Identification of Causal Agents of Rust of Saccharum spp. and Assessment of Resistance to Brown Rust in Erianthus arundinaceus Clones and Their Offspring
Jun-Lv Chen, Sheng-Ren Sun, Zhu-Qing Wang, Hua-Ying Fu, Huan-Yin Xu, Hai-Long Chang, San-Ji Gao, Qin-Nan Wang

TL;DR
This study identifies rust-causing pathogens in sugarcane and finds many Erianthus arundinaceus clones resistant to brown rust, including some without the known resistance gene.
Contribution
The study reveals new resistance sources in E. arundinaceus and suggests the presence of unknown resistance genes for brown rust.
Findings
17.9% of samples were infected with Puccinia melanocephala and 34.8% with Puccinia kuehnii.
61 E. arundinaceus clones showed high resistance to brown rust, some lacking the Bru1 gene.
13 offspring and 6 backcross parents of the 'ROC' series carried the Bru1 resistance gene.
Abstract
Sugarcane rust diseases are caused by Puccinia melanocephala (brown rust) and Puccinia kuehnii (orange rust), and significantly threaten the sustainable and stable development of the global sugarcane industry. Erianthus arundinaceus within the Saccharum complex is a potential germplasm resource for sugarcane breeding and is characterized by its tolerance of infertile land, drought, and diseases. However, the research on resistance to rust in E. arundinaceus clones and their offspring (F1 and backcross with modern sugarcane varieties) is limited. In this study, a total of 201 leaf samples from Saccharum spp. hybrids with rust symptoms were collected and screened for disease occurrence. PCR detection revealed that 17.9% and 34.8% of the samples were infected by P. melanocephala and P. kuehnii, respectively. Additionally, 12.9% of the samples were infected by both pathogens. A total of 88…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
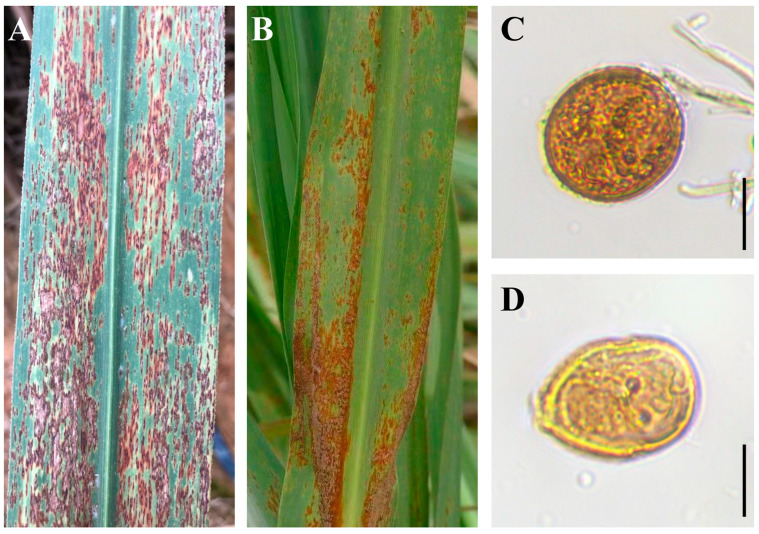 Figure 1
Figure 1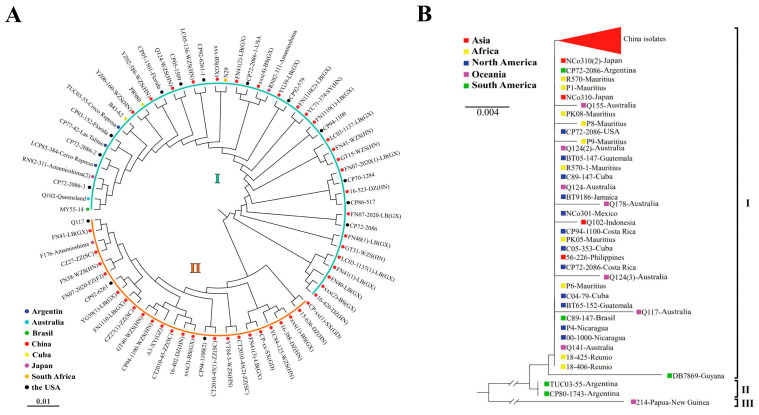 Figure 2
Figure 2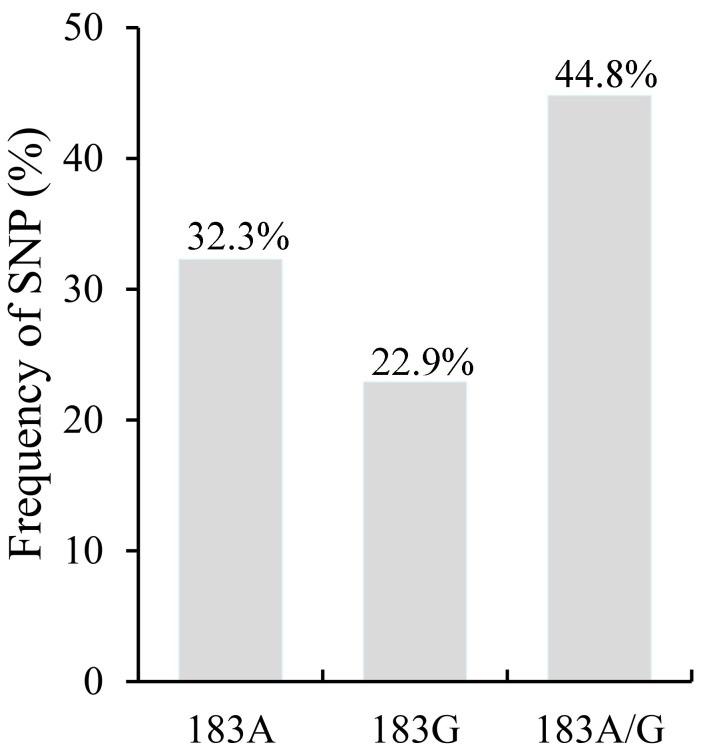 Figure 3
Figure 3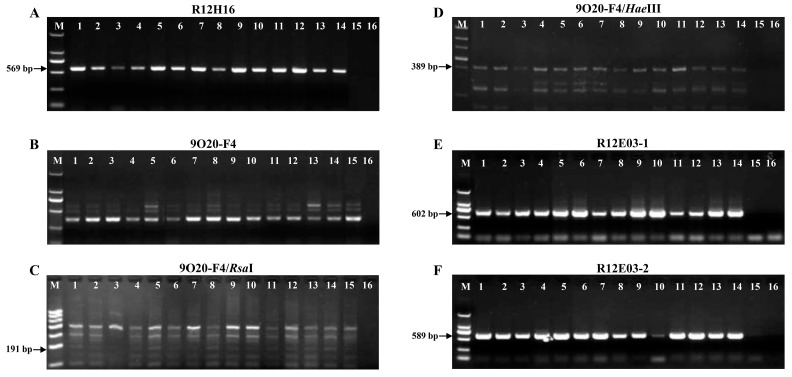 Figure 4
Figure 4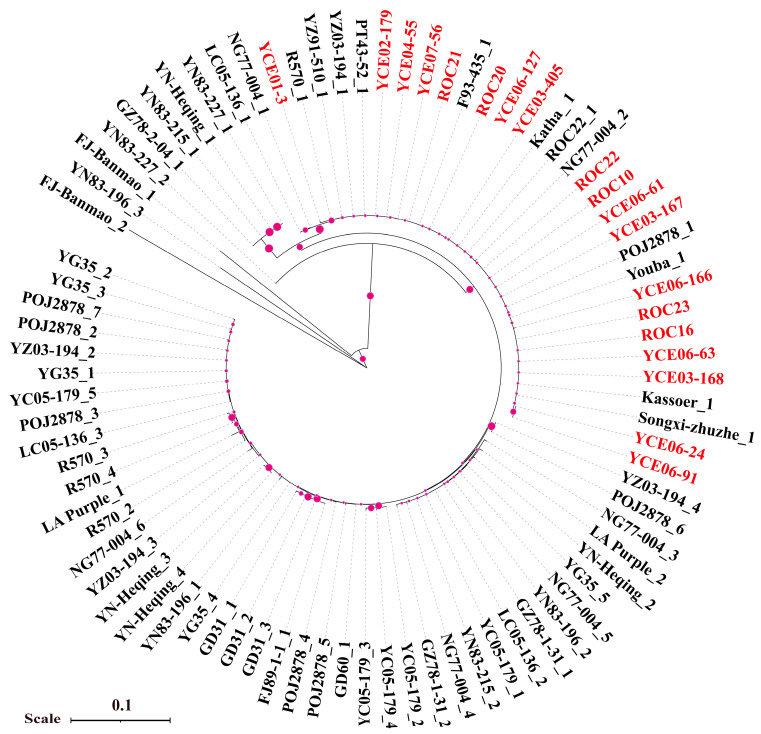 Figure 5
Figure 5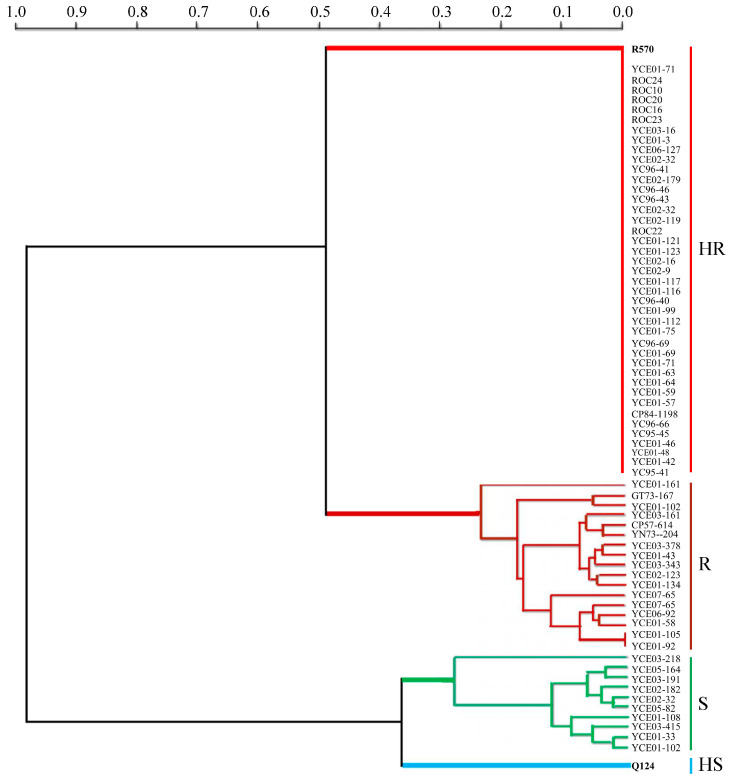 Figure 6
Figure 6- —GDAS’s Project of Science and Technology Development
