Toll-like Receptor Gene Polymorphisms as Predictive Biomarkers for Response to Infliximab in Japanese Patients with Crohn’s Disease
Jingjing Wei, Hiroki Kurumi, Hajime Isomoto, Ryohei Ogihara, Kayoko Matsushima, Haruhisa Machida, Tetsuya Ishida, Tatsuro Hirayama, Naoyuki Yamaguchi, Yukina Yoshida, Kazuhiro Tsukamoto

TL;DR
This study finds that genetic variations in TLR1 and TLR2 genes can predict how Japanese Crohn’s disease patients respond to infliximab treatment.
Contribution
The study identifies specific TLR gene polymorphisms as novel predictive biomarkers for infliximab response in Japanese Crohn’s disease patients.
Findings
The G/G genotype of rs5743565 in TLR1 is linked to lower response rates at 10 weeks.
C/T or T/T genotypes of rs5743604 in TLR1 and G/A or A/A genotypes of rs13105517 in TLR2 are associated with better early response.
The A/A genotype of rs13105517 in TLR2 is linked to lower response rates after one year of treatment.
Abstract
Objectives: To explore the possible relationship between Toll-like receptor (TLR) gene encoding and a predictive outcome for the loss of response (LOR) to IFX treatment among Japanese patients with Crohn’s disease (CD). Methods: An association analysis that involved 25 single-nucleotide polymorphisms (SNPs) across the TLR1, TLR2, TLR4, TLR6, TLR9, and TLR10 genes was performed on a cohort of 127 Japanese patients with CD. The therapeutic responses were evaluated at 10 weeks, 1 year, and 2 years using three different inheritance models. Results: The CD patients with a G/G genotype of rs5743565 in TLR1 were significantly less likely in the responders at 10 weeks compared with the non-responders (p = 0.023, OR = 0.206). The frequencies of the C/T or T/T genotypes of rs5743604 in the TLR1, G/A, or A/A genotypes of rs13105517 in TLR2, both in the minor allele dominant model, were…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
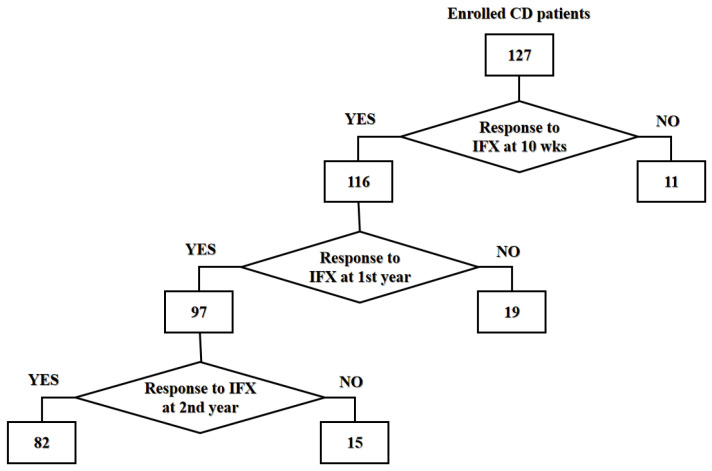 Figure 1
Figure 1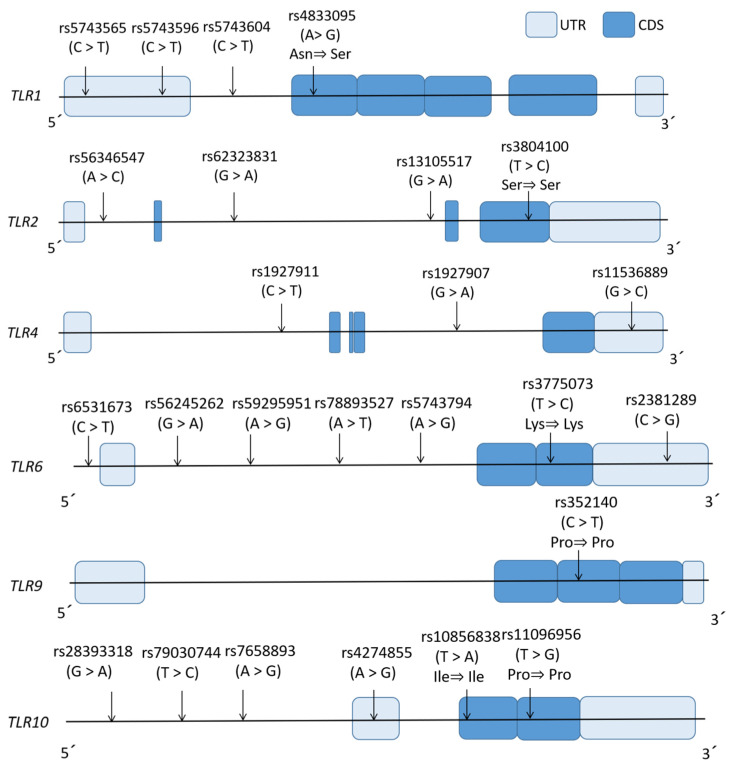 Figure 2
Figure 2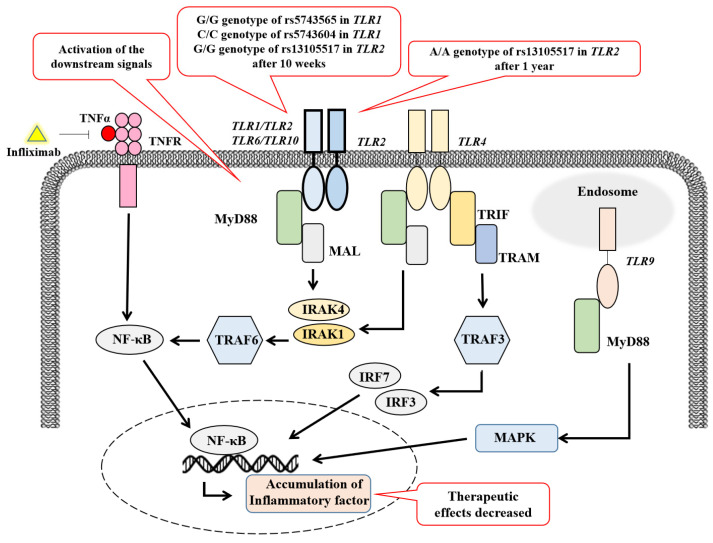 Figure 3
Figure 3Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInflammatory Bowel Disease · Immunodeficiency and Autoimmune Disorders · Microscopic Colitis
1. Introduction
Infliximab (IFX) was the first monoclonal antibody approved for clinical use in treating moderate-to-severe Crohn’s disease (CD) patients who showed an inadequate response to conventional therapy by the FDA in 1998 and was approved in Japan in 2001 [1]. It represented a biopharmaceutical agent characterized as a chimeric immunoglobulin G1 (IgG1) monoclonal antibody that targets tumor necrosis factor-alpha (TNF-α) by binding to it and blocking its interaction with cell receptors [2]. In CD patients, approximately 1030% exhibit primary non-response (PNR) to IFX therapy, while 2346% develop a secondary loss of response (LOR) with continued therapy [3]. However, the mechanisms of IFX non-response or secondary LOR remains unclear. Some multifactorial causes are supported by published data, including complex inflammatory signaling, antibodies produced against IFX [4], and abnormal IFX pharmacokinetics [5]. Further investigation is critically needed to uncover the mechanisms of IFX non-response and develop targeted interventions to improve the clinical outcomes for more CD patients.
Th17/IL-17-signaling-mediated immune response in the pathogenesis of CD
The pathogenesis of inflammatory bowel disease (IBD) involves a dysregulated immune response to alterations in intestinal commensal homeostasis, involving interactions between the gut bacteria [6], immune response [7], genetic susceptibility [8], and environmental triggers [9]. During the past 30 years, immune pathways mediated by Th1 and Th2 cells were considered as key roles in CD and UC, respectively [10]. With the exploration of more and more Th cell subtypes, especially Th17, Th9, and Treg cells [11], the theoretical system has been more extensively elucidated in recent years. Physiologic mucosal Th17 cells contribute to the epithelial barrier function via neutrophil/macrophage recruitment and antimicrobial peptide induction, maintaining an intestinal mucosal defense [12]. Upon the onset of inflammation, dendritic cells can be stimulated by microbial antigens to secrete pro-inflammatory mediators, such as IL-6 and IL-1β, which subsequently induce the pathogenic differentiation of Th17 with the increasing levels of Th17-related cytokines (IL-17, IL-21, and IL-22) [13]. The accumulation of Th17 cells and their associated cytokines (IL-17, IL-21, and IL-22) was observed in the inflammatory lesions of active IBD patients, higher levels of IL-17A and IL-17A mRNA were found in the serum and intestinal tissue of IBD patients when compared with healthy controls [14], and IL-17A could recruit a variety of immune cells by activating the NF-κB and MAPK pathways [15]. These findings imply that the IL-23/IL-17 axis plays key roles in active disease pathogenesis and mucosal injury. Besides inflammatory injury, another study focused on the effect of the Th17/IL-17 axis on intestinal fibrosis, demonstrating the fibrotic progression by activating myofibroblasts and enhancing collagen production via TGF-β, growth factors and novel mediators (TL1A/DR3, Ang-II) [16]. Our previous data also identified the IFX-treatment-response-associated SNPs of IL-17F as promising biomarkers for predicting the therapeutic response to IFX [17].
