Sulfurimonas microaerophilic sp. nov. and Sulfurimonas diazotrophicus sp. nov.: Two Novel Nitrogen-Fixing and Hydrogen- and Sulfur-Oxidizing Chemolithoautotrophs Within the Campylobacteria Isolated from Mangrove Sediments
Yangsheng Zhong, Yufei Li, Zhaodi Wang, Liang Cui, Shiwei Lv, Han Zhu, Qing Yuan, Qiliang Lai, Shasha Wang, Lijing Jiang

TL;DR
Two new species of bacteria that can fix nitrogen and use hydrogen and sulfur for energy were discovered in mangrove sediments and classified as Sulfurimonas microaerophilic and Sulfurimonas diazotrophicus.
Contribution
The discovery and classification of two novel nitrogen-fixing Sulfurimonas species with unique metabolic and genomic traits.
Findings
Strains HSL1-7T and HSL3-1T are obligate chemolithoautotrophs that use hydrogen and sulfur compounds for energy.
Both strains are strongly diazotrophic, capable of fixing nitrogen in the absence of a nitrogen source.
Genomic and phylogenetic analysis confirmed they represent two distinct new species within the genus Sulfurimonas.
Abstract
Two novel marine hydrogen- and sulfur-oxidizing bacteria, designated HSL1-7T and HSL3-1T, were isolated from mangrove sediments from Fujian Province, China. Strain HSL1-7T exhibited Gram-negative, rod-shaped to slightly curved morphology with polar flagellum-driven motility, whereas strain HSL3-1T was Gram-negative, rod-shaped and non-motile. Strain HSL1-7T and HSL3-1T were obligate chemolithoautotrophs, capable of using molecular hydrogen and thiosulfate as an energy source, and molecular oxygen and elemental sulfur as the electron acceptors for growth. Cellular fatty acid profiles revealed similar predominant components (C16:1ω7c, C16:0, C18:1ω7c, and C14:0) in both strains. Strains HSL1-7T and HSL3-1T were strongly diazotrophic, as demonstrated by 15N2 fixation when a fixed nitrogen source was absent from the growth medium. The DNA G+C contents of strains HSL1-7T and HSL3-1T were…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
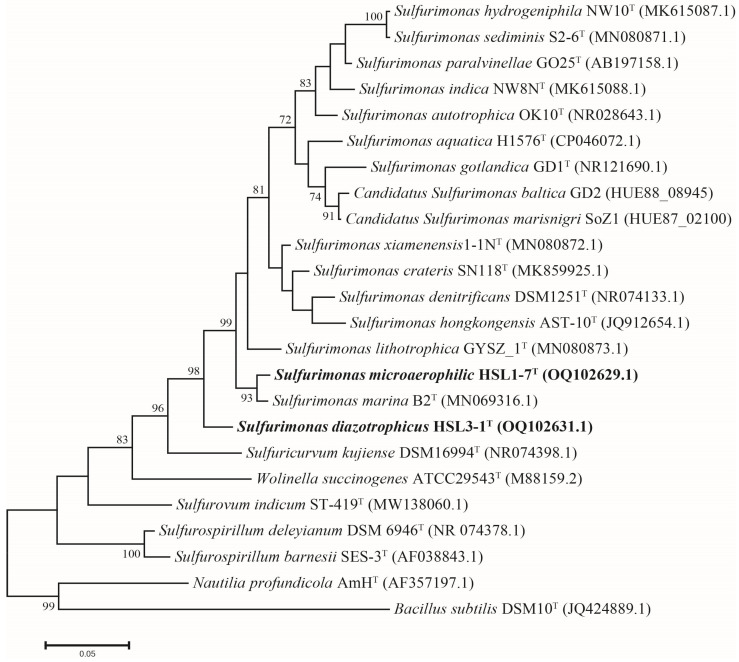 Figure 1
Figure 1 Figure 2
Figure 2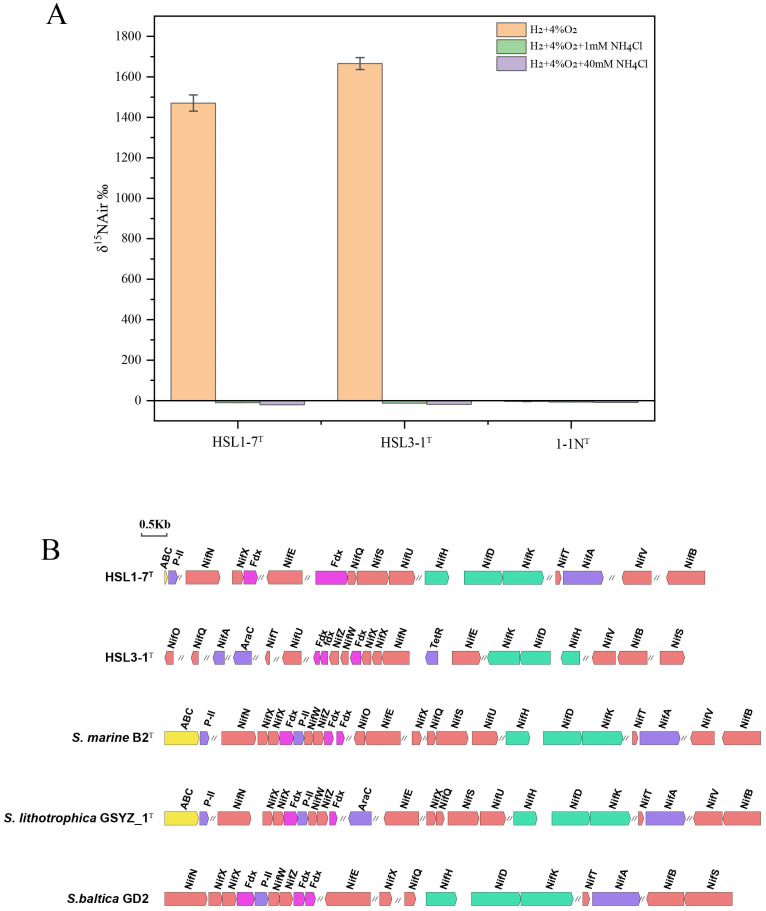 Figure 3
Figure 3- —National Natural Science Foundation of China
- —Scientific Research Foundation of Third Institute of Oceanography
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenomics and Phylogenetic Studies · Microbial Community Ecology and Physiology · Methane Hydrates and Related Phenomena
1. Introduction
Mangrove sediments are typically recognized as organic-rich but nitrogen-limited ecosystems [1,2]. As the rate-limiting step of nitrogen cycling, nitrogen fixation is particularly important to alleviate the nitrogen limitation of mangrove ecosystems. Our recent studies have demonstrated that chemolithoautotrophs, such as members of Campylobacterota, Proteobacteria, and Nitrospirota, are the dominant nitrogen fixers in mangrove sediments, rather than heterotrophs [3]. This study underscores the significance of chemolithoautotrophs in carbon-dominant ecosystems. Compared with heterotrophic nitrogen-fixing bacteria [4], fewer chemolithoautotrophic diazotrophs have been cultured so far. Therefore, isolating new related strains is conducive to deepening our understanding of the dark nitrogen fixation of this group in mangrove sediments.