- —Earmarked Fund for China Agriculture Research System
- —GDAS’s Project of Technical Innovation and Incubation Service Platform Construction
- —Special Project for Rural Revitalization Strategy in Guangdong Province
- —Young Talent Project of GDAS
- —Ministry of Science and Technology plans of China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsSugarcane Cultivation and Processing · Plant Pathogenic Bacteria Studies · Biofuel production and bioconversion
1. Introduction
Sugarcane (Saccharum spp. hybrids) is widely distributed in tropical and subtropical regions, accounting for approximately 80% of the sugar and 40% of the biofuel production worldwide. The cultivation of sugarcane relies on the vegetative propagation of stalks, leading to the accumulation of pathogenic microorganisms year-by-year, resulting in cultivar degeneration and quality degradation at both plant and ratoon crops [1,2]. The occurrence and prevalence of rust leads to reduced tillering numbers and thinner stems of sugarcane, giving rise to large-scale yield reductions (10–50%) and significant decreases in biomass (12–40%) [3,4]. Two types of rust have been found in sugarcane: brown rust caused by Puccinia melanocephala H. and P. sydow, and orange rust caused by P. kuehnii E. J. Butler [5]. Both rusts have been reported in the main sugarcane-growing countries, including Brazil, India, China, the United States, and Australia, which poses a great challenge to the safety and sustainable development of sugarcane cultivation [1,6].
In mainland China, brown and orange rust were reported in Yunnan and Fujian provinces in the early of 1980s, respectively, while the two sugarcane fungal diseases subsequently spread throughout China and have been major sugarcane diseases [7], resulting in 24.9% yield losses and a 3.1% sugar content reduction [8]. Currently, chemical fungicide application is an effective management strategy for controlling rust diseases [9]. In addition, the utilization of resistant cultivars developed through breeding programs is a longstanding and successful practice. Two methods of disease-resistance identification, natural inoculum pressure and artificial