Molecular Characterization and Nutritional Regulation of Two Fatty Acid Elongase (elovl8) Genes in Chinese Perch (Siniperca chuatsi)
Yu He, Zhengyong Wen, Luo Zhou, Wanhong Zeng, Panita Prathomya, Tilin Yi, Qiong Shi

TL;DR
This study identifies and characterizes two elovl8 genes in Chinese perch, showing their roles in long-chain fatty acid synthesis and response to diet.
Contribution
The study reports the molecular and evolutionary features of two elovl8 paralogues in Chinese perch for the first time.
Findings
Scelovl8a and Scelovl8b are highly conserved in teleosts and clustered into distinct subclades.
Scelovl8a is highly expressed in the liver, while Scelovl8b is most abundant in the gills.
Dietary interventions like food deprivation and oil injections regulate the expression of both elovl8 genes.
Abstract
Proteins for elongation of very long-chain fatty acids (ELOVLs) are critical for the synthesis of long-chain polyunsaturated fatty acids (LC-PUFAs), and they are one group of the rate-limiting enzymes responsible for the initial condensation reaction within the fatty acid elongation. Elovl8 is a newly identified member of the ELOVL protein family, and its evolutionary and functional characterizations are still rarely reported. Here, we identified two elovl8 paralogues (named Scelovl8 and Scelovl8b) from Chinese perch (Siniperca chuatsi), and then their molecular and evolutionary characteristics, as well as potential roles involved in LC-PUFA biosynthesis, were examined. The ORFs of both Scelovl8a and Scelovl8b genes were 810 bp and 789 bp in length, encoding proteins of 270 and 263 amino acids, respectively. Multiple protein sequence comparisons indicated that elovl8 genes were highly…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
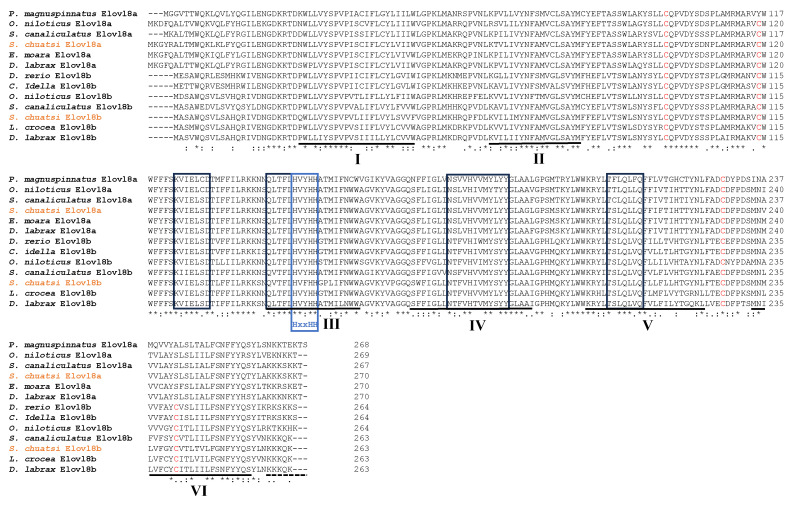 Figure 1
Figure 1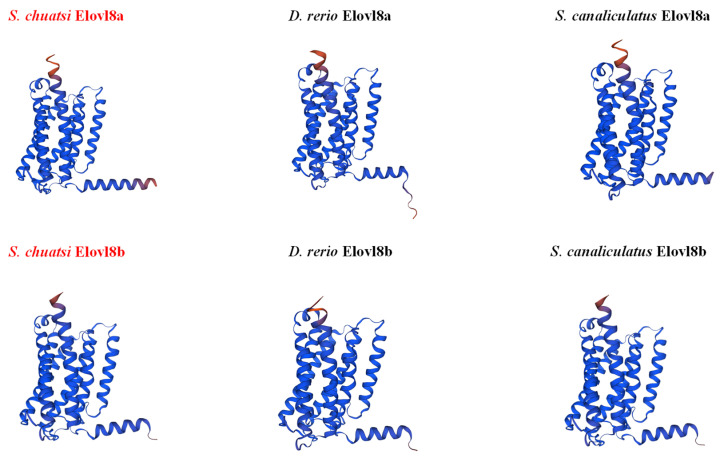 Figure 2
Figure 2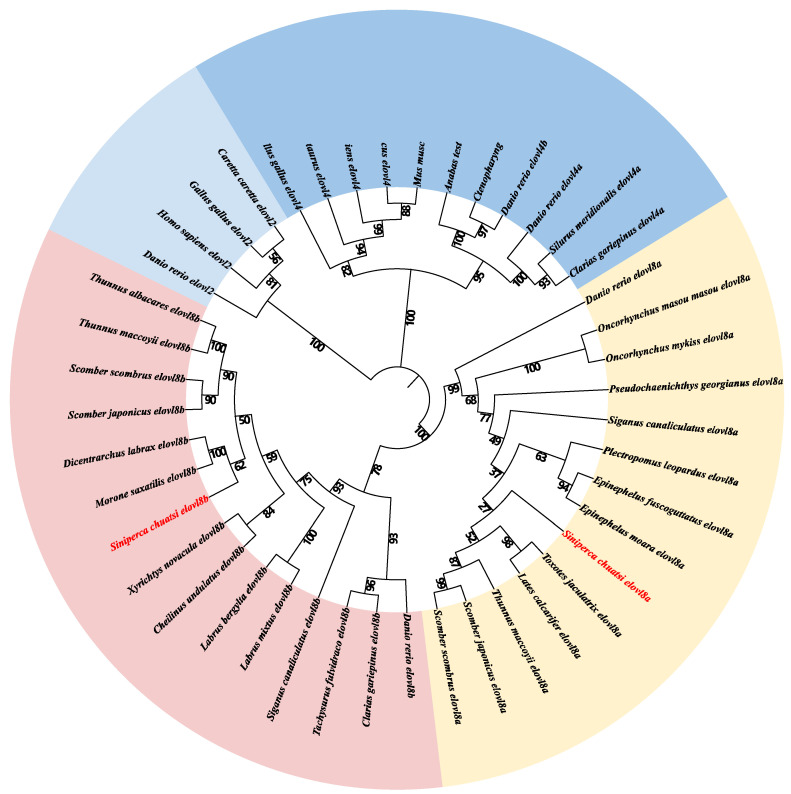 Figure 3
Figure 3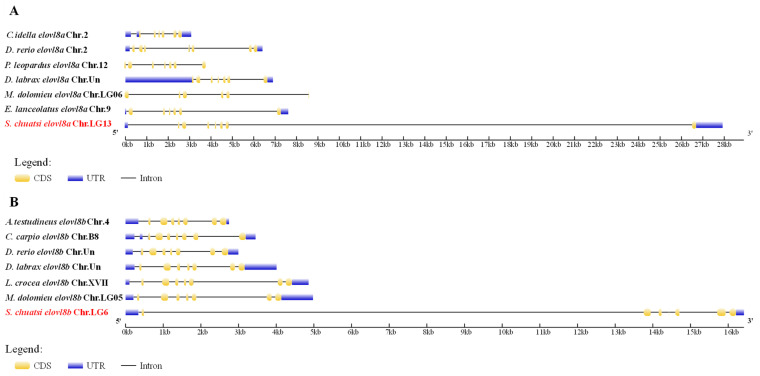 Figure 4
Figure 4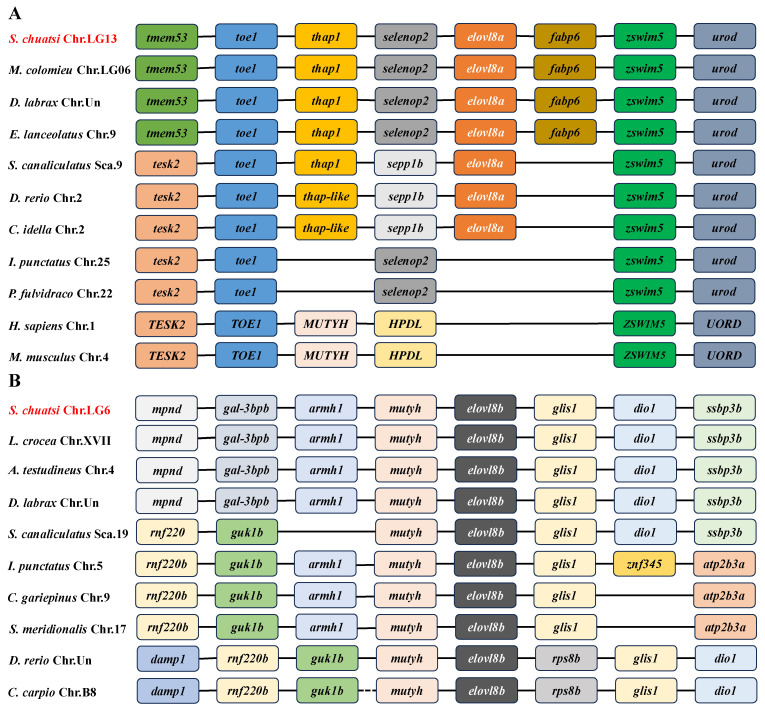 Figure 5
Figure 5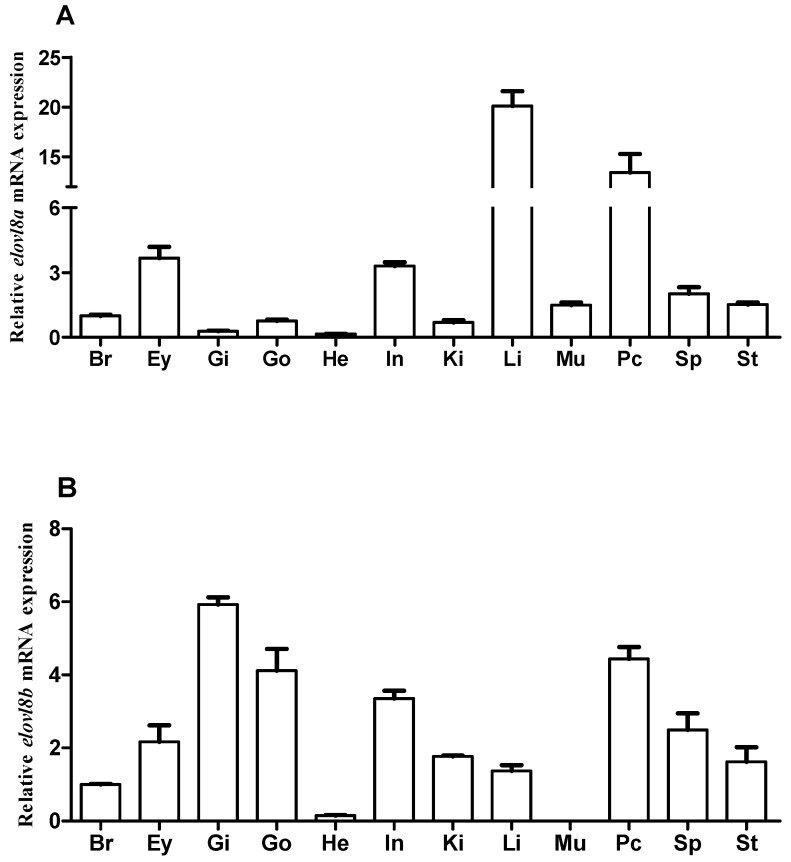 Figure 6
Figure 6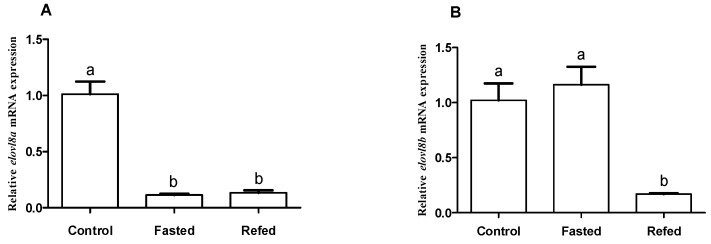 Figure 7
Figure 7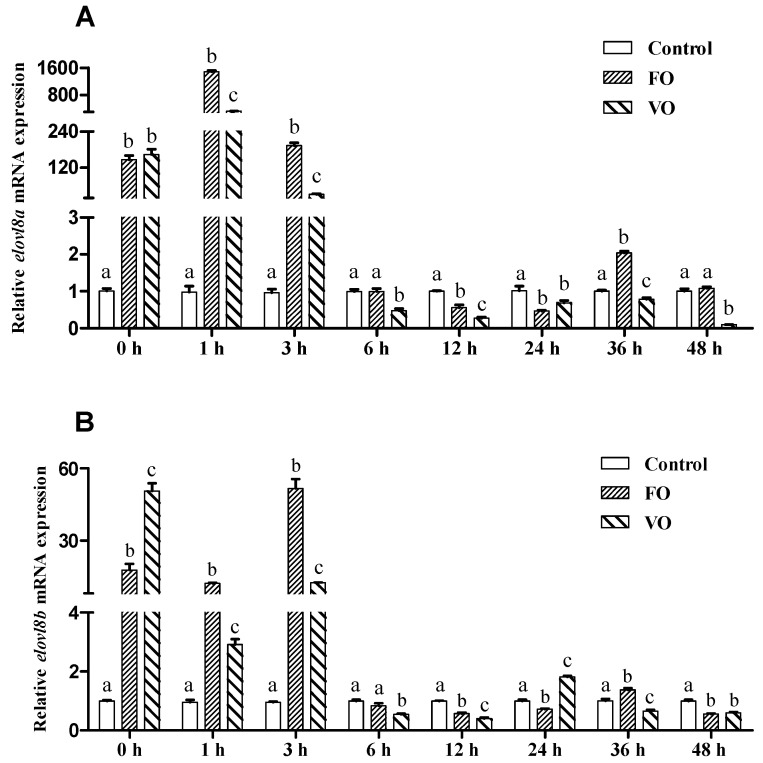 Figure 8
Figure 8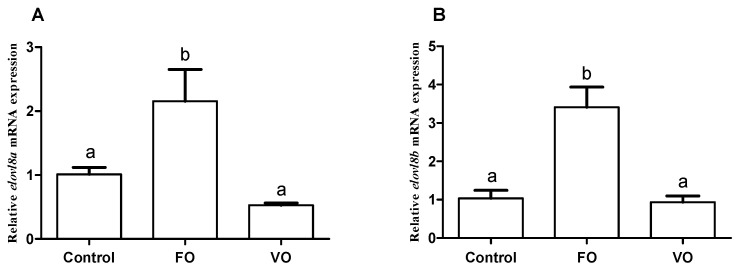 Figure 9
Figure 9- —Natural Science Fund of Sichuan Province of China
- —Project of Sichuan Provincial Department of Science and Technology
- —Key Laboratory of Sichuan Province for Fishes Conservation and Utilization in the Upper Reaches of Yangtze River