2.Toll-like receptor (TLR) signaling shapes the Th17/IL-17 axis responses in CD
Other important drivers of Th17 cell proliferation are TLRs signaling pathways, which can promote naive T-cell differentiation into Th17 cells and IL-17 secretion. TLRs are innate immune sensors that recognize pathogen-associated (PAMPs) and damage-associated (DAMPs) molecular patterns, triggering responses to both infectious and non-infectious stimuli [18,19]. In humans, TLR1-10 is expressed in intestinal epithelial cells and leukocytes and can be activated by commensal microbes [20,21,22]. For instance, TLR4 specifically binds lipopolysaccharides [23], while TLR2 recognizes Gram-positive bacterial components (e.g., lipoteichoic acid) and diverse glycolipids/lipoproteins [24]. TLR9 recognizes unmethylated CpG DNA from microbes within endosomes [25], unlike membrane-bound TLRs (TLR1/2/4/6/10) [26]. TLR4 acts as a homodimer, while TLR2 forms heterodimers with TLR1/6/10 [27,28,29,30]. While normally detecting microbial PAMPs, TLRs initiate NF-κB-mediated cytokine production to combat pathogens [31]. TLR gene polymorphisms are related with an elevated susceptibility to CD. TLR1/2/6/9 polymorphisms may be involved in disease progression in IBD patients [32,33]. However, further genetic evidence shows that the Saudi population with three major CARD15/NOD2 variant alleles and the CD14-159C/T polymorphism but not TLR4 (Thr399Il) are more susceptible to CD [34].
TLRs play a crucial role in infection defense, but overactivation may lead to inflammatory conditions and autoimmune disorders across acute and chronic spectrums [35]. Inappropriate TLR activation could lead to an imbalance in intestinal homeostasis, thereby resulting in a primary non-response and subsequent LOR to IFX therapy in CD patients. Genetic evidence suggests that TLRs are strongly associated with both a CD pathology and LOR to IFX treatment. In a Danish patient study, polymorphisms in TLR1/2/4/6/9/10 were associated with an anti-TNF treatment response [36]. Given the role of TLRs in inflammation, TLRs inhibitors are being developed as potential therapeutic drugs for inflammatory and autoimmune disorders [37,38].
This study aimed to explore TLR gene polymorphisms as predictive indicators of IFX treatment in Japanese CD patients. Based on the evidence, the IFX-treatment-related financial burden and side effects, such as increased susceptibility to infections, autoimmune disorders, and malignancies, could be prevented in non-responders to IFX. This will be meaningful in clinical decision-making by predicting IFX responsiveness in CD patients before therapy.
2. Materials and Methods
A multicenter cohort of 127 Japanese CD patients that initiated IFX therapy between 2004 and 2012 was prospectively evaluated across three tertiary care institutions: Oita Red Cross Hospital, Nagasaki Harbor Medical Center City Hospital, and Nagasaki University Hospital. This study was approved by institutional ethics committees (approval ID: 110926210) in compliance with the Declaration of Helsinki, which required written informed consent from all participants prior to enrollment.
The treatment efficacy was stratified using a composite clinical endpoint: the responders demonstrated sustained Crohn’s Disease Activity Index (CDAI) scores <150 alongside concurrent improvements in symptomatology, biochemical markers, or endoscopic findings across the treatment intervals, whereas the non-responders exhibited static/elevated CDAI values (>150, indicative of active disease) or clinical deterioration. Longitudinal monitoring comprised sequential evaluations at 10 weeks post-induction (n = 127), followed by annual assessments of the initial responders (n = 116, 91.3%) over a two-year period. Secondary LOR occurred in 19/116 (16.3%) and 15/97 (15.5%) patients at years 1 and 2, respectively, with demographic and baseline clinical parameters detailed in Table 1 (study schema: Figure 1).