inoculation, are used for screening sugarcane resistance to rust in breeding programs [1]. The identification of resistance to brown rust in new elite sugarcane varieties under natural field conditions has been conducted in China, which indicated that 25% of varieties have high resistance to brown rust [10]. Notably, the main variety YZ08-1609 and some “ROC” serial varieties are highly resistant, but the other main varieties LC05-136, GT42, and GT44 are susceptible or moderately susceptible to brown rust [10]. This resistance to brown rust was identified by the artificial inoculation method in 31 wild core germplasms of the Saccharum complex, and seven tested clones of Erianthus arundinaceus were highly resistant to this disease [11]. Few reports were found about the identification of sugarcane resistance to orange rust in China.
The identification of sugarcane resistance to brown rust relies on molecular markers, of which brown rust 1 (Bru1) is the most widely used. The self-bred progenies of the rust-resistant variety R570 showed an obvious separation ratio of 3:1 (resistant/susceptible) after inoculation with brown rust, and the resistance gene (Bru1) was consequently identified using amplified fragment length polymorphism (AFLP) and bulked segregant analysis (BSA) technologies [12]. A study by Costet et al. used 22 molecular markers linked to Bru1 at an interval of 8.2 centimorgan (cM) in the variety R570 to analyze 380 sugarcane accessions, including modern cultivated varieties and breeding materials representing worldwide diversity. Their results revealed that 86% of 194 resistant sugarcane accessions carried the Bru1 gene, indicating that this gene is a main source of resistance to brown rust in modern sugarcane varieties [13]. Subsequently, Bru1 was widely used for sugarcane breeding for resistance to brown rust [14,15]. Regarding orange rust, some molecular markers, including the quantitative trait loci (QTLs), such as G1 and Oru1/2/3/4/5, together with single nucleotide polymorphisms (SNPs) and insertion/deletions (InDels), have been linked to orange rust resistance in sugarcane, but the application of these markers is still limited in sugarcane breeding practices [16,17].
