BcAS2 Regulates Leaf Adaxial Polarity Development in Non-Heading Chinese Cabbage by Directly Activating BcPHB Transcription
Cheng Jiang, Qiang Ding, Ying He, Yiran Li, Zhanyuan Gao, Entong Li, Xilin Hou

TL;DR
This study shows how the BcAS2 gene controls leaf shape in non-heading Chinese cabbage by activating another gene called BcPHB.
Contribution
The study identifies BcAS2 as a direct activator of BcPHB, revealing a new molecular pathway for leaf polarity regulation in non-heading Chinese cabbage.
Findings
Overexpression of BcAS2 causes upward leaf curling in non-heading Chinese cabbage.
BcAS2 directly binds to the BcPHB promoter and activates its transcription.
BcPHB is confirmed to regulate leaf adaxial polarity development.
Abstract
Leaves are the primary organs for plant photosynthesis, and their flat, symmetric morphology is crucial for plant growth and development. The LBD family transcription factor ASYMMETRIC LEAVES 2 (AS2) plays a central role in the establishment of leaf polarity. In this study, we cloned the BcAS2 gene from the non-heading Chinese cabbage cultivar “NHCC001” and successfully generated overexpression strains through genetic transformation. Phenotypic analysis revealed that overexpression of BcAS2 led to significant upward curling of leaves in non-heading Chinese cabbage. Additionally, we found that the expression of BcPHB, a gene associated with leaf adaxial polarity development, was significantly up-regulated in BcAS2-overexpressing plants compared to controls. This interaction was further confirmed through yeast one-hybridization (Y1H), dual-luciferase reporter assays, and electrophoretic…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
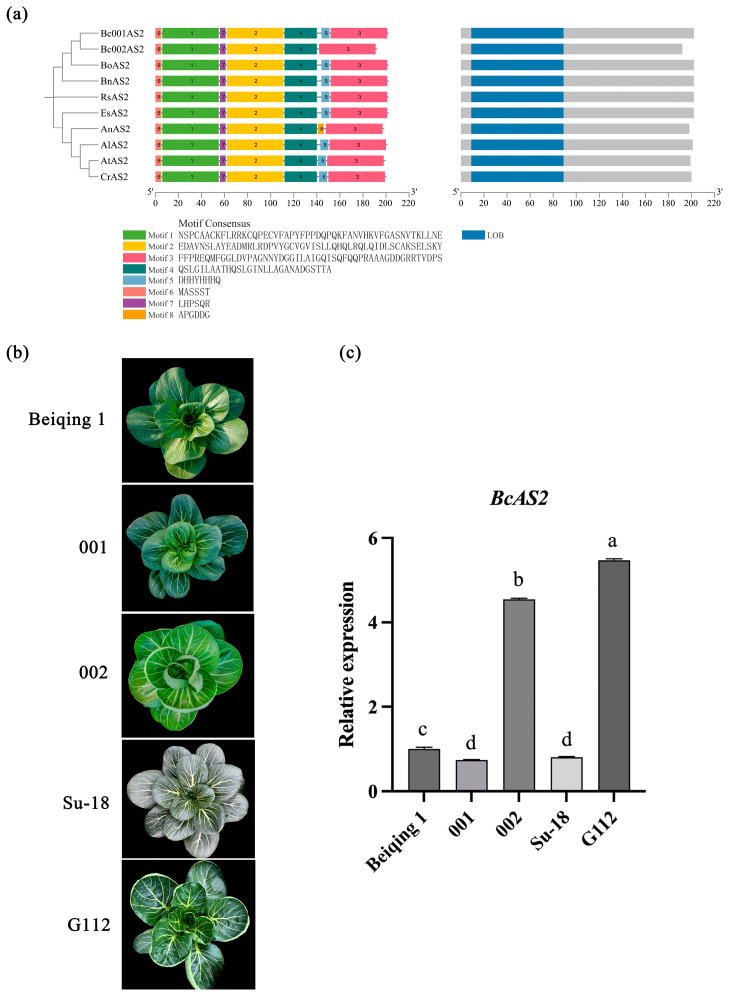 Figure 1
Figure 1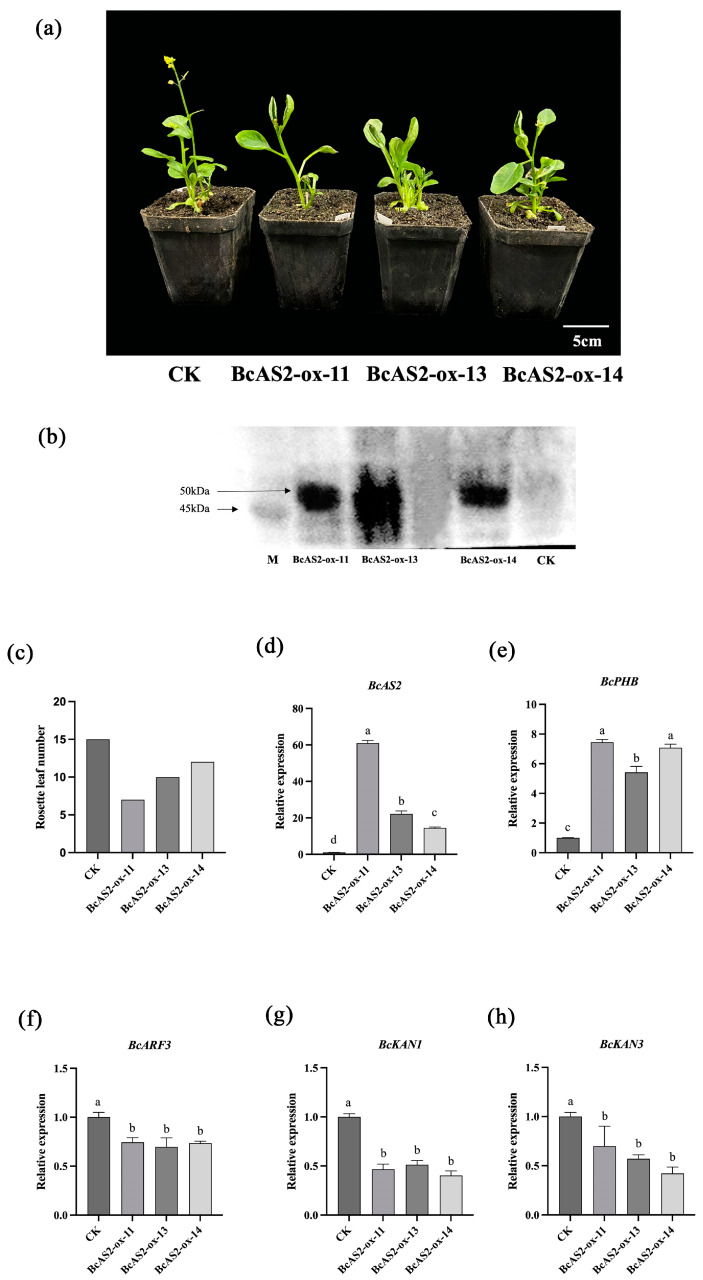 Figure 2
Figure 2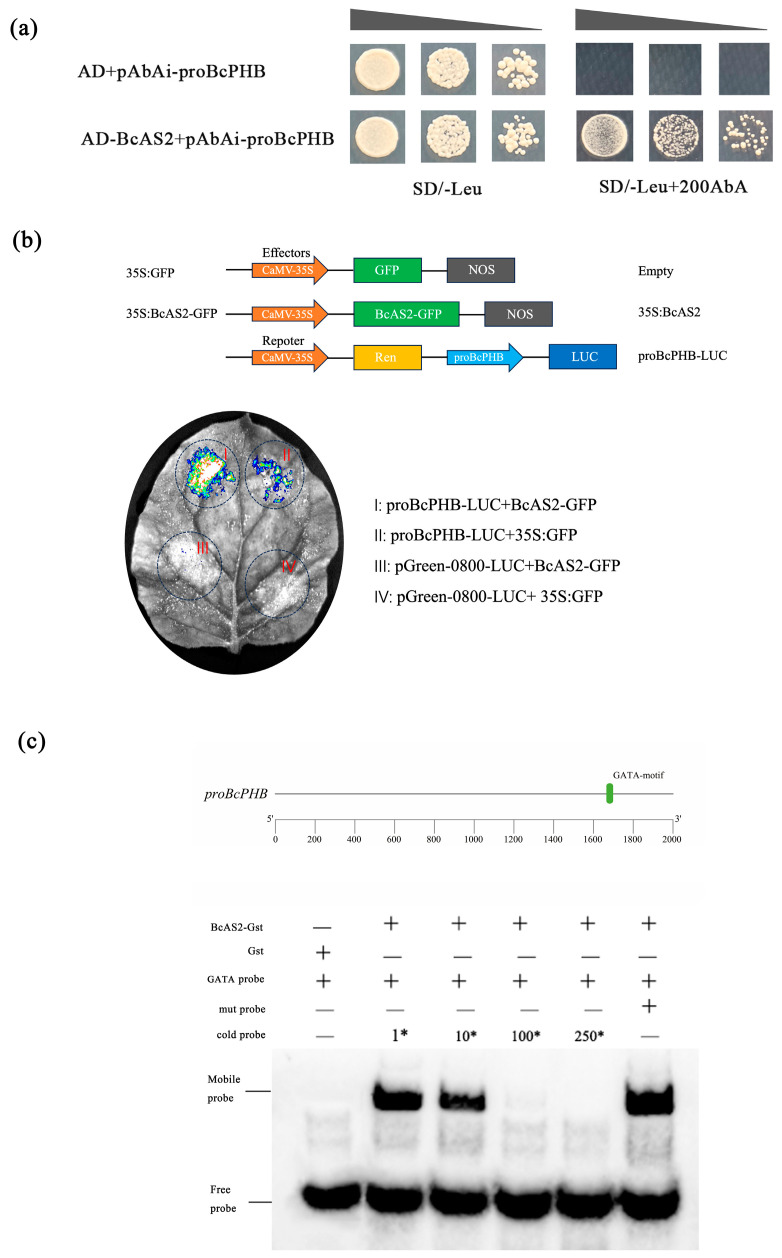 Figure 3
Figure 3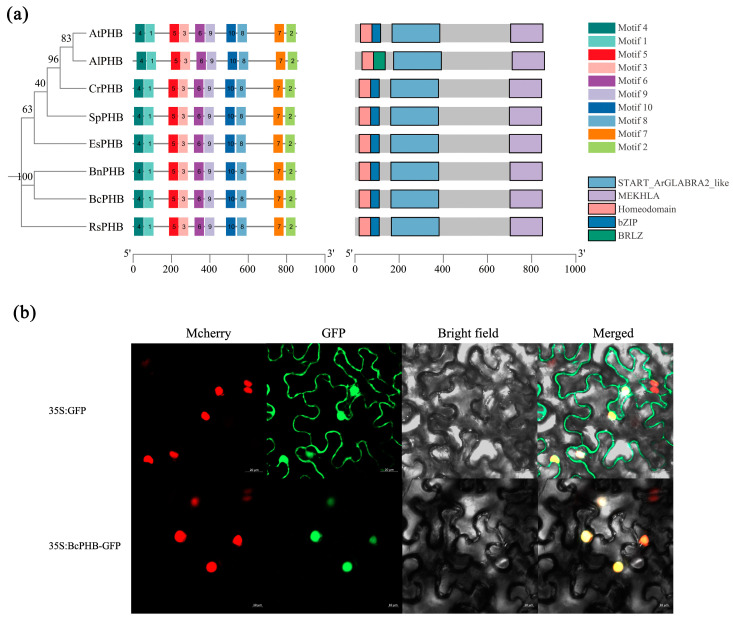 Figure 4
Figure 4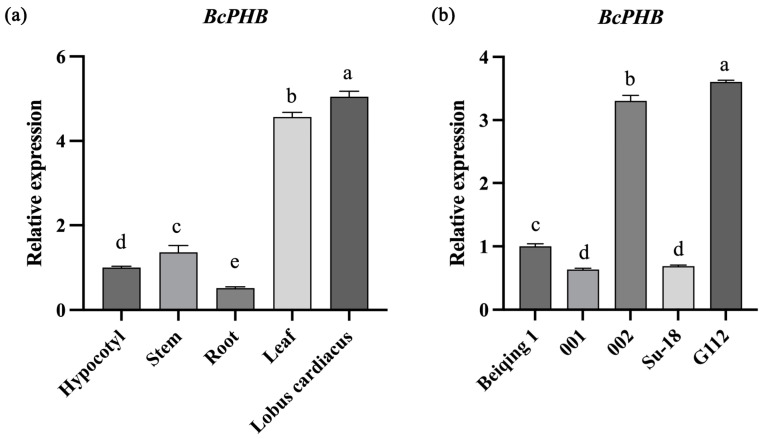 Figure 5
Figure 5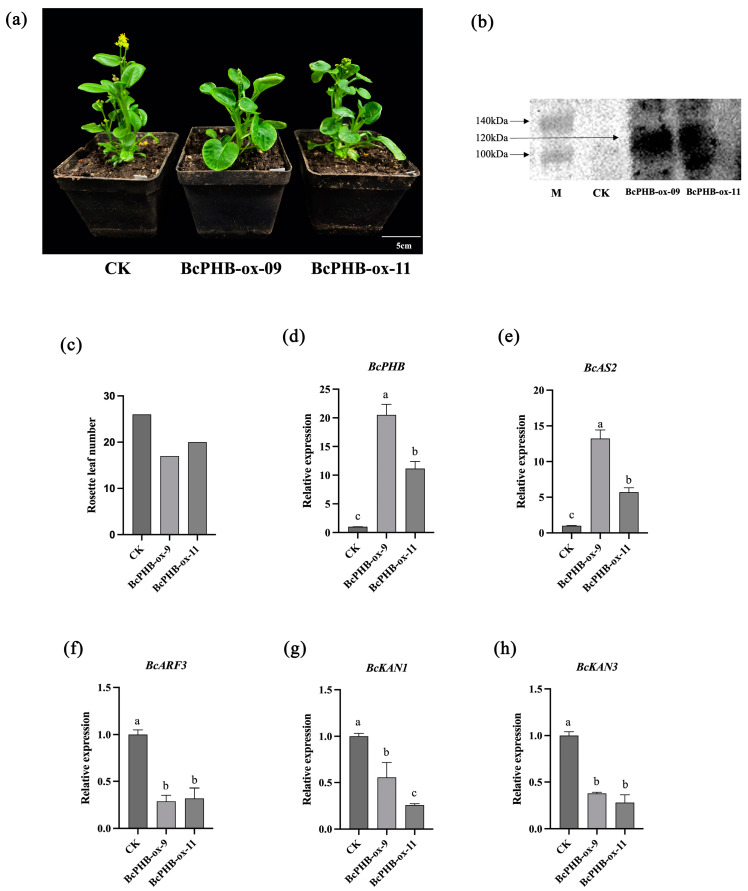 Figure 6
Figure 6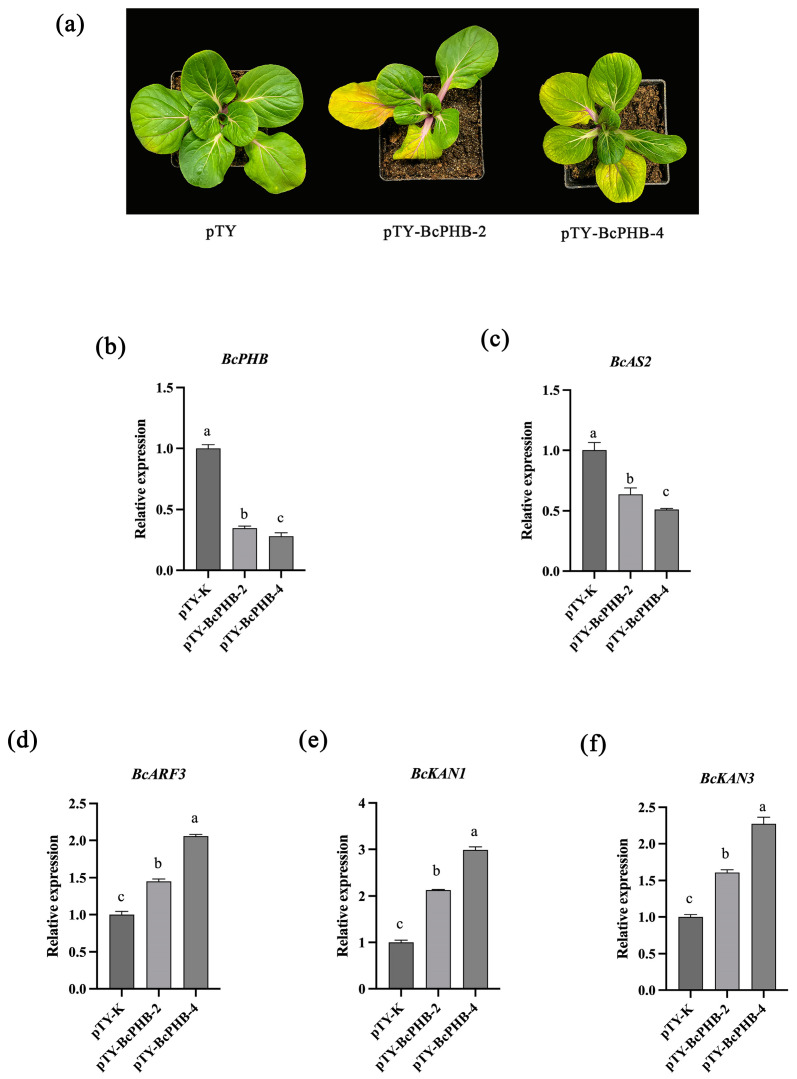 Figure 7
Figure 7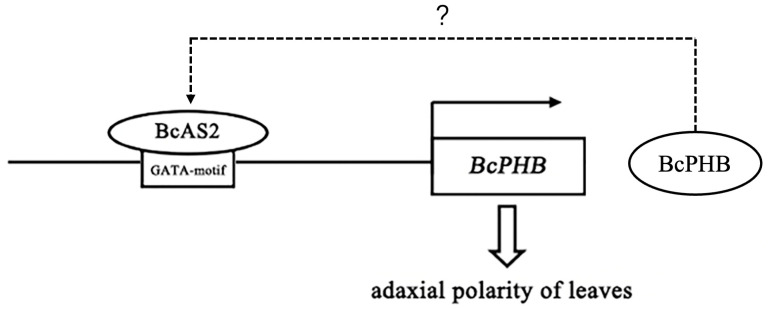 Figure 8
Figure 8- —Jiangsu Seed Industry Revitalization Project
- —Priority Academic Program Development of Jiangsu Higher Education Institutions
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Molecular Biology Research · Plant Reproductive Biology · Plant Gene Expression Analysis
1. Introduction
Leaf morphology plays a critical role in light energy capture and conversion, particularly under low-light conditions, and is recognized as the foundation for plant growth and a direct determinant of crop yield and quality. Leaf morphogenesis is an intricately regulated process governed by the coordinated interaction of genetic networks and environmental factors [1,2]. Notably, leaf morphology critically influences plant architecture, with bilaterally symmetric leaves being essential for proper organogenesis. The establishment of leaf polarity—a central determinant of morphogenesis—serves as the foundation for crop architecture optimization via molecular breeding [3]. Therefore, elucidating the regulatory mechanisms governing leaf morphology is imperative for advancing molecular breeding strategies to improve non-heading Chinese cabbage cultivars.