The genetic analysis focused on six TLR loci (TLR1, TLR2, TLR4, TLR6, TLR9, TLR10), with tag SNPs selected from the 1000 Genomes Project Phase 3 Tokyo Japanese cohort (hg19/GRCh37) using Haploview v4.2 (MAF ≥ 0.2, LD r^2^ ≥ 0.8) [39,40]. The gene architectures illustrating untranslated regions, coding sequences, and SNP positions (including exonic variants with/without amino acid substitutions) are depicted in Figure 2. Peripheral blood-derived genomic DNA isolated via the WB-Rapid Kit (Fujifilm Wako Chemicals. Co., Osaka, Japan) served as a template for the tripartite genotyping of 25 SNPs.
The PCR-RFLP analyses employed GeneAmp 9700/T100 thermal cyclers (Thermo Fisher Scientific. Co., Waltham, MA, USA) with a GoTaq Green Master Mix (Promega. Co., Tokyo, Japan), followed by restriction enzyme digestion (Table 2) and electrophoretic separation on polyacrylamide/agarose gels visualized via ethidium bromide staining. Sanger sequencing utilized ExoSAP-IT-purified amplicons sequenced with BigDye Terminator v3.1 on ABI 3100/3130xl platforms (Thermo Fisher Scientific. Co., Waltham, MA, USA), while high-resolution melting (HRM) profiling was conducted on a LightCycler 480 system using SYTO9/DMSO-enhanced reactions (Invitrogen Life Technologies, Carlsbad, CA, USA) analyzed through derivative curve modeling (Gene-Scanning v1.3) (Roche Diagnostic. Co., Tokyo, Japan).
The statistical workflows integrated dual-platform validation (SPSS v20/Prism 6), beginning with baseline comparisons via Mann–Whitney U and Fisher’s exact tests. Genetic associations were evaluated under allelic/dominant/recessive models using SNPAlyze v7.0 for Hardy–Weinberg equilibrium and linkage disequilibrium analyses, with multivariate logistic regression modeling of the gene–treatment interactions. The significance thresholds were set at p < 0.05 or 95% confidence intervals, excluding unity.
3. Results
3.1. Clinical Characteristics of Study Population
No significant differences were observed between the responders and non-responders groups (Table 1). Finally, a total number of 28 CD patients (25/127, 19.7%) underwent surgeries during the therapy. The ratio of surgeries in the non-responders of IFX-treated CD patients was 54.5% (6/11) at 10 wks, 47.4% (9/19) at year 1, and 66.7% (10/15) at year 2.
3.2. Polymorphisms Associated with the Response to IFX at the 10-Week Treatment
Statistical analyses showed that the patients with a G/G genotype of rs5743565 in TLR1 in the responders in the minor allele recessive model were significantly less likely to respond to IFX at the 10-week treatment as compared with the non-responders (p = 0.023, OR = 0.206; Table 3), suggesting a 4.9-fold higher chance of non-response to IFX. Conversely, the patients with an A/A or A/G genotype of rs5743565 had a 4.9-fold higher likelihood of responding to IFX.
Meanwhile, the frequency of the C/T or T/T genotype of rs5743604 in TLR1 in the minor allele dominant model was significantly higher in the responders at the 10-week treatment as compared with that in the non-responders (p = 0.035, OR = 4.401; Table 3), indicating a 4.4-fold increase in the response to IFX. In contrast, a C/C genotype of rs5743604 was associated with a 4.4-fold decrease in the response to IFX.
In addition, the patients with a G/A or A/A genotype of rs13105517 in TLR2 in the minor allele dominant model were significantly increased in the responders at the 10-week treatment in comparison with those in the non-responders (p = 0.017, OR = 5.473; Table 3). This indicated a 5.5-fold higher likelihood of responding to IFX. Conversely, the patients with a G/G genotype of rs13105517 were 5.5 times more likely to be non-responders to IFX.
3.3. Interaction of the Genetic Factors in Response to IFX at the 10-Week Treatment
The differences in the genotype frequencies between the responders and non-responders revealed several genetic factors associated with the IFX response at the 10-week treatment: the A/A or A/G genotype of rs5743565 in TLR1, the C/T or T/T genotype of rs5743604 in TLR1, and the G/A or A/A genotype of rs13105517 in TLR2. Further multivariate logistic regression analysis revealed that the A/A or A/G genotype of rs5743565 in TLR1 and the G/A or A/A genotype of rs13105517 in TLR2 independently contributed to the IFX response (p = 0.015 and 0.014, respectively; Table 4).