The Saccharum complex, including Saccharum, Erianthus, Miscanthus, Narenga, and Sclerostachya, shares common agronomic traits [18]. It is a crucial approach to introduce novel and elite resistance genes into sugarcane cultivars by distant hybridization between wild clones in the Saccharum complex and modern sugarcane varieties. Ensuring the resistance of wild germplasm resources is a priority procedure in sugarcane breeding programs. A study by Parco et al. showed that, among 1282 accessions from the world sugarcane germplasm resource database, the highest frequency of Bru1 was found in S. barberi (79.3%) and S. sinense (71.8%), followed by S. robustum (59.1%), S. officinarum (26.4%), the interspecific hybrid species (21.0%), and S. spontaneum (18.8%) [19]. In China, the two markers R12H16 and 9O20-F4 were used to screen for Bru1 in sugarcane clones and the results showed that 69.2% (18/26) of Chinese clones (resistant to brown rust) harbor this gene, which suggested that the donor of Bru1 in these clones is possibly from S. spontaneum or S. robustum of New Guinea [14]. Unexpectedly, Bru1 was absent in the clones of E. arundinaceus [11,14].
In this study, we investigated the distribution and incidence of the causal agents of rust in the main sugarcane planting provinces of China. Resistance identification for brown rust and molecular detection of Bru1 markers among E. arundinaceus and their offspring were carried out. These findings will provide an important clue for preventing and controlling rust and supply some novel wild germplasm resources in sugarcane breeding for rust resistance.
2. Materials and Methods
2.1. Collection of Rust Samples and Fungal Microscopy in China
A total of 201 leaf samples of Saccharum spp. hybrids with rust symptoms were collected from seven provinces of China, including Guangdong, Guangxi, Yunnan, Sichuan, Guizhou, Fujian, and Hainan, from 2013 to 2019. Another 88 clones consisting of E. arundinaceus offspring and their parents were from Hainan Breeding Research Base of Nanfan & Seed Industry, Guangdong Academy of Sciences (Guangzhou, China) during 2018–2019. After cleaning the leaf surface with 75% alcohol, the leaves were kept in a −80 °C refrigerator until further experiments. The information on these samples from Saccharum spp. hybrids is shown in Table S1. The samples of E. arundinaceus offspring and their pedigree information are listed in Tables S2 and S3.
Fungal spores were collected from rust-infected leaves of sugarcane for morphological analysis. The distilled water was dropped on a glass slide, and then spores were gently transferred into the water and spread evenly using a dissecting needle. The representative isolates of P. kuehnii and P. melanocephala were observed under a light microscope (BX53 microscope, Olympus, Hachioji, Japan).
2.2. Polymerase Chain Reaction (PCR) Detection of Rust
Two specific PCR primers Pm1-F (5′-AATTGTGGCTCGAACCATCTTC-3′)/Pm1-R (5′-TTGCTACTTTCCTTGATGCTC-3′) and Pk1-F (5′-AAGAGTGCACTTAATTGTGGCTC-3′)/Pk1-R (5′-CAGGTAACACCTTCCTTGATGTG-3′) were used to detect P. melanocephala and P. kuehnii, respectively [20]. Both primer pairs target the internal transcribed spacer (ITS1) region of fungal genomes of P. melanocephala and P. kuehnii. PCR amplification was performed using a VeritiPro thermal cycler (Thermo Fisher Scientific, Waltham, MA, USA). The total volume of 25 μL includes 1.0 μL DNA (about 100 ng) as the template, 1.0 U Ex Taq DNA polymerase, 2.5 μL 10 × Ex Taq buffer, 0.25 μM upstream and downstream primers, and 0.2 mM dNTPs. The reaction conditions were 94 °C for 5 min; 35 cycles of 94 °C for 30 s, 56 °C for 30 s, and 72 °C for 30 s; and 72 °C for 7 min. PCR amplification products were detected using 1.5% agarose gel electrophoresis.
2.3. Cloning and Sequencing of PCR Fragments
All PCR-positive products were purified and ligated into vector pMD19-T (TaKaRa, Dalian, China). Three positive clones per sample were picked up and sequenced with forward primer M13F (−47) and reverse primer M13F (−48). If the differences in nucleotide sequences at each site were simultaneously present two times in the same PCR amplifications, they were considered to be genetic variation. To determine the adaptive characteristics of P. kuehnii in China, the SNP analysis was performed on these PCR amplifications with PkPm-F/R primers in positive samples infected by P. kuehnii.