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAquaculture Nutrition and Growth · Aquaculture disease management and microbiota · Peroxisome Proliferator-Activated Receptors
1. Introduction
Long-chain polyunsaturated fatty acids (LC-PUFAs) are a series of straight-chain fatty acids with three or more double bonds and containing 20 to 24 carbon atoms, which play important roles in maintaining cell membrane fluidity, regulating fat metabolism, and enhancing immunity [1,2,3,4]. LC-PUFAs, especially arachidonic acid (ARA 20:4n-6), eicosapentaenoic acid (EPA 20:5n-3), and docosahexaenoic acid (DHA 22:6n-3), are physiologically important compounds required for normal growth and development in mammals and play critical roles in reducing inflammation, cardiovascular disease, and certain types of cancers [3,5]. In teleosts, LC-PUFAs have been proved to play important roles in fish development, growth, survival, pigmentation, stress, and disease resistance [6].
LC-PUFAs are essential nutrients that can be obtained directly from exogenous food resources or synthesized from endogenous precursor substances [7]. Endogenous synthesis of LC-PUFAs requires two key rate-limiting enzymes, including fatty acyl desaturases (Fads) and fatty acid elongases (Elovls; for elongation of very long-chain fatty acids protein) [1,7]. Among them, Fads can desaturate polyunsaturated fatty acids (PUFAs) by introducing double bonds at specific positions in the carbon chain, while ELOVLs are responsible for the initial condensation reaction within the fatty acid elongation [8,9,10]. Thus far, seven members of the ELOVL protein family (termed ELOVL1~7) have been identified in various mammals based on their protein characteristics and substrate specificity [10]. Some typical features of ELOVL family members include several transmembrane α-helices (TMHs), an endoplasmic reticulum (ER) retention signal, and four ‘conserved boxes’ (KEDT-, His-, Tyr-, and Gln-boxes) involved in the catalytic site [11,12]. Among them, ELOVL1, 3, 6, and 7 mainly have the ability to prolong saturated and monounsaturated fatty acids (MUFAs), while ELOVL2, 4, and 5 mainly can prolong PUFAs [10,13,14]. Interestingly, a new elovl gene family member (elovl8) was recently discovered, and it contained two paralogs in examined teleosts [11]. Certain studies have shown that the elovl8 gene is more closely related to elovl4, which could explain the previous misleading naming as elovl4-like [11,15].