The genus Sulfurimonas, belonging to the class Campylobacteria (formerly Epsilonproteobacteria) within the phylum of Campylobacterota (formerly Proteobacteria), was first proposed by Inagaki et al. in 2003 [5,6,7]. Members of the genus Sulfurimonas are globally distributed in sulfidic environments, including coastal sediments, pelagic redoxclines, deep-sea hydrothermal vents, and terrestrial soils [7,8]. They play an important role in the marine carbon, nitrogen, and sulfur cycles. The most widely shared feature of species of the genus Sulfurimonas is chemolithoautotrophic growth. A variety of electron donors and acceptors, such as molecular hydrogen (H_2_) and reduced sulfur compounds, are commonly utilized as the electron donors, and molecular oxygen (O_2_), nitrate (NO_3_^−^), and elemental sulfur as the common electron acceptors. This may contribute to their widespread distribution [9,10,11]. The unique feature of sulfur-oxidizing coupled denitrification enables its application in wastewater denitrification [12], thereby facilitating the development of novel environmental remediation technologies.
At the time of writing, the genus Sulfurimonas comprises ten species with validly published names and three Candidatus species; these can be found in an up-to-date website (https://lpsn.dsmz.de/genus/sulfurimonas accessed on 1 November 2024). Among these, five species were isolated from deep-sea hydrothermal vents environments, including Sulfurimonas indica NW8N^T^, Sulfurimonas autotrophica OK10^T^, Sulfurimonas hydrogeniphila NW10^T^, Sulfurimonas paralvinellae GO25^T^, and Sulfurimonas sediminis S2-6^T^ [7,13,14,15,16]. Four species were isolated from coastal marine sediments, including Sulfurimonas xiamenensis 1-1N^T^, Sulfurimonas denitrificans DSM 1251^T^, Sulfurimonas hongkongensis AST-10^T^, and Sulfurimonas lithotrophica GYSZ_1^T^ [17,18,19]. Sulfurimonas marina B2^T^ was isolated from a deep-sea sediment sample. Sulfurimonas crateris SN118^T^ was isolated from a terrestrial mud volcano, and Sulfurimonas aquatica H1576^T^ was isolated from a brackish lake, while Sulfurimonas gotlandica GD1^T^, Candidatus Sulfurimonas baltica GD2, and Candidatus Sulfurimonas marisnigri SoZ1 were isolated from the pelagic redoxcline of the Baltic Sea and Black Sea [20,21,22,23,24].
In this study, we report two novel strains, designated HSL1-7^T^ and HSL3-1^T^, isolated from mangrove sediments sampled at the Jiulong River estuary, Fujian Province, China. Their physiological properties and an 16S rRNA gene sequence analysis indicate that strains HSL1-7^T^ and HSL3-1^T^ represent members of the genus Sulfurimonas. A comparative genomic analyses indicates that they exhibit diverse metabolic profiles, especially their unique nitrogen fixation capacity, which enables us to further elucidate their potential ecological functions within mangrove ecosystems.