3.4. Verification of Genetic Test to Predict the Response to IFX at the 10-Week Treatment
To better predict the IFX response in the CD patients at the 10-week treatment, a genetic test was conducted using either a single genetic factor (TLR1 or TLR2) or a combination of these independent genetic factors as biomarkers. The combination of the A/A or A/G genotype of rs5743565 in TLR1 and the G/A or A/A genotype of rs13105517 in TLR2 showed a significant response to IFX (p = 0.024, OR = 5.735; Table 5). For this genetic test, the sensitivity, specificity, positive predictive value (PPV), and negative predictive value (NPV) were estimated at 56.0%, 81.8%, 97.0%, and 15.0%, respectively. This combination of biomarkers showed a better specificity and PPV.
3.5. Polymorphisms Associated with the IFX Response at the 1-Year Treatment
The statistical analyses showed that the patients with an A/A genotype of rs13105517 in TLR2 in the responders in the minor allele recessive model were significantly less likely to respond to the IFX at the 1-year treatment than those in the non-responders (p = 0.004, OR = 0.195; Table 6), indicating a 5.1-fold decrease in the IFX response. In contrast, the patients with a G/G or G/A genotype of rs13105517 showed a 5.1-fold higher response to IFX.
3.6. Verification of Genetic Testing for Predicting the IFX Response at the 1-Year Treatment
To better predict the IFX response in CD patients at the 1-year treatment, a genetic test with the G/G or G/A genotype of rs13105517 was conducted. The sensitivity, specificity, PPV, and NPV of this test were 91.8%, 31.6%, 87.3%, and 42.9%, respectively (Table 7). This biomarker demonstrated a superior sensitivity and PPV.
3.7. Polymorphisms Associated with the IFX Response at the 2-Year Treatment
There were no significant differences in the allele and genotype frequencies of tag SNPs between the responders and non-responders at the 2-year treatment (Table 8).
4. Discussion
Our results suggest that TLR1 and TLR2 may be involved in both the initial non-response and SLOR to IFX in Japanese CD patients. However, the functional roles of rs5743565 and rs5743604 in TLR1 and rs13105517 in TLR2 remain unclear. The HaploReg analysis indicated that rs5743565 and its highly related variants with high LD (r^2^ ≥ 0.8) could alter the transcription or chromatin states. This in silico analysis showed different affinities to more than 20 transcription factors for the reference and alternative alleles of rs5743565 and several variants with a high LD. In contrast, since rs5743604 does not have a high LD with other common variants, it demonstrated different affinities for the four transcription factors and distinct expression quantitative trait loci between its reference and alternative alleles. rs13105517 and variants with high LDs can also predict different affinities for several transcription factors [41]. Based on these results, the TLR1 and TLR2 polymorphisms could induce the gain of function in CD patients with specific genotypes: the G/G genotype of rs5743565 in TLR1, the C/C genotype of rs5743604 in TLR1 at the 10-week treatment, and the A/A genotype of rs13105517 in TLR2 at both the 10-week and 1-year treatments. These changes could contribute to the activation of intracellular downstream signals in the TLR signaling pathway, which eventually resulted in the acceleration of chronic intestinal inflammatory processes. As shown in Figure 3, persistent intestinal inflammation could diminish the IFX efficacy, which led to both an initial non-response to IFX at the 10-week treatment and a secondary LOR to IFX at the 1-year treatment. Some evidence shows that a transcriptional dysregulation of circulating monocytes would induce hyperactivation of the pro-inflammatory pathways, contributing to the resistance to IFX treatment [42]. Another activated pro-inflammatory pathway mediated by IL-1β, IL-6, and IFN-γ in CD patients, different from TNF-α signaling, might be resistant to anti-TNF treatment [36], suggesting that treatment strategies should be broadened for CD patients.
Conversely, the CD patients with specific genotypes could experience the loss of function of TLR1 and TLR2. This included patients with the A/A or G/A genotype of rs5743565 in TLR1, C/T or T/T genotype of rs5743604 in TLR1 at the 10-week treatment, and G/A or G/G genotype of rs13105517 in TLR2 at both the 10-week and 1-year treatments. This could inhibit the intracellular downstream signals, which eventually resulted in the continued suppression of signals in the TNFR signaling pathway. Therefore, these patients could benefit from IFX at the 10-week and 1-year treatments in this study.