The elovl8b gene was first identified in the freshwater teleost African catfish, and functional tests showed that it can prolong C18 and C20 PUFAs [16]. Subsequently, both elovl8a and elovl8b genes were identified from the herbivorous marine teleost rabbitfish (Siganus canaliculatus), and they were found to be widely distributed in examined tissues [11]. Interestingly, elovl8a expression was at the highest level in the heart, while elovl8b level was the highest in the brain [11]. Elovl8b was proved to have an ability to convert C18 PUFA (18:2n-6, 18:3n-3, and 18:4n-3) and C20 PUFA (20:4n-6 and 20:5n-3) to LC-PUFAs, but Elovl8a has lost this functionality [11]. Another study reported two elovl8 genes in zebrafish (Danio rerio) but considered elovl8 not unique to teleosts due to a rather unique and ample phylogenetic distribution [17]. Knocking out both elovl8a and elovl8b genes in zebrafish resulted in an accumulation of C20 fatty acids in the liver and a block in the synthesis of C22:0, as well as reduced resistance ability to cold stress [18]. The tissue distribution profile of elovl8 in hybrid grouper (Epinephelus fuscoguttatus ♀× Epinephelus lanceolatus ♂) was similar to the pattern of elovl8b in rabbitfish, with higher levels in the brain and liver [11,19]. Recently, we identified a single elovl8b gene in yellow catfish (Pelteobagrus fulvidraco), while the elovl8a gene has been lost in the yellow catfish genome [20]. Further functional experiments revealed that diets full of LC-PUFAs can inhibit elovl8b transcription in the liver, suggesting the existence of a negative feedback regulation of the LC-PUFA synthetic pathway [20]. In addition, elovl8a and elovl8b genes were recently identified in an amphidromous fish (ayu sweetfish) [21]. However, the evolutionary and functional characteristics of this new elongase member are still largely unclear.
Chinese perch (Siniperca chuatsi), belonging to the order Centrarchiformes and the family Sinipercidae, is widely distributed in China, Russia, North Korea, South Korea, and Japan [22,23]. This fish has gained a lot of popularity due to its advantages of tender meat, tasty flavor, and absence of interstitial spines [22]. As an economically important fish in China, its annual production was approximately 400,000 tons with high commercial value [24,25]. In our present study, we identified two elovl8 genes in Chinese perch, and then their evolutionary and functional characterizations were explored. Our findings will further enrich the understanding about endogenous biosynthetic mechanisms of LC-PUFAs and related regulatory mechanisms in teleosts.
2. Materials and Methods
2.1. Fish Sampling
Individuals of Chinese perch were purchased from a local fish farm in Neijiang City, Sichuan Province, China. They were moved into 1.5 m × 2 m × 2 m ponds at the Key Laboratory of Sichuan Province for Fishes Conservation and Utilization in the Upper Reaches of the Yangtze River, Neijiang Normal University, Neijiang, China. These fish were temporarily reared under natural light conditions (12 L/12D) for two weeks, with the water temperature between 24 and 26 °C. They were fed with larvae fish daily at 8:00 and 18:00, and the water was changed by 1/3 every 2–3 days.
Three healthy Chinese perch (average body weight: 35 ± 6.25 g) were randomly selected and anesthetized with MS-222, and then their brain, pyloric cecum, gill, gonad, heart, intestine, kidney, liver, muscle, spleen, and stomach tissues were collected. Tissue samples were immediately frozen in liquid nitrogen and then stored at −80 °C until use.