2. Materials and Methods
2.1. Isolation of the Novel Strain and Cultivation
Sediment samples were collected from Zhangzhou mangroves in the Jiulong River estuary, Fujian Province, China (24°23’33” N, 117°54’11” E), and transported to the laboratory at 4 °C. For microbial enrichment, approximately 1 g of fresh sediment was aseptically inoculated into the MMJHS medium, followed by incubation at 28 °C for 7 days under a gas mixture of 76% H_2_/20% CO_2_/4% O_2_ (200 kPa) according to established protocols [25]. The MMJHS medium consisted of NaCl (30 g l^−1^), NH_4_Cl (0.25 g l^−1^), KCl (0.33 g l^−1^), CaCl_2_·2H_2_O (0.14 g l^−1^), MgCl_2_·6H_2_O (4.18 g l^−1^), K_2_HPO_4_ (0.14 g l^−1^), NaHCO_3_ (1 g l^−1^), Na_2_S_2_O_3_·5H_2_O (10 mM), Wolfe’s vitamins (1 mL l^−1^), and a trace element solution (10 mL l^−1^) [26]. After successful enrichment with the MMJHS medium, strains HSL1-7^T^ and HSL3-1^T^ were successfully isolated as axenic cultures using the dilution-to-extinction method [27]. The purity of the cultures was confirmed by microscopic examination coupled with a full-length 16S rRNA gene sequence analysis.
2.2. Phenotypic and Chemotaxonomic Analyses
Gram staining was performed following the manufacture’s protocol (Hangzhou Tianhe Microorganism Reagent, Hangzhou, China) using the standard Schaeffer-Fulton method with quality control strains included for validation. The cell morphology was observed using the cells grown in the MMJHS liquid medium at 32 °C for one day, using a transmission electron microscopy (TEM, JEM-1230, JEOL, Tokyo, Japan). The growth of strains HSL1-7^T^ and HSL3-1^T^ was measured via direct cell counting under a phase-contrast microscope using a hemocytometer. To determine the physiological characteristics of both strains, triplicate cultures grown in the MMJHS medium were analyzed under each experimental condition.
The growth temperature was determined by incubating the cultures in the MMJHS medium for 24 h across a temperature gradient from 4 to 55 °C (4, 10, 15, 20, 25, 28, 30, 35, 37, 40, 45, 50, and 55 °C). The salinity tolerance was assessed using NaCl concentrations ranging from 0 to 9.0% (w/v) in 0.5% increments. For pH profiling, cultures in the MMJHS medium were incubated for 24 h across pH 4.0–9.0 (0.5 pH unit intervals) with specific buffer systems: acetate/acetic acid (pH 4.0–5.0), MES (pH 5.0–6.0), PIPES (pH 6.0–7.0), HEPES (pH 7.0–7.5), and Tris/CAPSO (pH 8.0 and above) [28]. Oxygen requirements were evaluated under controlled atmospheres with O_2_ concentrations from 0 to 20% (0, 1, 2, 4, 6, 8, 10, 12, 15, and 20% at 200 kPa). In the case of oxygen absence, 10 mM nitrate was added as a potential electron acceptor.
The utilization of alternative electron donors was evaluated by substituting sulfite (5 mM), thiocyanate (5 mM), tetrathionate (5 mM), elemental sulfur (1% w/v), or sodium sulfide (50–100 μM), for thiosulfate in the MMJS medium with a gas phase of 76% N_2_/20% CO_2_/4% O_2_ (200 kPa). The molecular hydrogen was also examined in the MMJH medium in the absence of thiosulfate under a gas phase of 76% H_2_/20% CO_2_/4% O_2_ (200 kPa). For the electron acceptor analysis, nitrate (10 mM), nitrite (5 mM), ferric citrate (20 mM), ferrihydrite (20 mM), and manganese (IV) oxide (200 μM) were tested with thiosulfate as the sole electron donor under 80% N_2_/20% CO_2_ (200 kPa).
The metabolic capability of inorganic nitrogen sources was evaluated by supplementing the nitrogen-deprived MMJHS medium (lacking all nitrogen sources) with ammonium chloride (5 mM), sodium nitrate (5 mM), or sodium nitrite (5 mM). Cultures were incubated under a gas phase of 76% H_2_/20% CO_2_/4% O_2_ (200 kPa). Utilization of N_2_ was also tested under a 76% N_2_/20% CO_2_/4% O_2_ (200 kPa) gas phase. Potential nutrients required for growth, such as selenite, tungstate, and vitamins, were examined with the MMJHS medium with, and without, the specified nutrients.
The heterotrophic growth capability was investigated in the MMJHS medium by substituting NaHCO_3_ with various organic carbon sources under a gas phase of 96% N_2_/4% O_2_ (200 kPa). To determine whether there was culture after 24 h of incubation, direct cell counting using a hemocytometer was carried out. These organic carbon sources included: 0.1% (w/v) peptone, starch, tryptone, casein, yeast extract, and casamino acids; 5 mM acetate, citrate, formate, succinate, propionate, tartrate and pyruvate; 5 mM of each of 20 amino acids; and 0.02% (w/v) sucrose, galactose, glucose, fructose, lactose, maltose, and trehalose. To systematically identify the alternative energy sources, these organic compounds were used as an energy source in the MMHJS medium to replace thiosulfate under a gas phase of 76% N_2_/20% CO_2_/4% O_2_ (200 kPa).
The whole-cell fatty acid profiles were analyzed using the MIDI Sherlock Microbial Identification System (v 6.0B). Cultures grown in the MMJHS medium (32 °C, 24 h) underwent sequential saponification, extraction, and methylation. Fatty acid methyl esters (FAMEs) were separated via an Agilent 6850 gas chromatograph and identified against the TSBA 6.0 reference database [29].