2.4. Multiple Sequence Alignment and Phylogenetic Analysis
A total of 62 sequences of P. melanocephala and 96 sequences of P. kuehnii were obtained from rust leaf samples with nucleotide lengths of 480 base pairs (bps) and 527 bps, respectively. In addition, some reference sequences of P. melanocephala (n = 11) and P. kuehnii (n = 39) from the United States, Argentina, Cuba, Australia, and Brazil were downloaded from GenBank (http://www.ncbi.nlm.nih.gov (accessed on 13 March 2024)). All sequence information is listed in Table S5. A phylogenetic tree was constructed using MEGA 11 software [21]. A phylogenetic tree was constructed using the neighbor-joining (NJ) method with default parameters in MEGA11. Robustness of the node of the phylogenetic tree was assessed from 1000 bootstrap replicates. Nucleotide sequence identity analysis was conducted using BioEdit 7.1.9 software [22].
2.5. Molecular Detection of Bru1 Gene
Four molecular markers, including R12H16-F/-R, 9O20-F4-F/-R, R12E03-1-F/-R, and R12E03-2-F/-R, were used to detect the Bru1 gene (Table S4). The PCR volume and procedures were performed as described by Wang et al. [14]. The PCR amplification products of R12H16 were directly detected using 1.5% agarose gel electrophoresis. For 9O20-F4/HaeIII, the PCR products were detected by HaeIII restriction enzyme digestion (Thermo Fisher Scientific, Waltham, MA, USA), and then the band polymorphisms were analyzed. The Bru1 gene was cloned and sequenced from sugarcane clones using four sets of molecular markers (R12H16, R12E03-1, 236 R12E03-2, and 9O20-F4). The SNP analysis was conducted on these Bru1 sequences.
2.6. Collection of Spores of P. melanocephala
The spores of P. melanocephala were collected in Dehong, Yunnan Province (China) in October 2018 and 2019, following the protocol described by Li et al. [11]. Briefly, sugarcane leaves with brown rust symptoms were immersed in a bucket of water for 2 h, and then were gently rubbed by hand to collect spores. Solution containing spores in the bucket was filtered through two layers of gauze. The filtrated suspension was used for artificial inoculation. The number of spores was calculated using a blood cell-counting plate to obtain a final concentration of 10^4^ CFU/mL for inoculation.
2.7. Inoculation Method for Brown Rust
The artificial inoculation for brown rust was carried out in the greenhouse based on the protocol described by Li et al. [11]. Namely, the spore suspension was evenly sprayed on sugarcane leaves before the drops flowing on leaves. Subsequently, the plants were thoroughly covered by transparent plastic bags to increase air humidity. A total of 88 clones comprising 4 E. arundinacus, 71 E. arundinaceus hybrids, and 13 parental clones (12 Saccharum spp. and no S. officinarum) were inoculated (Table S2). The varieties R570 and Q124 acted as the controls for resistance and susceptibility to brown rust, respectively. The plants were grown in a greenhouse at 28 °C with a relative humidity of 80% in October 2018 and 2019.
2.8. Experimental Designs to Identify Resistance to Brown Rust
Single-bud stalk cuttings of offspring of E. arundinaceus and their parents were cleaned by hot water treatment at 50 °C for 2 h. Then, they were planted in 40 × 35 cm pots filled with 2/3 soil and organic matter (3:1.5) in each pot. The five-leaf-stage plants were used for artificial inoculation. The experiment was based on randomized block design each year using three biological replicates, each consisting of three pots with six plants per pot for each variety. A spore suspension of P. melanocephala was inoculated on plant leaves, while the monk-inoculation of sterile water was used as blank control. Incidence ratios and disease severities were observed and recorded five times every five days post inoculation. The classification standard of disease severity for brown rust is grades 1–9 [11]. Disease index (%) was calculated using the following formula [8]:
PCR detection was conducted with Pm1-F/Pm1-R on some representative leaves to verify the P. melanocephala infection in each inoculated variety.
2.9. Data Analysis
A paired sample t-test was used to examine the statistically significant difference between disease indexes (the last time-point) among 88 clones of E. arundinaceus (HN92-77) hybrids and their parents together with 3 additional E arundinaceus clones between two years (2018 vs. 2019) (Table S3). A cluster analysis was performed with the Centetoid Method on disease indexes (the last time-point). All statistical analysis was performed using the SAS 9.2 software (https://support.sas.com (accessed on 20 January 2021)).
3. Results
3.1. Morphology of Rust Fungi in Sugarcane
To observe symptoms of rust and the morphology of casual fungi, representative leaf samples with rust symptoms were visually discriminated. The classical symptoms of brown rust were characterized by reddish-brown to brown lesions and sporulating pustules on the lower leaf surfaces (Figure 1A). In contrast, orange rust presented with orange to orange-brown lesions and pustules, lacking the darker brown pigmentation observed in brown rust. Notably, the pustules in orange rust formed clustered groupings on the lower leaf surfaces, distinguishing them from the more randomly distributed pustules associated with brown rust (Figure 1B). The morphologies of the urediniospores of brown rust were a dark color with no apical thickening (Figure 1C), while the urediniospores of orange rust were usually pear-shaped and orange to yellow-brown in color (Figure 1D).