Twelve healthy Chinese perch (average body weight: 45 ± 4.38 g) were selected for a long-term starvation experiment. They were randomly divided and cultured in three tanks (n = 4 for each group) with a diameter of 1 m. One group was set as the normal feeding group (control), and the other two groups were arranged as fasting treatment and fasting-refeeding treatment, respectively. The water temperature was kept at 24–26 °C, and the dissolved oxygen was maintained at 6 mg/L or above. The experiment lasted for 18 d, and then the control group and refeeding group were fed at 18:00; at 19:00, three fish from each group were randomly selected and anesthetized with MS-222, and their livers were rapidly collected and stored as described above.
Seventy-two healthy Chinese perch (average body weight: 13 ± 1.13 g) were selected and randomly divided into three experimental groups. The control group was intraperitoneally injected with 10 μL of saline, and two experimental groups were intraperitoneally injected with 10 μL of fish oil (FO) or vegetable oil (VO), respectively. Subsequently, three fish were randomly taken from each group, anesthetized with MS-222, and their livers were rapidly collected at 0 h, 1 h, 3 h, 6 h, 12 h, 24 h, and 48 h after the injection.
In addition, 150 completed domesticated Chinese perch (average body weight: 45.5 ± 5.3 g) were purchased and randomly divided into nine 50 × 50 × 50 cm glass tanks in three treatments with three replicates per treatment. During the experiment period, the fish were fed with three different diets, including a commercial diet, a commercial diet supplemented with 2% fish oil, or a commercial diet supplemented with vegetable oil. Fatty acid composition of the experimental diets and details of the proximate composition were determined by the area normalization method, and detailed information was summarized in Table 1. At the end of the 2-week feeding trial, four fish were randomly selected from each group, and their livers were sampled and stored quickly.
2.2. Identification of the Scelovl8a and Scelovl8b Genes in Chinese Perch
We obtained genomic and mRNA sequences of Scelovl8a and Scelovl8b genes by scanning the whole-genome and transcriptome databases created by our research group previously (CNA0013732) [26], and then these isolated sequences were validated by NCBI-BLASTn with highly conserved sequence identity to corresponding genes in the NCBI database (accession numbers: XM_044218690.1 and XM_044200424.1). Quantitative primers for detecting the mRNA expression levels of both elovl genes in Chinese perch (listed in Table 2) were designed using Primer Premier 5.0 (Premier Biosoft International, Palo Alto, CA, USA). qRT-PCR was performed using related quantitative primers and cDNA from the liver of Chinese perch. Agarose gel electrophoresis was conducted at the end of PCR amplification to determine primer specificity and verify the existence of Scelovl8a and Scelovl8b transcripts in the liver.
2.3. Bioinformatics Analyses of the Scelovl8a and Scelovl8b Genes
Putative protein sequences were predicted using the Primer Premier 5.0 software, and then multiple protein sequence comparisons of Elovl8a and Elovl8b were performed using Clustal X (http://www.clustal.org). Transmembrane domains were identified using TMHMM Server v. 2.0 and validated by comparative analysis according to a previous study [11]. SWISS-MODEL (https://swissmodel.expasy.org) was utilized to predict the three-dimensional structures of Elovl8a and Elovl8b of several representative species. Meanwhile, we performed comparative analyses of genomic synteny and gene structure of both elovl8a and elovl8b genes by using some representative teleost genomic data and/or related results from previous reports to determine their conservation during teleost evolution [11,20].
In addition, a phylogenetic analysis was conducted to explore the evolutionary history of the elovl8 gene family. Protein sequences used for the phylogeny were downloaded from the NCBI or Ensembl database. After calculation, JTT + G was chosen as the best model to construct a phylogenetic tree using the neighbor-joining (NJ) method. Meanwhile, both Bayesian inference and the maximum likelihood methods were performed, sharing a similar topology to validate the phylogeny (see more details in Supplementary Figures S1 and S2). Finally, the accuracy of the tree topology was assessed by a non-parametric bootstrap analysis with 1000 resampling replicates. Resources of the selected ELOVL family proteins are provided in Table 3.
2.4. Total RNA Extraction and cDNA Preparation
Total RNA was extracted from 10 to 20 mg of each sample using the TRIzol agent (Thermo Fisher Scientific, Waltham, MA, USA), and then its concentration was measured by a micro-protein nucleic acid meter (Thermo Fisher Scientific) for maintenance at 100–1000 ng/μL. Subsequently, the quality of extracted RNA was examined by gel electrophoresis [27,28]. Finally, the extracted RNA was stored at −80 °C for further utilization.