2.3. Phylogeny Analysis Based on the 16S rRNA Gene Sequences
The genomic DNA isolation from strains HSL1-7^T^ and HSL3-1^T^ was performed with a commercial bacterial DNA extraction kit (SBS Genetech Co., Ltd., Shanghai, China) following the manufacturer’s protocol. The DNA purity and concentrations were determined using a NanoDrop 2000 spectrophotometer (Thermo Scientific, Waltham, MA, USA). The amplification of the full-length 16S rRNA genes was achieved with the universal primer pair 27F (5′-AGAGTTTGATCMTGGCTCAG-3′) and 1492R (5′-GGTTACCTTGTTACGACTT-3′), as previously described [30]. The 16S rRNA genes were sequenced via the Sanger method (Applied Biosystems^TM^ 3730XL, Waltham, MA, USA) by Sangon Biotech (Shanghai) Co., Ltd., Shanghai, China. The full-length 16S rRNA gene sequencing was performed using the EzTaxon-e server [31]. The sequences of related taxa were obtained from the GenBank database. The phylogenetic reconstruction was conducted in MEGA 7.0 [32] employing three distinct algorithmic approaches: neighbor-joining [33], maximum-likelihood [34], and minimum evolution [35]. The multiple alignment was first performed using CLUSTAL_W, followed by an evolutionary distance calculation through Kimura’s two-parameter model [36]; the bootstrap values were determined based on 1000 replications [37].
2.4. Genome Analysis and Functional Annotation
The complete genome of strains HSL1-7^T^ and HSL3-1^T^ was sequenced by Shanghai Majorbio Bio-Pharm Technology Co., Ltd. (Shanghai, China) on a Pacific Biosciences (PacBio, Menlo Park, CA, USA) sequencing platform using a single-molecule real-time (SMRT) technology. The sequenced reads were filtered and the high-quality paired-end reads were assembled to reconstruct the circulating genome using SOAPdenovo2 software (v 2.0) [38]. The genomic DNA G+C content was determined from the complete genome sequences. To identify tRNA and rRNA within the genome, tRNAscan-SE and rRNAmmer were utilized [39]. The complete 16S rRNA gene sequence was extracted from the whole genome using RNAmmer [40]. The gene prediction was performed using Glimmer (v 3.02) [41]. The genome annotation was carried out using RAST server [42], METABOLIC (v 4.0) [43] and KofamtKOALA [44] against the KEGG database. An orthoANI algorithm [45] was implemented to calculate the average nucleotide identity (ANI) values between the paired genomes. The digital DNA-DNA hybridization (dDDH) values were estimated through the Genome-to-Genome Distance Calculator (GGDC) platform (https://ggdc.dsmz.de/, accessed on 5 December 2024). The phylogenomic tree was reconstructed with the Up-to-date Bacterial Core Gene (UBCG) pipeline (v 3.0) [46]. The phylogenomic tree was visualized and decorated using ChiPlot (https://www.chiplot.online/, accessed on 25 December 2024) [47].
2.5. Characterization of N2 Fixation
The nitrogen-fixing ability of isolates in this study were determined using the ^15^N_2_ assimilation method, as previously described [3]. Cells were grown microaerobically in a 60 mL serum bottle containing 10 mL of the nitrogen-free MMJHS medium, under a gas phase of 70% H_2_/20% CO_2_/4% O_2_ (200 kPa). 10 mL of ^15^N_2_ (99% atomic ^15^N; Isotec) was added to the growth medium. The nitrogen fixation ability was also tested by the addition of inorganic nitrogen compounds including 1 mM or 40 mM NH_4_Cl. In addition, the strain Sulfurimonas xiamenensis 1-1N^T^, isolated from the coastal sediments of Xiamen Island, Fujian Province, China, was used as the control; this lacked nitrogen fixation gene clusters. All the cultures were incubated at 32 °C in the dark for 24 h, and subsequently harvested by centrifugation (10,000× g, 4 °C, 20 min) when visible turbidity was observed. The cells were then washed twice with ice-cold 20 mM Tris buffer prepared in artificial seawater, followed by freeze-drying overnight [48]. The ^15^N atomic percentage of freeze-dried cells was determined by a CarloErba elemental analyzer (Model NA 1500, Fisons Instruments, San Carlos, CA, USA) couple with a Finnegan MAT (ThermoQuest, Temecula, CA, USA) Delta S isotope ratio mass spectrometer [49].
3. Results and Discussion
3.1. Morphological and Phenotypic Characteristics
The cells of the strain HSL1-7^T^ were Gram-stain-negative, rods to slightly curved, with dimensions of approximately 1.8–2.3 μm in length and 0.4–0.6 μm in width and exhibited motility via a single polar flagellum (Figure S1a). The cells of strain HSL3-1^T^ were Gram-stain-negative, rod-shaped, with dimensions of approximately 0.7–2.5 μm in length and 0.2–0.5 μm in width, and non-motile with no flagellum observed in the MMJHS medium (Figure S1b). Strain HSL1-7^T^ exhibited growth over a temperature range of 4 °C to 40 °C (optimum 32 °C). It also grew in NaCl concentrations from 1.0 to 4.0% (w/v) (optimum 3.0%). The pH range for growth was 5.0 to 8.5 (optimum pH 7.0). Additionally, growth was observed in oxygen concentrations ranging from 1% to 10% (optimum 4% O_2_). No growth was observed without oxygen, or with over 15% in the headspace gas (Table 1). Growth of strain HSL3-1^T^ was observed at temperatures ranging from 4 °C to 45 °C (optimum 37 °C), at NaCl concentrations ranging from 2.0–4.0% (w/v) (optimum 3.0%), at a pH range of 5.0–8.5 (optimum pH 7.0), and at an O_2_ range of 0–20% (optimum 6% O_2_). Strains HSL1-7^T^ and HSL3-1^T^ both exhibit morphological and phenotypic similarities with the other species in the Sulfurimonas genus, including growth temperature range, pH range, and maximum O_2_ concentration. However, there were some differential features between strains HSL1-7^T^ and HSL3-1^T^ and the other reference strains, as shown in Table 1.