3.2. Sugarcane Rust Detection Using PCR Method
To uncover the distribution and incidence of brown rust and orange rust in the main sugarcane planting provinces of China, a total 201 sugarcane samples was diagnosed by PCR with two primer pairs (Pm1-F/-R and Pk1-F/-R) for P. melanocephala and P. kuehnii, respectively (Figure S1). The sequencing results confirmed that these positive samples were indeed infected with one or both pathogenic fungi. Of the 201 samples, the infection rates of P. melanocephala and P. kuehnii were 17.9% and 34.8%, respectively, and the mixed infection rate was 12.9%. With the exception of samples from Fujian province, both P. melanocephala and P. kuehnii were detected in samples from all locations, including Guangdong, Guangxi, Yunnan, Hainan, Sichuan, and Guizhou province (Table 1).
3.3. Phylogenetic Analysis of P. melanocephala and P. kuehnii
To understand the phylogenetic relationships of each causal fungus causing sugarcane rust, two phylogenetic trees were constructed based on the nucleotide sequences of ITS1 for P. melanocephala (this study = 62 and GenBank = 11) and P. kuehnii (this study = 96 and GenBank = 39). The phylogenetic analysis showed that the sequences of P. melanocephala could be divided into two groups (groups I and II) (Figure 2A), whereas the sequences of P. kuehnii could be clustered into three groups (I, II, and III) (Figure 2B). The sequences of P. melanocephala from China were distributed in both groups (Figure 2A) while the sequences of P. kuehnii from China were clustered in Group I (Figure 2B). The sequence identity analysis indicated that the Chinese isolates of P. melanocephala had 98.7~100.0% sequence identities and shared 94.7–100.0% identities with the foreign isolates (GenBank No. PP555955–PP556016). Meanwhile, the Chinese isolates of P. kuehnii had 97.6–100.0% sequence identities and shared 93.9–100.0% identities with the foreign isolates (GenBank No. PP556017–PP556112).
3.4. Single Nucleotide Polymorphism (SNP) Analysis of P. kuehnii
To analyze the SNPs among P. kuehnii sequences, a total of 96 nucleotide sequences (606 bps) from PCR amplifications with the primers Pk1-F/Pk1-R were used for the SNP analysis. Among the 96 PCR fragments, 31, 22, and 43 sequences were adenine (183A), guanine (183G), or both bases (183A/G) at the 183rd nucleotide site, accounting for 32.3%, 22.9%, and 44.8%, respectively (Figure 3).
3.5. Detection of Bru1 Gene in E. arundinaceus Hybrids and Their Parents
Four sets of molecular markers (R2H16, 9O20-F4, R12E03-1, and R12E03-2) were used for screening whether the Bru1 gene was present in 88 E. arundinaceus hybrids and their parents. Of the 88 tested clones, 19 (21.6%) clones carrying Bru1 were found, including 13 offspring of E. arundinaceus and 6 backcross parents of the “ROC” series (Table 2 and Figure 4). Bru1 was also present in the resistant variety R570, but this gene was not detected in four E. arundinaceus varieties (Hainan92-79, Hainan92-91, Hainan92-77, and Hainan92-105), nor in S. officinarum (Badila) and their offspring (Table S2). Bru1 is absent in the susceptible variety Q124. Notably, the clone YCE06-127 (no. 43 in Table 2) did not harbor the Bru1 gene as revealed by 9O20-F4/RsaI (191 bp target band was absent after enzyme digestion) (Figure 4C), whereas the opposite result was gained based on 9O20-F4/HaeIII (389 bp target band was present after enzyme digestion) (Figure 4D).
3.6. SNP Analysis of Bru1 Gene
The SNP analysis was carried out on all sequences of the Bru1 gene generated from four molecular markers (R12H16, R12E03-1, R12E03-2, and 9O20-F4) in the 19 above-mentioned clones carrying this gene (Figure S2). Among the Bru1 sequences (570 bps) from the R12H16 marker, three clones (YCE06-24, YCE06-91, and YCE01-3) exhibited two SNP sites, 146T/C (thymine/cytosine) and 394C/G (cytosine/guanine). Among the Bru1 sequences (600 bps) from the R12E03-1 marker, four clones (YCE04-55, YCE06-127, YCE06-24, and YCE01-3) had two SNP sites, 323C/A (cytosine/adenine) and 519T/C. Among the Bru1 sequences (589 bps) from the R12E03-2 marker, three clones (YCE06-24, YCE01-3, and YCE06-127) exited three SNP sites, including 41C/A, 369A/G (adenine/guanine), and 543A/G, while the clone YCE04-55 was present in two SNP sites including 269A/G and 301T/C (thymine/cytosine). Among the Bru1 sequences (200 bps) from the 9O20-F4/HaeIII marker, four clones (YCE06-166, YCE06-24, YCE06-91, and YCE01-3) had three SNP sites, including 159T/C, 189A/G, and 230T/C. Notably, no SNP sites were found in seven clones (YCE02-179, YCE03-168, YCE03-167, YCE03-405, YCE06-61, YCE06-63, and YCE07-56) and six “ROC” clones (Figure S2).