Two distinct developmental phases have been characterized in leaf morphogenesis: the initial differentiation of leaf primordia from the shoot apical meristem (SAM), followed by subsequent expansion and maturation [4,5]. Within this developmental continuum, leaf polarity establishment emerges as a critical determinant of final morphology [6,7]. Polarity disruptions result in aberrant leaf morphologies, reduced lamina flatness, compromised physiological functionality, and ultimately decreased agricultural productivity [8,9,10].
In Arabidopsis thaliana, the ASYMMETRIC LEAVES 2 (AS2/AtLBD6) transcription factor, a member of the Lateral Organ Boundaries Domain (LBD) family, functions as a master regulator of adaxial-abaxial polarity. AS2 promotes adaxial cell fate specification to shape leaf morphology, with as2 loss-of-function mutants displaying downward-curling leaves [11,12]. Mechanistically, AS2 coordinates a transcriptional network through repression of abaxial determinants (e.g., KANADI family genes) while activating adaxial factors, including Class III Homeodomain-Leucine Zipper (HD-ZIP III) proteins such as PHABULOSA (PHB), PHAVOLUTA (PHV), and REVOLUTA (REV) [13,14,15,16,17]. These HD-ZIP III genes display dynamic spatiotemporal expression patterns, initially enriched within the SAM before becoming progressively restricted to the adaxial domain of developing leaf primordia, where they establish polarity [18,19,20]. This observation suggests these genes contribute to polarity regulation through additive phenotypic effects [21,22]. Their conserved roles across species (e.g., Arabidopsis, rice, raize) underscore their fundamental importance in polarity regulation [23,24,25]. Notably, dominant gain-of-function alleles such as phb-1d and rev-10d induce extreme adaxialization phenotypes, producing needle-like, rod-shaped, or trumpet-like leaves with adaxialized internal tissues [26,27], further demonstrating the necessity of proper polarity for normal leaf development [28]. In addition, leaf curvature modulates photosynthetic efficiency by altering light distribution and gas exchange. For instance, upward-curling leaves in rice crops improve light capture under dense planting, whereas excessive downward curling restricts stomatal conductance. Moderate leaf rolling in corn reduces mutual leaf shading and increases photosynthesis in the population, thereby increasing tolerance to high-density planting [29,30,31]. Optimizing curvature is thus critical for yield potential.
Non-heading Chinese cabbage (Brassica campestris (syn. Brassica rapa) ssp. Chinensis) belongs to the Brassica genus within the Cruciferae family and is cultivated for its nutrient-rich leaves, which determine its commercial value through morphological characteristics [32]. Although previous studies have isolated BcAS2 from the cultivar “NHCC001” and demonstrated its leaf-curling capacity when overexpressed in Arabidopsis [33], the molecular mechanisms governing BcAS2 function in its native species remain poorly understood. Specifically, it remains unknown whether BcAS2 regulates downstream polarity determinants or interacts with conserved HD-ZIP III family members such as BcPHB in non-heading Chinese cabbage.
This study addresses these knowledge gaps through the generation of BcAS2-overexpressing transgenic lines in non-heading Chinese cabbage, which exhibited pronounced adaxialization phenotypes with upward leaf curling. BcPHB was subsequently identified as a direct transcriptional target of BcAS2 using integrated molecular approaches. Functional validation via BcPHB overexpression and silencing confirmed its role in adaxial polarity regulation. These findings elucidate the BcAS2-BcPHB regulatory axis and provide molecular targets for optimizing leaf morphology in Brassica crops through breeding strategies.
2. Results
2.1. Identification and Expression Analysis of AS2 in Related Species
Previous studies have established the essential role of the LBD family transcription factor ASYMMETRIC LEAVES 2 (AS2) in regulating adaxial-abaxial polarity [11,12,34,35,36]. Sequence alignment of AS2 proteins from nine Brassicaceae species revealed that BcAS2 shares 92.3% identity with the Brassica napus BnAS2 (XP_013878412.1), with conserved LOB domains (Leu-35 to Gly-118) (Figure 1a). MEME analysis further identified four conserved motifs (E-value < 1 × 10^−10^) in the AS2 protein across Cruciferae species, with Motif 1 containing the characteristic LBD structural domain, defined by the sequence “CX2CX6CX3C” (Figure 1a). This suggests that AS2 is functionally relatively conserved in most cruciferae species.