Physiological characterization through the chemoautotrophic cultivation experiments revealed that strains HSL1-7^T^ and HSL3-1^T^ could grow with molecular hydrogen and thiosulfate as the energy sources, while utilizing molecular oxygen and elemental sulfur as electron acceptors. Nitrate and nitrite could not be utilized as electron acceptors. The results of the inorganic nitrogen source tests indicated that strains HSL1-7^T^ and HSL3-1^T^ could grow with ammonium and molecular nitrogen as the nitrogen source. Nitrite, tungsten, selenium, and vitamin supplementation were not required for growth. The results of the heterotrophic growth tests indicated that none of the organic compounds sustained the growth of these two strains as the carbon or energy source. These results indicate that strains HSL1-7^T^ and HSL3-1^T^ are obligate chemolithoautotrophs.
3.2. Chemotaxonomy
To analyze the cellular fatty acid, strains HSL1-7^T^ and HSL3-1^T^, and the closely related species, were cultivated in the MMJHS medium at 32 °C for 24 h. The fatty acid compositions of strains HSL1-7^T^ and HSL3-1^T^ and the closely related type species of the genus Sulfurimonas are listed in Table 2. The major cellular fatty acids of strains HSL1-7^T^ and HSL3-1^T^ were C_16:1_ω7c, C_16:0_, C_18:1_ω7c, and C_14:0_, which are similar to those of S. marina B2^T^, S. lithotrophica GYSZ_1^T^, and S. xiamenensis 1-1N^T^. However, differences were observed in the relative abundance of some fatty acids between strains HSL1-7^T^ and S. marina B2^T^. For example, the fatty acid C_16:1_ω5c was observed at 0.6% in HSL1-7^T^ but was not found in S. marina B2^T^. The fatty acid C_16:1_ω7c was observed at 28.7% in HSL1-7^T^, while it was observed at 37.7% in S. marina B2^T^. Some differences in fatty acids profiles were also observed between strains HSL3-1^T^ and S. marina B2^T^; the fatty acid C_12:0_ was observed at 4.0% in HSL3-1^T^, while it was only observed at 1.3% in S. marina B2^T^. The fatty acid C_16:1_ω5c was observed at 0.4% in HSL3-1^T^ but not found in S. marina B2^T^ (Table 2). In addition, there were some differences between strains HSL1-7^T^ and HSL3-1^T^. The fatty acid C_12:0_ was observed at 0.3% in HSL1-7^T^, while it was observed at 4.0% in HSL3-1^T^. The fatty acid of C_18:0_ was observed at 2.6% in HSL3-1^T^ but was observed at 0.7% in HSL1-7^T^ (Table 2).
3.3. Phylogeny of 16S rRNA Gene Sequences
The comparative analysis of the full-length 16S rRNA gene sequences (1450bp) revealed that strain HSL1-7^T^ shares the closest phylogenetic affiliation with S. marina B2^T^, exhibiting a 98.45% sequence identity; the other strains shared sequence similarities below 95.5%. Strain HSL3-1^T^ was also most closely related to S. marina B2^T^ with a 94.45% sequence identity; the other strains shared sequence similarities below 94.0%. These sequence similarities were lower than the threshold of 98.65% for species delineation [50]. The phylogenetic analysis using the maximum-likelihood method revealed that strain HSL1-7^T^ was clustered with S. marina B2^T^, and strain HSL3-1^T^ was clustered with the other reference strains of Sulfurimonas (Figure 1). The phylogenetic topology was further supported by neighbor-joining and minimum-evolution analyses, which indicates that strains HSL1-7^T^ and HSL3-1^T^ represent members of the genus Sulfurimonas (Figures S2 and S3).
3.4. Genomic Properties
The genome size of strains HSL1-7^T^ and HSL3-1^T^ were 2.28 Mb and 2.49 Mb, respectively. The G+C contents of the genomic DNA of strains HSL1-7^T^ and HSL3-1^T^ were determined to be 36.1% and 57.3%, respectively (Table 1). The pairwise ANI values between HSL1-7^T^ and the closest related type strain S. marina B2^T^ was 89.2%, and the dDDH values between them were 36.9% (Table S1). The pairwise ANI values between strain HSL3-1^T^ and the closest related type strain S. marina B2^T^ were 67.41%, and the dDDH values between them were 23.7% (Table S2). All these values fell well below the established thresholds for prokaryotic species delineation (95–96% ANI and 70% dDDH) [51,52]. These data indicate that strains HSL1-7^T^ and HSL3-1^T^ represent two novel species of the genus Sulfurimonas.