3.7. Phylogenetic Analysis of Bru1 Gene
To explore the phylogenetic relationship of Bru1 among 13 offspring of E. arundinaceus and 6 “ROC” varieties, a phylogenetic tree was constructed based on sequences generated from the molecular marker 9O20-F4 together with 65 according to sequences reported in the study by Wang et al. [14] (Figure 5). The 13 offspring of E. arundinaceus were clustered with clones of Saccharum, including NG77-004 (S. robustum), Youba_1 (S. sinense), Songxi-zhuzhe_1 (S. sinense), Katha_1 (S. barberi), R570 (cultivated species), and ROC22 (Saccharum spp.). The clone FJ-Banmao of E. arundinaceus was assigned into a unique evolutionary branch and the clone LA Purple of S. officinarum and the clones of S. spontaneum were clustered into a large evolutionary branch.
3.8. Identification of Resistance to Brown Rust in E. arundinaceus Hybrids and Their Parents
Resistance identification of brown rust was performed in 88 E. arundinaceus hybrids and their parents plus 2 control clones (R570 resistant and Q124 susceptible to brown rust) in October 2018 and 2019. The typical symptoms of brown rust were developed among clones after being inoculated by spores of P. melanocephala (Figure S3). These inoculated samples showing rust were controlled for the presence of P. melanocphala by PCR with primers Pm1-F/Pm1R. No significant difference was found in the disease indexes of these tested varieties between the two years (p = 0.992) (Table S2). Thus, the average disease indexes among 88 tested clones varied from 0 to 67.9%. Among them, 69.3% (61/88), 10.2% (9/88), and 9.1% (8/88) clones showed high resistance, resistance, and medium resistance to brown rust, respectively. Ten clones (11.4%) showed susceptibility to brown rust (Table S2).
Based on the average disease indexes, resistance identification was conducted in the tested clones. Four categories were proposed (Table S2): (1) the control clone Q124 showed high susceptibility to brown rust (disease index = 67.9%); (2) 10 clones were susceptible to brown rust with disease indexes ranging from 30.6% (YCE03-218) to 36.4% (YCE01-102); (3) 16 clones were resistant to brown rust with disease indexes ranging from 5.8% (YCE01-92) to 22.9% (YCE01-61); and (4) the remaining 62 clones, including the resistant control clone R570, were highly resistant to brown rust (disease index = 0). The cluster analysis further supported the grouping by disease indexes (Figure 6).
4. Discussion
In China, the emergence of sugarcane rust occurred in the dominant cultivar F176 in Taiwan in the 1970s. The prevalence of sugarcane rust is strongly correlated with cultivar resistance, therefore making the development and deployment of resistant varieties the most economically effective strategy for controlling brown rust epidemics [7,10]. This study extensively assessed the prevalence of brown rust and orange rust in sugarcane in seven sugarcane planting provinces in China. Our results suggest that the two sugarcane rust diseases are widely disseminated in all surveyed provinces with single or mixed infections. Overall, orange rust caused by P. kuehnii is more prevalent than brown rust caused by P. melanocephala in China. P. kuehnii is a predominant fungus in the largest sugarcane planting area (Guangxi province), but P. melanocephala is a predominant pathogen in the second largest sugarcane planting area (Yunnan province). Similar results have been reported in previous studies [7,8]. Orange rust is an emerging major disease in some specific provinces, particularly in Guangxi, China.
The evolutionary arms race between plants and pathogens is a sophisticated process. Usually, novel virulent races that break through plant resistance genes often trigger rapid disease epidemics, therefore it is necessary to continuously monitor pathogenic variability in agricultural systems. Two races (1040 and 2143) of P. kuehnii occurred in Florida [5] and pathogenic variants of this pathogen (races PkSp01-01a4 and PkSp01-01) have also been identified in Brazil [23,24]. Four pathogenic races of P. melanocephala occurred in Florida [25], but there was no significant variation in the virulence of this causal agent in Brazil [26]. Notably, the variability in pathogenicity in P. melanocephala was associated with host genotypes [27]. Here, three phylogenetic groups of P. kuehnii and two groups of P. melanocephala were found in China. Although molecular evidence showed the occurrence of several SNPs in P. kuehnii and P. melanocephala, there is still a need for more in-depth investigations of the pathogenic variation in the two causal pathogens. Some SNPs present in P. kuehnii may be one of the main reasons for the prevalence of orange rust in various sugarcane areas in China [20]. Additionally, non-synonymous mutations or InDels in fungal genomic loci are possibly associated with the pathogenicity of pathogens, contributing to breakthroughs in the rust resistance of sugarcane breeding [20,28]. However, more biological experiments need to be explored in this inference.
