’TwoSampleMR’ package in R (version 4.3.0). To ensure the robustness of our findings, sensitivity analyses were conducted using MR-Egger regression, weighted median, weighted mode, and simple mode to detect directional pleiotropy [49]. Horizontal pleiotropy was evaluated through the MR-Egger intercept test, and heterogeneity was assessed using Cochran’s Q-test in both IVW and MR-Egger analyses. The presence of horizontal pleiotropy and heterogeneity can be ruled out if the following conditions are satisfied: the p-value of the MR-Egger intercept is >0.05, the p-value of Cochran’s Q test is >0.05, and the effect estimates of IVW are directionally consistent with at least two other estimation methods. Odds ratios (ORs) and 95% confidence intervals (CIs) were applied to quantify the associations between proteins and outcomes. A 5% false discovery rate (FDR) correction was applied to address multiple comparisons to avoid Type I errors in finding genetic associations [54]. In the reverse MR analysis, GBS was considered the exposure (with IVs meeting the criteria p < 5 × 10^−8^, R^2^ < 0.1, and F-statistic > 10), while plasma proteins were treated as the outcomes. Reverse causality estimates were similarly adjusted for multiple testing using FDR correction across all tested protein traits.
4.3. PPI Network and Enrichment Analysis
To investigate the potential mechanisms underlying drug-induced GBS, PPI analysis was conducted to assess the interactions between GBS susceptibility proteins and drug targets using the STRING database (https://string-db.org, accessed on 24 March 2025) [55]. The average expression levels of these critical protein-coding genes were examined across 54 human tissues using GTEx v8 data and presented as a heatmap. Furthermore, gene enrichment analysis revealed potential pathways associated with these genes using the ‘GENE2FUNC’ tool in FUMA (https://fuma.ctglab.nl/, accessed on 24 March 2025) [52].
4.4. Limitations
We conducted an in-depth investigation into the possible mechanisms of drug-induced GBS through FAERS, Mendelian Randomization, and Protein Interaction Networks, but there are still some limitations. Firstly, FAERS is based on voluntary reporting, which may introduce bias and result in underreporting of adverse events (AEs), thereby affecting the accuracy of the data. Secondly, the limited number of cases available in GWAS related to GBS restricted our ability to identify genes associated with GBS risk. Furthermore, both GWAS and proteomic datasets predominantly originated from populations of European ancestry; thus, caution should be exercised when extrapolating findings regarding GBS susceptibility proteins to other demographic groups. Our exploration of potential mechanisms between susceptible proteins and drug targets was based on bioinformatics predictions that require validation through biological experiments or clinical cohort studies in the future.
5. Conclusions
Our results investigated the association between drug use and GBS by analyzing adverse event reports from the FAERS and employing MR and PPI network analysis to explore underlying mechanisms. The study identified specific drugs strongly associated with GBS, such as Nelarabine. The findings suggested TNF and PDCD1LG2 as potential therapeutic targets for drug-induced GBS, offering insights into both immune regulation and metabolic pathways involved in the disease. Our research provided possible biological mechanisms of drug-induced GBS, offering potential targets for therapeutic intervention and further research into the pathogenesis of GBS.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Wijdicks E.F.M. Klein C.J. Guillain-Barré Syndrome Mayo Clin. Proc.20179246747910.1016/j.mayocp.2016.12.00228259232 · doi ↗ · pubmed ↗
- 2Xu L. Zhao C. Bao Y. Liu Y. Liang Y. Wei J. Liu G. Wang J. Zhan S. Wang S. Variation in worldwide incidence of Guillain-Barré Syndrome: A population-based study in urban China and existing global evidence Front. Immunol.202415141598610.3389/fimmu.2024.141598639318625 PMC 11420027 · doi ↗ · pubmed ↗
- 3Kozyreva A.A. Bembeeva R.T. Druzhinina E.S. Zavadenko N.N. Guillain-Barre Syndrome in children Zh. Nevrol. Psikhiatr. Im. S. S. Korsakova 2023123203210.17116/jnevro 20231230922037942969 · doi ↗ · pubmed ↗
- 4Haber P. De Stefano F. Angulo F.J. Iskander J. Shadomy S.V. Weintraub E. Chen R.T. Guillain-Barré Syndrome following influenza vaccination JAMA 20042922478248110.1001/jama.292.20.247815562126 · doi ↗ · pubmed ↗
- 5Awong I.E. Dandurand K.R. Keeys C.A. Maung-Gyi F.A. Drug-associated Guillain-Barré Syndrome: A literature review Ann. Pharmacother.19963017318010.1177/1060028096030002128835052 · doi ↗ · pubmed ↗
- 6Fagius J. Osterman P.O. Sidén A. Wiholm B.E. Guillain-Barré Syndrome following zimeldine treatment J. Neurol. Neurosurg. Psychiatry 198548656910.1136/jnnp.48.1.653156214 PMC 1028185 · doi ↗ · pubmed ↗
- 7Raschetti R. Maggini M. Popoli P. Caffari B. Da Cas R. Menniti-Ippolito F. Spila-Alegiani S. Traversa G. Gangliosides and Guillain-Barré Syndrome J. Clin. Epidemiol.1995481399140510.1016/0895-4356(95)00557-97490603 · doi ↗ · pubmed ↗
- 8Okuyan E. Cakar M.A. Dinckal M.H. Guillain-Barré Syndrome after thrombolysis with streptokinase Cardiol. Res. Pract.2010201031585610.4061/2010/31585621234097 PMC 3014663 · doi ↗ · pubmed ↗