The phylogenetic tree based on the 92 core gene sequences revealed that strain HSL1-7^T^ formed a distinct branch with S. marina B2^T^, while HSL3-1^T^ clustered together with other reference strains of Sulfurimonas (Figure 2). This result corroborated phylogenetic analyses of the 16S rRNA gene sequences, which supports that strains HSL1-7^T^ and HSL3-1^T^ represent members of the genus Sulfurimonas. Therefore, the genome-based phylogeny clearly indicates that strains HSL1-7^T^ and HSL3-1^T^ represent two novel species of the genus Sulfurimonas.
3.5. Genomic Functional Analysis
3.5.1. Carbon Metabolism
The genome analysis revealed that strains HSL1-7^T^ and HSL3-1^T^ possess the complete set of enzymes required for carbon fixation via the reductive tricarboxylic acid (rTCA) cycle, including the key enzymes encoding ATP citrate lyase (AclAB), 2-oxoglutarate: ferredoxin oxidoreductase (OorABCD), and pyruvate:ferredoxin oxidoreductase (PorABCD) (Figure 2). This genome characteristic was accordant with the metabolic features of chemoautotrophic growth in strains HSL1-7^T^ and HSL3-1^T^, which was also conserved in the Sulfurimonas. In addition, the genomes of strains HSL1-7^T^ and HSL3-1^T^ contained enzymes for the intact tricarboxylic acid (TCA) cycle, including aconitehydratase (Acn), 2-oxoglutarateoxidoreductase (KorABGD), and succinyl-CoAsynthetase (SucCD) (Figure 2). Emerging evidence reveals the TCA cycle operates bidirectionally as a metabolic orchestrator, not only fueling oxidative phosphorylation but dynamically supplying core metabolic nodes (α-ketoglutarate, succinyl-CoA, and oxaloacetate) which coordinate anabolic flux partitioning through moonlighting enzyme-mediated substrate channeling [53].
3.5.2. Hydrogen Metabolism
The genomic analysis revealed that strain HSL1-7^T^ contained one type of [Ni-Fe]-hydrogenase (Group I), while strain HSL3-1^T^ contained three types of [Ni-Fe]-hydrogenases (Group I, Group II, and Group IV) (Figure 2). Group I hydrogenases are membrane-bound enzymes that catalyze the respiratory oxidation of hydrogen, coupled to the reduction of electron acceptors [54]. Like most members of the genus Sulfurimonas, strains HSL1-7^T^ and HSL3-1^T^ both harbor two copies of Group I [Ni-Fe]-hydrogenase. This suggests that Group I hydrogenases might be essential for growth, like the genus Sulfurimonas. Group II hydrogenases are possibly involved in hydrogen sensing or energy conversion at low hydrogen concentrations [30]; this was only found in strain HSL3-1^T^ and some isolates from the seawater and sediments of the genus Sulfurimonas. Moreover, only strain HSL3-1^T^ contained Group IV hydrogenases Ech. However, neither strain HSL1-7^T^ nor HSL3-1^T^ contained Group IV hydrogenases Hyc and Coo (Figure 2).
3.5.3. Sulfur Metabolism
The genomic analysis revealed that strains HSL1-7^T^ and HSL3-1^T^ harbor two distinct Sox gene clusters (soxABXY_1_Z_1_ and soxCDY_2_Z_2_), which collectively constitute a complete Sox multienzyme system (Figure 2). While in the Sulfurimonas, only strains Sulfurimonas xiamenensis and Sulfurimonas lithotrophica had the soxCDY_2_Z_2_ gene cluster [55]. Thus, the complete Sox gene clusters confer the capability of the isolated strains to oxidize thiosulfate to sulfate. Moreover, the genomes of strains HSL1-7^T^ and HSL3-1^T^ also contained flavocytochrome c sulfide dehydrogenase (Fcc) and sulfide:quinone oxidizing reductase (Sqr) for sulfide oxidation (Figure 2), which are also found in all members of genus Sulfurimonas. Only strain HSL3-1^T^ contained the complete enzymes required for the assimilatory sulfate reduction pathway, including the key enzymes sulfate adenylyltransferase (CysDN), adenylylsulfate kinase (CysC), phosphoadenosine phosphosulfate reductase (CysH), and ferredoxin-sulfite reductase (SirA), which are capable of reducing sulfate to organic sulfur compounds. Further, the genomes of strains HSL1-7^T^ and HSL3-1^T^ both encoded polysulfide reductase (Psr) (Figure 2), which could reduce elemental sulfur to sulfide.