To investigate the role of BcAS2 in regulating leaf polarity in non-heading Chinese cabbage, we conducted qRT-PCR analysis in five cultivars—flat-leaved “Beiqing 1” (control), upward-curling “NHCC002” and “G112”, and downward-curling “NHCC001” and “Su-18”. The results showed significantly higher expression of BcAS2 in the upward-curling lines and reduced expression in downward-curling lines (Figure 1b,c), suggesting that BcAS2 promotes adaxial development.
2.2. Phenotypic Analysis of Leaf Polarity in BcAS2-Overexpressing Plants
To further validate the role of BcAS2 in the development of adaxial polarity in “NHCC” leaves, three BcAS2 overexpression lines, BcAS2-overexpressed (BcAS2-ox) (ox-11, ox-13, ox-14) were obtained by Agrobacterium-mediated genetic transformation for subsequent analyses (Figure 2a). It was further verified by qRT-PCR and Western blot experiments (Figure 2b,d). Phenotypic analysis showed that leaves of transgenic plants overexpressing BcAS2 exhibited significant upcurling (Figure 2a), and Rosette leaf numbers decreased by about 35% (Figure 2c). In addition, we assessed the expression levels of other genes known to be involved in the regulation of leaf polarity. To elucidate the potential factors underlying the curling phenotype on the observed leaves of BcAS2 overexpressing plants, we analyzed the relative expression levels of downstream leaf polarity regulators, including BcPHB, BcKAN1, BcKAN3, and BcARF3, found in BcAS2 overexpressing plants (Figure 2d–h). As expected, in BcAS2 overexpressing plants, compared to control plants, qRT-PCR revealed about 5.8-fold upregulation of BcPHB and suppression of abaxial genes (BcKAN1, BcKAN3, BcARF3) in OX lines (Figure 2d–h).
2.3. BcAS2 Directly Activates BcPHB Transcription
The overexpression of BcAS2 significantly altered the expression of downstream leaf polarity regulators, which in turn affects the leaf polarity of nonbearing cabbage. Notably, BcAS2 had a significant effect on the expression level of BcPHB. These observations suggest a correlation between BcAS2 and BcPHB, which has the potential to regulate the downstream leaf polarity development pathway in Pak-choi. Yeast one-hybrid (Y1H) assays demonstrated direct BcAS2 binding to the BcPHB promoter (Figure 3a). The dual luciferase reporter system further confirmed that BcAS2 could directly bind to the BcPHB promoter, promoting its transcription (Figure 3b). To further understand the interaction between transcription factors and promoters, the BcPHB promoter sequence was analyzed, and the possible binding sites for BcAS2 were predicted. Electrophoretic mobility shift assays (EMSA) confirmed sequence-specific binding to the GATA-motif (−752 to −739 bp), with binding abolished by cold competition but unaffected by mutated probes (GATA→CTCT) (Figure 3c). These findings establish BcAS2 as a direct transcriptional activator of BcPHB.
Based on these results, it is hypothesized that BcAS2 may regulate the development of leaf blade adaxial polarity in “NHCC” leaves through the BcAS2-BcPHB pathway. The researchers investigated the effect of BcAS2 on BcPHB transcription using a dual luciferase assay. The results showed that the fluorescence intensity of 35S:BcAS2-GFP and proBcPHB-luc was significantly higher than the combination of 35S:GFP + proBcPHB-luc. This confirmed enhanced transcriptional promotion of BcPHB by BcAS2 (Figure 3b). It was further demonstrated by EMSA experiments that BcAS2 could directly bind to the BcPHB promoter in vitro through the binding site GATA-motif (Figure 3c). In conclusion, BcAS2 regulates leaf polarity development by directly promoting the transcription of BcPHB, which promotes the development of leaf adaxial polarity.
2.4. Characterization of BcPHB
Numerous studies have demonstrated that the HD-ZIP III (Class III Homeodomain-leucine Zipper) family gene PHB is a key factor in the establishment of adaxial polarity in plant leaves and plays a crucial role in the regulation of leaf polarity development in a variety of crops [21]. In order to elucidate the sequence structure of BcPHB, we performed a comparative analysis of PHB amino acid sequences in a variety of horticultural and model plants. Phylogenetic analysis showed that BcPHB is closely related to BnPHB as well as to other crucifers (Figure 4a). MEME analysis showed that PHBs have the same motif composition and order in the crucifers. As shown in Figure 4a, PHB has a conserved SANT structural domain in all plants except AlPHB, suggesting that PHB is relatively functionally conserved in most plants. Analysis of the conserved structural domains showed that AtPHB (Arabidopsis thaliana), BnPHB (Brassica napus), RsPHB (Raphanus sativus), EsPHB (Eutrema salsugineum), SpPHB (Schrenkiella parvula), CrPHB (Capsella rubella), AlPHB (Arabidopsis lyrata), and BcPHB (Brassica campestris (syn. Brassica rapa) ssp. Chinensis) exhibited relatively conserved patterns, with each structural domain containing the START_ArGLABRA2_like, MEKHLA, and Homeodomain structural domains (Figure 4a). The above results indicate that PHB is functionally more conserved in Cruciferae species. The subcellular localization of BcPHB was detected by transient analysis of 35S: BcPHB-GFP, where the GFP reporter was translated and fused to the 3′ end of BcPHB cDNA. The results showed that BcPHB was localized to the nucleus (Figure 4b). In conclusion, as a member of the HD-ZIP III family of transcription factors, BcPHB is structurally and functionally similar to other PHBs in plants.
2.5. Expression Pattern of BcPHB
In order to verify the expression pattern of BcPHB in different tissues of “NHCC”, a qPCR assay was carried out to determine its expression level using a 1-month-old sample “NHCC001”. The results showed that high BcPHB expression in young leaves and apical buds, with minimal expression in roots and hypocotyls (Figure 5), is consistent with its role in leaf morphogenesis [16]. In addition, BcPHB expression positively correlated with BcAS2 levels across cultivars, further supporting their functional linkage (Figure 5b).