Gut homeostasis involved in the mechanism of IFX therapy is under investigation. A functional I602S SNP in TLR1 can regulate the innate immune response to lipopeptides caused by a pathogen infection, which could potentially affect inflammatory pathways [43], although there is no study about this allele in the Japanese population [44]. An in silico analysis of microarray experiment GSE16879 in the GEO database revealed higher expressions of TLR1 and TLR2 in CD patients resistant to IFX treatment. After an IFX treatment, the downregulation of TLR2 was found in patients who responded to IFX, but remained unchanged in non-responders [45] (data accessible at NCBI GEO database [46], accession GSE16879). Evidence suggests that the polymorphisms of TLR1 and TLR2 might be strongly related to the non-response and LOR to IFX in CD patients.
In our study, the combination of the G/A or A/A genotype of rs5743565 in TLR1 and the G/A or A/A genotype of rs13105517 in TLR2 proved to be a useful biomarker for predicting the IFX response, showing a high OR of 5.735, specificity of 81.8%, and PPV of 97.0% in genetic testing at the 10-week treatment. A higher PPV indicates that the CD patients with this combination polymorphism were more likely to respond successfully to the IFX treatment at the 10-week mark. Likewise, the G/G or G/A genotype of rs13105517 in TLR2 could be considered a useful biomarker for predicting an IFX response, with a higher OR of 3.736, sensitivity of 91.8%, and PPV of 87.3% in genetic testing at the 1-year mark after treatment, indicating that the CD patients with this polymorphism are more likely to respond successfully to IFX treatment and be continuously effective for at least 1 year.
Taken together, our findings demonstrate significant associations between TLR1/2 genetic variants and a differential response to IFX in CD patients. The results should be interpreted with consideration of its limitations. First, the data were based on a small cohort limited to the Japanese population; future studies should be considered to investigate diverse ethnic groups. The optimal combination of genetic biomarkers for a better prediction of IFX effectiveness should also be discussed in the future. Furthermore, the exact mechanisms are essential for the refinement of therapeutic strategies for IFX non-response in CD patients.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Croft M. Salek-Ardakani S. Ware C.F. Targeting the TNF and TNFR superfamilies in autoimmune disease and cancer Nat. Rev. Drug Discov.20242393996110.1038/s 41573-024-01053-939448880 · doi ↗ · pubmed ↗
- 2Leone G.M. Mangano K. Petralia M.C. Nicoletti F. Fagone P. Past, Present and (Foreseeable) Future of Biological Anti-TNF Alpha Therapy J. Clin. Med.202312163010.3390/jcm 1204163036836166 PMC 9963154 · doi ↗ · pubmed ↗
- 3Ben-Horin S. Chowers Y. Review article: Loss of response to anti-TNF treatments in Crohn’s disease Aliment. Pharmacol. Ther.20113398799510.1111/j.1365-2036.2011.04612.x 21366636 · doi ↗ · pubmed ↗
- 4Navajas Hernández P. Mouhtar El Halabi S. González Parra A.C. Valdés Delgado T. Maldonado Pérez B. Castro Laria L. Charpentier C. Argüelles-Arias F. Carriage of the HLA-DQA 1⋆05 haplotype is associated with a higher risk of infratherapeutic drug concentration and higher immunogenicity in patients undergoing treatment with anti-TNF for inflammatory bowel disease Therap. Adv. Gastroenterol.2024171756284824127814510.1177/17562848241278145 PMC 1148369739421002 · doi ↗ · pubmed ↗
- 5Srinivasan A. De Cruz P. Sam M. Toong C. van Langenberg D.R. Dose intensification strategy influences infliximab pharmacokinetics but not clinical response after the same number of doses J. Gastroenterol. Hepatol.20233872473210.1111/jgh.1613336692034 · doi ↗ · pubmed ↗
- 6Qiu P. Ishimoto T. Fu L. Zhang J. Zhang Z. Liu Y. The Gut Microbiota in Inflammatory Bowel Disease Front. Cell. Infect. Microbiol.20221273399210.3389/fcimb.2022.73399235273921 PMC 8902753 · doi ↗ · pubmed ↗
- 7Saez A. Herrero-Fernandez B. Gomez-Bris R. Sánchez-Martinez H. Gonzalez-Granado J.M. Pathophysiology of Inflammatory Bowel Disease: Innate Immune System Int. J. Mol. Sci.202324152610.3390/ijms 2402152636675038 PMC 9863490 · doi ↗ · pubmed ↗
- 8Jarmakiewicz-Czaja S. Zielińska M. Sokal A. Filip R. Genetic and Epigenetic Etiology of Inflammatory Bowel Disease: An Update Genes 202213238810.3390/genes 1312238836553655 PMC 9778199 · doi ↗ · pubmed ↗