3.5.4. Nitrogen Metabolism
The comparative genome analysis revealed that strains HSL1-7^T^ and HSL3-1^T^ contain the complete nitrogen fixation pathway catalyzed by the nitrogenase encoded NifH (nitrogenase iron protein), NifD (nitrogenase Mo-iron alpha chain), and NifK (nitrogenase Mo-iron beta chain) (Figure 2). While in genus Sulfurimonas, only strains S. marisnigri SoZ1, S. lithotrophica GYSZ_1, S. baltica GD2, and S. marina B2 possessed the nitrogen fixation pathways [56]. Mangrove sediments are established as nitrogen-limited ecosystems due to the unbalance of carbon and nitrogen caused by the burying of plant litter or animal corpses. Nitrogen-fixing bacteria are considered to play a key role in nitrogen amendment [3]. Thus, the strains that contained nitrogenase genes in this study may have a competitive advantage in mangrove sediments. Moreover, strains HSL1-7^T^ and HSL3-1^T^ lack the genes encoding all the components required for the complete reduction of nitrate to nitrogen gas. Specifically, the genes for nitrate reductases (NapABGH), nitrite reductases (NirKS), nitric oxide reductases (NorBC), and nitrous oxide reductases (NosZ) were not detected in these strains (Figure 2), suggesting that they are unable to utilize nitrate as a terminal electron acceptor. In addition, strains HSL1-7^T^ and HSL3-1^T^ possessed the complete assimilatory nitrate reduction pathway, including the key enzyme encoding ferredoxin-nitrate reductase (NarAB) and ferredoxin-nitrite reductase (NirA).
3.6. Nitrogen Fixation Activity
The ^15^N_2_ isotope labelling experiments demonstrated that both strains, HSL1-7^T^ and HSL3-1^T^, possessed the capability to fix ^15^N_2_ in the absence of an available nitrogen source in the medium, and that they exhibited similar nitrogen-fixing rates (Figure 3A). Whereas the reference strain Sulfurimonas xiamenensis 1-1N^T^ from the coastal sediments, which lacks nitrogen fixation gene clusters, could not perform nitrogen fixation. Moreover, ^15^N_2_ fixation in both strains were completely inhibited by the addition of ammonia, regardless of whether the concentration was low or high (1 mM and 40 mM of NH_4_Cl) (Figure 3A). This observation aligns with the fact that nitrogen fixation is often repressed when exogenous nitrogen sources are readily available in their environment [57]. While in Sulfurimonas, only strains S. marina B2^T^, S. lithotrophica GYSZ_1^T^, and S. baltica GD2 harbored the complete nitrogenase gene clusters, suggesting that they have the potential for nitrogen fixation (Figure 3B), but this has not been verified so far. These results suggest an important role for nitrogen fixation of these isolates in the nitrogen-limited mangrove ecosystems in which they may be present.
3.7. Taxonomic Conclusion
Based on these results, strains HSL1-7^T^ and HSL3-1^T^ represent two novel species of the genus Sulfurimonas, for which the names Sulfurimonas microaerophilic sp. nov. and Sulfurimonas diazotrophicus sp. nov. are proposed.
3.7.1. Description of Sulfurimonas microaerophilic sp. nov.
Sulfurimonas microaerophilic (mi.cro.ae.ro’phi.lus. Gr. adj. mikros small, little; Gr. n. aer-aeros air; N.L. masc. adj. philus (from Gr. masc. adj. philos) loving; N.L. masc. adj. microaerophilus low-air loving, where the type strain was isolated).
The bacterial cells are Gram-stain-negative, rod-shaped to slightly curved (about 1.8–2.3 µm length × 0.4–0.6 µm in width), exhibiting motility via a single polar flagellum. The isolate was an obligate chemolithoautotroph capable of growing with molecular hydrogen and thiosulfate as an energy source, and molecular oxygen and elemental sulfur as the electron acceptors. Sodium bicarbonate and carbon dioxide can be used as inorganic carbon sources, whereas the growth of the strain was not observed under organic carbon source conditions such as peptone, yeast paste, and starch. The strain exhibited facultative microaerophilic physiology (growth up to 0–10% O_2_ in the gas phase; optimum growth in the presence of 4% O_2_). Growth parameters were established with temperature optima at 32 °C (range 4–40 °C), a salinity preference at 3.0% NaCl (w/v) (range 1.0–4.0%), and a pH optima at 7.0 (range 5.0–8.5). The major cellular fatty acids are C_16:0_ (31.6%), C_16:1_ω7c (28.7%), C_18:1_ω7c (19.5%), and C_14:0_ (10.5%).
The type strain, HSL1-7^T^ (=MCCC 1A18899^T^ = KCTC 25640^T^), was isolated from mangrove sediments of the Jiulong River tributaries in Zhangzhou, Fujian province, China. The genomic DNA G+C content of the type strain is 36.1%. The GenBank accession number for the 16S rRNA gene sequence is OQ102629, and the genome accession number is PRJNA989520, respectively.
3.7.2. Description of Sulfurimonas diazotrophicus sp. nov.
Sulfurimonas diazotrophicus (di.a.zo.trophi.cus. Gr. pref. di two, double; N. L. n. azotum nitrogen; Gr. adj. trophikos tending or feeding; N. L. masc. adj. diazotrophicus, one that feeds on dinitrogen).
The bacterial cells are Gram-stain-negative, displaying rod-shaped morphology (approximately 0.7–2.5 µm length × 0.2–0.5 µm in width) with non-motile characteristics. As a strict chemolithoautotroph, the isolate utilizes molecular hydrogen and thiosulfate as an energy source, while employing molecular oxygen and elemental sulfur as terminal electron acceptors. Nitrogen assimilation occurs through ammonium ions, sodium nitrate, and dinitrogen fixation, whereas growth showed no requirement for selenium, tungsten, or vitamin supplementation. Nitrogen fixation is completely inhibited by NH_4_Cl. Growth occurs at 4–45 °C with optimum growth at 37 °C, and at pH 5.0–8.5 with an optimum at pH 7.0. Growth is observed in the presence of 2.0–4.0% (w/v) NaCl with an optimum at 3.0% NaCl (w/v). The major cellular fatty acids are C_16:1_ω7c (35.0%), C_16:0_ (29.6%), C_18:1_ω7c (16.7%), and C_14:0_ (11.3%).