In general, 1 μg of total RNA was reverse transcribed into cDNA using a cDNA. First Strand Synthesis kit (Novoprotein, Shanghai, China). The final volume of the reaction was set as 20 μL, and the reaction procedure was designed as follows: 50 °C for 15 min and then 85 °C for 30 s. After that, the quality of transcribed cDNA was measured by PCR and gel electrophoresis, and the β-actin gene was used as the internal reference.
2.5. Quantitative Real-Time PCR
Quantitative real-time PCR (qRT-PCR) was conducted to detect the transcription levels of Scelovl8a and Scelovl8b on a Light Cycler real-time system (Roche Diagnostics, Indianapolis, IN, USA) with a final volume of 20 μL. Meanwhile, the relative expression levels of mRNAs were normalized by β-actin after assessing the stability of five internal reference genes. The primers of β-actin (Table 1) were obtained and selected from a previous study [29]. Finally, relative expression levels were calculated with the Pfaffl method [30]. Specific primer sequences used for the qRT-PCRs are listed in Table 1.
2.6. Statistical Analysis
SPSS 26.0 (IBM, Armonk, NY, USA) and GraphPad Prism 5.0 (GraphPad Prism Software Inc., San Diego, CA, USA) were applied for statistical analysis. All data were presented as mean ± standard error of the mean (SEM). Significant differences were determined using one-way analysis of variance (ANOVA), followed by Tukey’s test, and differences were designed as significant when p < 0.05.
3. Results
3.1. Presence of Both elovl8a and elovl8b Genes in Chinese Perch
In our present study, both elovl8a and elovl8b genes were identified for the first time from Chinese perch. The open reading frame (ORF) of Scelovl8a was 810 bp, encoding a protein of 270 amino acid (aa) residues, whereas the ORF of Scelovl8b was 789 bp, encoding a protein of 263 aa (Figure 1). In comparison with several teleost Elovl8s, the protein sequences of ScElovl8s were found to be highly conserved, containing six conserved transmembrane α-helical structural domains (TMHs), four conserved functional elongase motifs, a highly conserved histidine structural domain, and three highly conserved cysteine residues (see more details in Figure 1). Notably, Elovl8a is commonly 5 aa longer than Elovl8b in teleosts. Additionally, a highly conserved cysteine residue was identified in the sixth transmembrane region of Elovl8b rather than Elovl8a (see Figure 1).
3.2. Predicted 3D Structures of Both Elovl8a and Elovl8b in Chinese Perch
The 3D structures of Elovl8 proteins of several representative fish species were predicted, and our results showed that both ScElovl8a and ScElovl8b were highly similar to those in yellow catfish and rabbitfish (Figure 2). These Elovl8s have eight longer α-helix structures, seven of which are similar in positional space and size, with a relatively less coherent α-helix structure at the N-terminus (see more details in Figure 2). It is evident that the Elovl8b structures of the three species are more conserved in comparison with Elovl8a.
3.3. Phylogenetic Tree of the Examined Elovl Genes in Teleost
For a better understanding of the evolutionary relationship of elovls in teleosts, we constructed a relative phylogenetic tree with MEGA-X using the protein sequence datasets of representative teleost species. The tree is obviously divided into three subfamilies, including elovl2, elovl4, and elovl8; among them, the elovl4 and elovl8 clades are clustered together (Figure 3). Meanwhile, the elovl8 clusters were subdivided into elovl8a and elovl8b branches, and elovl8a and elovl8b of the Chinese perch were closely related to corresponding paralogs in the Percomorpha (see more details in Figure 3).