2.6. Phenotypic Analysis of Leaf Polarity in BcPHB-Overexpressing Plants
To further validate the role of BcPHB in the development of adaxial polarity in “NHCC” leaves, two BcPHB overexpressing lines, BcPHB-overexpressed (BcPHB-ox) (ox-09, ox-11), were obtained by Agrobacterium-mediated genetic transformation for subsequent analyses (Figure 2a). It was further verified by qRT-PCR and Western blot experiments (Figure 6b,d). Phenotypic analysis showed that leaves of transgenic plants overexpressing BcPHB exhibited significant upcurling (Figure 6a), and the number of rosette leaves in BcAS2-OX plants was reduced by about 30% compared to null-loaded control plants (Figure 6c). To elucidate potential factors underlying the curled phenotype on the observed leaves of BcPHB overexpressing plants, we analyzed relevant leaf polarity regulators, including BcAS2, BcKAN1, BcKAN3, and BcARF3, and found that the relative expression levels of BcPHB in overexpressing plants (Figure 6e–h) were, as expected, significantly reduced in comparison to control plants, with the BcPHB-OX plants showed significantly lower mRNA levels of BcKAN1, BcKAN and BcARF3 (Figure 6f–h). In contrast, the mRNA levels of BcAS2 were significantly higher in BcPHB-OX plants than in control plants (Figure 6e).
2.7. Phenotypic Analysis of BcPHB-Silenced Plants
To further investigate the function of BcPHB, we employed the virus-induced gene silencing (VIGS) technique to silence BcPHB in Pak-choi plants. The control plants (pTY) were injected with an empty pTY vector as a control (Figure 7a). It was observed that the leaves of BcPHB-silenced plants exhibited a downward curling phenotype compared to control plants (Figure 7a). BcPHB expression in silenced plants was reduced to about 28% of the control compared with control plants (Figure 7b). Analysis of the expression levels of BcAS2, BcARF3, BcKAN1, and BcKAN3 (Figure 7c–f) revealed that BcKAN1, BcKAN3, and BcARF3 were upregulated, while BcAS2 was downregulated in BcPHB-silenced plants (Figure 7c).
3. Discussion
Leaf polarity establishment is a fundamental process for normal plant growth and development, governed by complex genetic regulatory networks [37]. Previous studies have found that only when the adaxial-abaxialaxis polarity is established can the leaf begin to develop normally [1,38]. Ye Lin [33] isolated the BcAS2 gene, which has a typical LOB structural domain and is homologous to AS2 in Arabidopsis thaliana, from the non-coccluding cabbage cultivar “NHCC001”, and initially explored its biological role in non-coccluding cabbage, but the downstream regulatory pathways have not yet been developed. The downstream regulatory pathway has not been investigated. Therefore, we further investigated the downstream leaf polarity regulators regulated by BcAS2.
Our study provides compelling evidence that BcAS2 plays a pivotal role in regulating adaxial-abaxial polarity in non-heading Chinese cabbage leaves. Overexpression of BcAS2 induced pronounced upward leaf curling, a phenotype reminiscent of Arabidopsis mutants with disrupted abaxial polarity determinants, such as kan1 kan2 double mutants and arf3 arf4 mutants [19,39,40]. The observed suppression of abaxial identity genes (BcKAN1, BcKAN3, BcARF3) and concomitant activation of the adaxial regulator BcPHB in BcAS2-overexpressing (OX) plants suggests a conserved regulatory hierarchy similar to that in Arabidopsis, where AS2 antagonizes abaxial-promoting factors while reinforcing adaxial fate [16,26,41,42,43].
Intriguingly, the number of rosette leaves in BcAS2-OX plants was reduced by about 35%, contrasting with reports in tomato (Solanum lycopersicum) by Xu, where overexpression of SlAS2 increased leaf number [44]. This discrepancy may reflect species-specific subfunctionalization of AS2 orthologs. In eudicots like tomato, AS2 may primarily regulate leaf initiation through modulation of cell cycle genes such as CYCD3 [45], whereas in Brassica species, BcAS2 appears to prioritize the optimization of leaf curvature, potentially to enhance photosynthetic efficiency under specific environmental conditions. This functional divergence highlights the evolutionary plasticity of AS2-mediated regulatory networks in shaping leaf architecture across taxa. In addition, this compensatory growth pattern (i.e., reduced leaf number but increased individual leaf area in OX lines) mirrors observations in rice HD-ZIP III mutants, where prolonged SAM activity redirected resources towards post-initiation leaf expansion [46]. Such conserved trade-offs suggest a universal developmental strategy to optimize photosynthetic output under varying organ initiation rates. Further field evaluations are needed to determine if this restructuring ultimately benefits yields under agronomic conditions
Mechanistically, we demonstrated that BcAS2 directly binds to the GATA motif within the BcPHB promoter to activate its transcription. This interaction was rigorously validated through multiple experimental approaches: yeast one-hybrid assays confirmed physical binding, dual-luciferase reporter assays quantified transcriptional activation (3.2-fold enhancement), and EMSA resolved the specificity of BcAS2 for the GATA sequence (Figure 3). The functional significance of this regulatory axis was further corroborated by phenotypic analyses of BcPHB-OX and BcPHB-silenced plants. Overexpression of BcPHB recapitulated the upward-curling phenotype of BcAS2-OX lines, while silencing BcPHB via virus-induced gene silencing (VIGS) resulted in downward-curling leaves (Figure 6 and Figure 7). These reciprocal phenotypes underscore BcPHB’s central role in adaxial polarity establishment, consistent with prior studies in Arabidopsis, where HD-ZIP III genes like PHB specify adaxial identity [15,16,47,48].
Notably, we identified a feedback regulatory loop between BcAS2 and BcPHB. While BcAS2 directly activates BcPHB transcription, BcPHB overexpression conversely upregulates BcAS2 expression (Figure 6e). Conversely, BcPHB silencing downregulated BcAS2 (Figure 7c). The mutual regulation of BcAS2 and BcPHB implies a stabilizing feedback loop. While yeast one-hybrid assays confirmed that BcPHB cannot directly bind to the BcAS2 promoter (Figure S2), we propose that BcPHB indirectly stabilizes BcAS2 expression, potentially via auxin signaling pathways [43]. However, the intermediary factors bridging this regulatory relationship remain to be elucidated.