The type strain, HSL3-1^T^ (=MCCC 1A18844^T^), was isolated from mangrove sediments of the Jiulong River tributaries in Zhangzhou, Fujian province, China. The G+C content of DNA was 57.3%. The GenBank accession number for the 16S rRNA gene sequence is OQ102631 and the genome accession number is PRJNA1085846, respectively.
4. Conclusions
The phylogenetic analysis based on the 16S rRNA gene sequences and whole genome, revealed that strains HSL1-7^T^ and HSL3-1^T^ represent two novel members of the genus Sulfurimonas. The 16S rRNA gene sequence similarity, and the ANI and dDDH values of strains HSL1-7^T^ and HSL3-1^T^ with closely related species, were lower than the thresholds for the delineation of a novel species. These results indicate that strains HSL1-7^T^ and HSL3-1^T^ do not represent any previously described species and should be considered to represent two novel species.
In addition, the physiological and biochemical characteristics of strains HSL1-7^T^ and HSL3-1^T^ differed from those of other species of the genus Sulfurimonas in terms of morphology, major fatty acid fractions, and patterns of electron donor-acceptor utilization. The comparative genomic analysis revealed that strains HSL1-7^T^ and HSL3-1^T^ possessed diverse metabolic profiles, including carbon fixation, hydrogen oxidation, sulfur oxidation, and nitrogen fixation. These unique metabolic capabilities suggests their potential role in driving the biogeochemical cycling of carbon, nitrogen, and sulfur in mangrove ecosystems. Notably, unlike most members of the genus Sulfurimonas, neither strain HSL1-7^T^ or HSL3-1^T^ genomes contained a complete denitrification pathway, but both harbored key genes for nitrogen fixation (nifHDK). Further, the ^15^N_2_ isotope labeling experiments demonstrated that they both exhibited strong nitrogen-fixing abilities. This characteristic implies that they may play an important role in the dark nitrogen fixation process in the mangrove ecosystem. The combined phenotypic, chemotaxonomic and phylogenetic evidence indicates that strains HSL1-7^T^ and HSL3-1^T^ represents two novel species in the genus Sulfurimonas. The findings of our study expand the understanding of Sulfurimonas species and their adaptation to mangrove environments.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Duke N.C. Meynecke J.O. Dittmann S. Ellison A.M. Anger K. Berger U. Cannicci S. Diele K. Ewel K.C. Field C.D. A world without mangroves?Science 2007317414210.1126/science.317.5834.41b 17615322 · doi ↗ · pubmed ↗
- 2Holguin G. Vazquez P. Bashan Y. The role of sediment microorganisms in the productivity, conservation, and rehabilitation of mangrove ecosystems: An overview Biol. Fertil. Soils 20013326527810.1007/s 003740000319 · doi ↗
- 3Wang S. Jiang L. Zhao Z. Chen Z. Wang J. Alain K. Cui L. Zhong Y. Peng Y. Lai Q. Chemolithoautotrophic diazotrophs dominate dark nitrogen fixation in mangrove sediments ISME J.202418 wrae 11910.1093/ismejo/wrae 11938916247 PMC 11474244 · doi ↗ · pubmed ↗
- 4Sengupta A. Chaudhuri S. Ecology of heterotrophic dinitrogen fixation in the rhizosphere of mangrove plant community at the Ganges river estuary in India Oecologia 19918756056410.1007/BF 0032042028313699 · doi ↗ · pubmed ↗
- 5Biderre-Petit C. Courtine D. Hennequin C. Galand P.E. Bertilsson S. Debroas D. Monjot A. Lepère C. Divne A.-M. Hochart C. A pan-genomic approach reveals novel Sulfurimonas clade in the ferruginous meromictic Lake Pavin Mol. Ecol. Resour.202424 e 1392310.1111/1755-0998.1392338189173 · doi ↗ · pubmed ↗
- 6Inagaki F. Takai K. Kobayashi H. Nealson K.H. Horikoshi K. Sulfurimonas autotrophica gen. nov., sp. nov., a novel sulfur-oxidizing epsilon-proteobacterium isolated from hydrothermal sediments in the Mid-Okinawa Trough Int. J. Syst. Evol. Microbiol.2003531801180510.1099/ijs.0.02682-014657107 · doi ↗ · pubmed ↗
- 7Waite D.W. Vanwonterghem I. Rinke C. Parks D.H. Zhang Y. Takai K. Sievert S.M. Simon J. Campbell B.J. Hanson T.E. Comparative Genomic Analysis of the Class Epsilonproteobacteria and Proposed Reclassification to Epsilonbacteraeota (phyl. nov.)Front. Microbiol.20178682 Erratum in Front. Microbiol. 2018, 9, 77210.3389/fmicb.2017.0068228484436 PMC 5401914 · doi ↗ · pubmed ↗
- 8Campbell B.J. Engel A.S. Porter M.L. Takai K. The versatile epsilon-proteobacteria: Key players in sulphidic habitats Nat. Rev. Microbiol.2006445846810.1038/nrmicro 141416652138 · doi ↗ · pubmed ↗