Identification of resistance genes or loci is an effective and durable way to regenerate resistance varieties in sugarcane breeding programs. Some molecular markers, such as Bru1 and G1, are closely linked to brown rust and orange rust-resistance genes in sugarcane, respectively [16,29]. The efficiency of the G1 marker for predicting rust resistance was higher than that of the Bru1 marker [30]. Numerous investigations showed that the Bru1 gene is extensively present in the Saccharum complex, including wild germplasms [11], modern sugarcane cultivars [10,13,14], and sugarcane accessions in Brazil [30] and worldwide [19]. In this study, the tested clones of E. arundinaceus were highly resistant to brown rust, but they did not contain the Bru1 gene, which indicated that some alternative resistant genetic sites or genes possibly exist in these E. arundinaceus clones. Additionally, the interaction between sugarcane and races should be considered because the divergence in P. melanocephala pathogenicity was related to sugarcane genotypes [27].
It is noteworthy that some main sugarcane cultivars (LC05-136, GT42, and GT44) being planted in China are susceptible to sugarcane rust and they do not carry the Bru1 gene [10]. Therefore, mining and the application of novel resistant gene sources from other wild germplasm resources in the Saccharum complex, such as E. arundinaceus, is a crucial strategy for improving sugarcane disease resistance in modern sugarcane breeding programs. In addition, there is an urgent need to develop more robust and accurate molecular markers for resistance identification in sugarcane rust, especially orange rust, which will speed up the sugarcane breeding process.
5. Conclusions
The application of resistant cultivars developed by breeding programs is an effective and efficient strategy for preventing and controlling sugarcane rust. In this study, artificial inoculation identification and molecular detection were conducted to determine the resistance to brown rust in E. arundinaceus hybrids and their parents. Most of the test clones showed resistance to brown rust, but only 24.4% of the resistant varieties harbored the Bru1 gene. The Bru1 gene in the offspring might be offered by “ROC” varieties. E. arundinaceus germplasm resources exhibit resistance to rust, but no Bru1 gene was detected in these clones. These results suggest that other rust resistance genes might be present in the germplasms of E. arundinaceus. More in-depth investigations are needed to explore novel genes or genetic sites for rust resistance, which will help overcome the limitation of only relying on a single source of resistance. Notably, mining and the utilization of other disease-resistant genes from the Saccharum complex is an alternative strategy in genetic improvement programs of sugarcane.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Cárdenas D.E. Carrillo-Tarazona Y. Sood S.G. Hincapie M.A. Wang J. Rott P.C. Cano L.M. A diagnostic guide for orange rust disease in sugarcane Plant Health Prog.20242551452610.1094/PHP-02-24-0013-DG · doi ↗
- 2Viswanathan R. Degeneration in sugarcane varieties: Does the Sugar Industry realize it?Sugar Tech.2024261501150410.1007/s 12355-024-01439-4 · doi ↗
- 3Comstock J. Shine Jr J. Raid R. Effect of rust on sugarcane growth and biomass Plant Dis.19927617517710.1094/PD-76-0175 · doi ↗
- 4Sanjel S. Chaulagain B. Small I.M. Comstock J.C. Hincapie M. Raid R.N. Rott P. Comparison of progress of brown rust and orange rust and conditions conducive for severe epidemic development during the sugarcane crop season in Florida Plant Dis.201910382583110.1094/PDIS-05-18-0862-RE 30806575 · doi ↗ · pubmed ↗
- 5Sanjel S. Hincapie M. Wang Y. Todd J. Chaulagain B. Sood S. Comstock J.C. Raid R.N. Rott P. Occurrence of two races of Puccinia kuehnii causing orange rust of sugarcane in Florida Plant Pathol.2021701616162510.1111/ppa.13405 · doi ↗
- 6Rott P. Bailey R.A. Comstock J.C. Croft B.J. Saumtally A.S. A Guide to Sugarcane Diseases CIRAD/ISSCT Montpellier, France 200085–89, 121–125
- 7Fu H.Y. Xiao S.H. Liu Y.H. Sun S.R. Wu X.B. Chen R.K. Gao S.J. Molecular detection of causal pathogens causing rust disease and Bru 1 resistance-gene in elite Sugarcane Clones Chin. J. Trop. Crops 201637958963
- 8Li W.F. Wang X.Y. Shan H.L. Zhang R.Y. Li Y.H. Lu W.J. Huang Y.K. Occurrence and damage of epidemic fungal diseases in middle and late stages of sugarcane growth in Yunnan Province of China Eur. J. Plant Pathol.202216435336410.1007/s 10658-022-02566-y · doi ↗