The conservation of the AS2-PHB regulatory module across Brassicaceae is striking. Phylogenetic analyses revealed high sequence similarity between BcAS2/BnAS2 and AtAS2, with preserved LOB domains and motif architectures (Figure 1a). While motif5 is absent in NHCC-002-derived BcAS2 compared to NHCC-001, both variants preserve the intact LOB domain required for polarity regulation (Figure 1a) [12]. Complete sequence conservation within the LOB domain (motifs 1/2/7) was confirmed (Figure S1), supporting functional conservation despite motif5 divergence. Similarly, BcPHB shares conserved functional domains (SANT, MEKHLA, Homeodomain) with its Arabidopsis counterpart (Figure 4a), supporting functional conservation. However, subtle differences in expression patterns (e.g., tissue-specific BcPHB enrichment in young leaves; Figure 5) may reflect species-specific adaptations in leaf development regulation.
From an applied perspective, this study provides actionable targets for molecular breeding. The BcAS2-BcPHB pathway could be manipulated to engineer leaf curvature, a trait critical for optimizing light interception in dense planting systems. For instance, moderate overexpression of BcAS2 or BcPHB might reduce mutual shading among leaves, potentially enhancing photosynthetic efficiency and yield. Conversely, targeted suppression of this pathway could benefit cultivars requiring flatter leaves for mechanical harvesting. In addition, while our data provide suggestive evidence of biomass advantages in upward-curling lines, this observation requires rigorous field validation, particularly given the documented genotype × environment interactions influencing yield-related traits mediated by leaf morphology [49].
In conclusion, our work delineates a linear regulatory pathway in which BcAS2 directly activates BcPHB transcription to promote adaxial polarity in non-heading Chinese cabbage leaves (Figure 8). This mechanism integrates with broader polarity networks involving KANADI and ARF family genes, forming a robust system to coordinate leaf symmetry. These findings not only advance our understanding of leaf development in Brassica crops but also establish a framework for precision breeding of leaf morphology traits.
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
Non-heading Chinese cabbage cultivars “NHCC001” and “49 Caixin” (Brassica campestris (syn. Brassica rapa) ssp. Chinensis), and tobacco (Nicotiana benthamiana) seedlings were cultivated in growth chambers (ZRX-380; Kesheng Experimental Instruments Co., Ltd., Ningbo, China) at 21 ± 2 °C under 16/8 h light/dark cycles (250 μmol·m^−2^·s^−1^, 40% relative humidity (RH)). Seeds of the experimental materials used in this study were were generously provided by the Laboratory of Cabbage Systems Biology at Nanjing Agricultural University, Nanjing, China. Leaf discs and plants were maintained in organic-rich substrate. Samples were flash-frozen in liquid nitrogen and stored at −80 °C.
4.2. Cloning and Analysis of BcPHB
BcPHB coding sequence (CDS) was amplified using specific primers and cloned into the pRI101-GFP vector (NdeI/KpnI sites). Homologous sequences were identified via the online BLAST tool (https://blast.ncbi.nlm.nih.gov/Blast.cgi, accessed on 15 March 2024). Multiple sequence alignment and conserved motif analysis were performed using MEGAX 7.0 and MEME website (http://meme-suite.org/tools/meme, accessed on 1 May 2024). A neighbor-joining phylogenetic tree was constructed with 1000 bootstrap replicates. Primers are listed in Table S1.
4.3. Subcellular Localization of BcPHB in Tobacco
BcPHB CDS (without stop codon) was fused to pRI101-GFP. BcPHB:GFP and empty 35S:GFP vectors were transformed into Agrobacterium tumefaciens GV3101 (OD600 = 0.8–1.0, wavelength of 600 nm) and infiltrated into tobacco leaves. Fluorescence signals were captured after 48–60 h using a confocal microscope (Zeiss LSM 500, Oberkochen, Germany). Primers are listed in Table S1.
4.4. Vector Construction and Transgenic Plant Generation
Full-length BcAS2 and BcPHB CDS were cloned into pRI101 to generate overexpression vectors, and pRI101 was empty as a CK control. Transgenic lines were generated via Agrobacterium-mediated transformation (GV3101 strain) of NHCC cotyledon explants [50], NHCC transgenic overexpression (OE) lines were established through tissue culture-mediated transformation, with successful transformation validated by qRT-PCR and Western blotting. Refer to (File S1) for specific methodology. Primers are listed in Table S1.
4.5. BcPHB Silencing via VIGS
Virus-induced gene silencing (VIGS) was performed as described in a previous study [51]. The target gene BcPHB 40 bp interfering fragment and its antisense sequence (ATGAACAAACTCTTGATGGAAGAGAATGATCGTCTTCAGATCTGAAGACGATCATTCTCTTCCATCAAGAGTTTGTTCAT) were inserted into the pTY vector. Gold particle-coated pTY-Empty (control) and pTY-BcPHB plasmids were bombarded into NHCC001 seedlings using a gene gun (1300 psi, PDS-1000/He, Bio-Rad, Hercules, CA, USA). Plants were grown under controlled conditions and sampled for qRT-PCR.
4.6. RNA Extraction and qRT-PCR
Total plant RNA was extracted using the RNA simple Total RNA Kit (No. DP432; Tengen Biotechnology Co., Ltd., Beijing, China), and reverse transcription of the RNA was performed using the Hifair^®^ V one-step RT-gDNA digestion Super Mix for qPCR (No. 11142ES10; Yesheng Co., Ltd., Shanghai, China) was used for reverse transcription of RNA. Samples were analyzed by quantitative polymerase chain reaction (qPCR) using Hieff^®^ qPCR SYBR Green Master Mix (No. 11201ES08; YEASEN, Shanghai, China) and CFX Connect Real-Time PCR Detection System (Bio-Rad, Hercules, CA, USA) according to the manufacturer’s instructions. Expression data were calculated using the 2^−ΔΔCt^ method with three independent replicates [52], the relative expression levels of the target genes were calculated, and the results of NHCC were normalized using BcGAPDH (BraC08g031360.1) as an internal reference gene [53]. Primers are listed in Table S1.
4.7. Yeast One-Hybrid (Y1H) Assay
The 2000 bp sequence upstream of the BcPHB transcriptional start site was used as the promoter region, and the amplified DNA target fragment was inserted into the pAbAi vector to obtain the bait vector pAbAi-BcPHB. The CDS, the terminatorless codon of BcAS2, was fused to the pGADT7(AD) vector. Yeast one-hybrid (Y1H) assays were performed following the Yeast Protocols Handbook (Clontech, San Jose, CA, USA). Bait (pAbAi-BcPHB) and prey (pGADT7-BcAS2) vectors were co-transformed into Y1H Gold yeast strains. Protein-DNA interactions were assessed on SD/-Ura/-Leu plates containing aureobasidin A (AbA) at optimized concentrations. Refer to (File S2) for specific methodology. Primers are listed in Table S1.
4.8. Dual-Luciferase Assay
The dual luciferase assay was performed according to the method of Wang et al. [54]. The BcPHB promoter sequence was inserted into the pGreen0800-LUC vector to obtain a reporter vector. The BcAS2 stop codon-free CDS was cloned into the pRI101 vector to generate an effector vector. The vector product was then transformed into A. tumefaciens GV3101 (psoup) (Tolo Biotech, Shanghai, China). Agrobacterium cells containing the reporter gene and effector constructs were injected into tobacco leaves at a ratio of 1:9. After 48–96 h, fluorescence at the injection site was detected using a Tanon 4600 (Shanghai, China) in vivo imager. Activity levels of firefly luciferase (LUC) and Ray nira luciferase (REN) were determined using the Dual Luciferase Reporter Kit (No. 11402ES60; YEASEN) according to the manufacturer’s instructions. Primers are listed in Table S1.
4.9. Electrophoretic Mobility Shift Assay (EMSA)
BcAS2 was cloned into the pGEX-4T-1 (GST) vector, and the resulting recombinant plasmid was then transformed into E. coli (DE3) fertile cells. Oligonucleotide probes were synthesized using online resources (http://www.tsingke.net/, accessed on 23 May 2024) and labeled with biotin at the 5′ and 3′ ends. The biotin-labeled probes and mutation probes were synthesized by Shanghai Sangong Biotechnology Co. (Shanghai, China). Double-stranded DNA probes were prepared by annealing complementary oligonucleotides. The annealing process consisted of heating the samples to 95 °C for 3 min, then gradually cooling the samples from 1 °C to 25 °C at 90 s intervals and separating the bound probes from the probes on a nondenaturing acrylamide gel. The electrophoretic mobility shift assay (EMSA) was performed using a chemiluminescent EMSA kit (Shanghai Beo Tianmei Biotechnology Co., Ltd., Shanghai, China) and imaged with a protein imaging system (Tanon 4600; Tanon Science & Technology Co., Ltd., Shanghai, China). Details of the DNA probes used are shown in Table S1.
4.10. Statistical Analysis
Experiments included three biological and technical replicates. Statistical analyses used ANOVA with Tukey’s test (p < 0.05).
5. Conclusions
This study we findings establish the BcAS2-BcPHB pathway as a key regulator of leaf polarity in non-heading Chinese cabbage. Overexpression of BcAS2 induces pronounced upward leaf curling by directly activating BcPHB transcription, which suppresses abaxial identity genes (e.g., BcKAN1, BcKAN3, BcARF3) and reinforces adaxial cell fate. Molecular validation through Y1H, dual-luciferase, and EMSA assays confirmed that BcAS2 binds the GATA-motif in the BcPHB promoter to enhance its expression. Reciprocal experiments—BcPHB overexpression phenocopying BcAS2-OX lines and BcPHB silencing causing downward curling—definitively link BcPHB to adaxial polarity regulation. Furthermore, the mutual regulatory loop between BcAS2 and BcPHB highlights a stabilizing mechanism for adaxial identity. This study provides both mechanistic insights into leaf polarity establishment in Brassica crops and actionable targets for molecular breeding of leaf morphology traits.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bowman J.L. Eshed Y. Baum S.F. Establishment of polarity in angiosperm lateral organs Trends Genet.20021813414110.1016/S 0168-9525(01)02601-411858837 · doi ↗ · pubmed ↗
- 2Bowman J.L. Axial patterning in leaves and other lateral organs Curr. Opin. Genet. Dev.20001039940410.1016/S 0959-437X(00)00103-910889056 · doi ↗ · pubmed ↗
- 3Byr Sne M. Timmermans M. Kidner C. Martienssen R. Development of leaf shape Curr. Opin. Plant Biol.20014384310.1016/S 1369-5266(00)00133-311163166 · doi ↗ · pubmed ↗
- 4Hudson A. Axioms and axes in leaf formation?Curr. Opin. Plant Biol.19992566010.1016/S 1369-5266(99)80011-910047567 · doi ↗ · pubmed ↗
- 5Byrne M.E. Barley R. Curtis M. Arroyo J.M. Dunham M. Hudson A. Martienssen R.A. Asymmetric leaves 1 mediates leaf patterning and stem cell function in Arabidopsis Nature 200040896797110.1038/3505009111140682 · doi ↗ · pubmed ↗
- 6Douglas S.J. Chuck G. Dengler R.E. Pelecanda L. Riggs C.D. KNAT 1 and ERECTA regulate inflorescence architecture in Arabidopsis Plant Cell 20021454755810.1105/tpc.01039111910003 PMC 150578 · doi ↗ · pubmed ↗
- 7Venglat S.P. Dumonceaux T. Rozwadowski K. Parnell L. Babic V. Keller W. Martienssen R. Selvaraj G. Datla R. The homeobox gene BREVIPEDICELLUS is a key regulator of inflorescence architecture in Arabidopsis Proc. Natl. Acad. Sci. USA 2002994730473510.1073/pnas.07262609911917137 PMC 123716 · doi ↗ · pubmed ↗
- 8Chien J.C. Sussex I.M. Differential regulation of trichome formation on the adaxial and abaxial leaf surfaces by Gibberellins and photoperiod in Arabidopsis thaliana (L.) Heynh Plant Physiol.19961111321132810.1104/pp.111.4.13218756507 PMC 161017 · doi ↗ · pubmed ↗