3.4. Gene Structures and Genomic Synteny of Both elovl8 Genes in Teleost
A comparative analysis of gene structures can reveal their variability across various species. In this study, the elovl8a gene contains eight exons and seven introns in Chinese perch and zebrafish (Danio rerio), whereas the elovl8a gene of grass carp (Ctenopharyngodon idella), smallmouth bass (Micropterus dolomieu), leopard coralgrouper (Plectropomus leopardus), and giant grouper (Epinephelus lanceolatus) contains seven exons and six introns, but the European seabass (Dicentrarchus labrax) elovl8a gene only has six exons and five introns (Figure 4A). Differently, the structure of the elovl8b gene in Chinese perch was similar to that of those in climbing perch (Anabas testudineus), zebrafish, European seabass, greater amberjack (Seriola dumerili), and smallmouth bass; all of them have eight exons and seven introns (Figure 4B). Interestingly, the gene structures of elovl8a and elovl8b genes are different in smallmouth bass and European seabass, whereas they are consistent in Chinese perch (Figure 4).
In addition, comparative genomic synteny was obtained to investigate the evolutionary process of elovl8 genes in vertebrates. As detailed in Figure 5, the elovl8a is absent in the genome of mammals and Siluriformes fishes, whereas both elovl8a and elovl8b genes are present in Chinese perch, rabbitfish, European perch, and zebrafish. The genetic loci of elovl8a and elovl8b genes are significantly variable; however, two relatively conserved gene clusters, toe1-thap1-elovl8a-zswim5-urod and mutyh-elovl8b-glis1, are identified in the genome of these representative species.
3.5. Tissue Distribution Pattern of Both elovl8a and elovl8b Genes in Chinese Perch
Distribution patterns of both elovl8a and elovl8b genes were determined by qRT-PCR, and a total of 11 tissues (including brain, pyloric cecum, gill, gonad, heart, intestine, kidney, liver, muscle, spleen, and stomach) were detected in Chinese perch. Obviously, the elovl8a gene was extensively detectable in all measured tissues, with relatively higher expression levels in the liver and pyloric cecum (Figure 6A). Similarly, the elovl8b gene was also widely expressed in these measured tissues, with the highest expression level in gills, although no expression level was detected in the muscle (Figure 6B).
3.6. Effects of Different Nutritional Status on Transcriptional Changes of elovl8a and elovl8b Genes in Chinese Perch
In the long-term (18 d) food deprivation experiment of Chinese perch, we explored transcriptional changes of both elovl8a and elovl8b genes in the liver using qRT-PCR and also examined the expression pattern of both genes under different nutritional statuses. As shown in Figure 7, the transcription of elovl8a in Chinese perch was significantly decreased during the long-term starvation, while there was no significant change after refeeding compared with the starvation group (Figure 7A). In contrast, the expression of elovl8b did not change significantly during the long-term starvation compared with the control group, while it decreased significantly after refeeding compared with the control and starvation groups (Figure 7B).
3.7. Effects of Fish Oil and Vegetable Oil Administration on the Expression Levels of Both elovl8a and elovl8b Genes in Chinese Perch
Transcriptional changes of both elovl8a and elovl8b in response to intraperitoneal injection of fish oil (FO) and vegetable oil (VO) were detected. Our results showed that the transcription of both elovl8a and elovl8b was significantly increased at 0, 1, and 3 h after intraperitoneal injection of FO and VO in Chinese perch and then decreased at 6~48 h after injection with some variations (Figure 8). Interestingly, at 1, 3, 6, 12, 36, and 48 h after injection, transcription of elovl8a in the FO group was significantly higher than in the VO group (Figure 8A), and a similar pattern was observed for elovl8b at 1, 3, 6, 12, and 36 h after administration (Figure 8B).
3.8. Effects of Dietary Fish Oil and Vegetable Oil Supplementation on Both elovl8a and elovl8b Transcriptions in Chinese Perch
To investigate the transcriptional change patterns of both elovl8a and elovl8b genes in response to different dietary fish oil and vegetable oil supplementations, we detected their mRNA expression levels in the liver of Chinese perch. Our results showed that the hepatic elovl8a transcription level was significantly higher in fish fed with 2% FO supplementation diet than that of those fed with the control and with 2% VO supplementation diets (Figure 9A). A consistently similar pattern was observed for the elovl8b gene in Chinese perch (Figure 9B).
4. Discussion
LC-PUFAs are essential nutrients for several important physiological processes, including maintenance of cell membrane structure, energy metabolism, gene regulation, cell signaling, and immune function [31,32]. As a key rate-limiting enzyme in the endogenous biosynthesis of LC-PUFAs, ELOVL can extend the carbon chain of LC-PUFAs [10,20]. Several members of the ELOVL family (ELOVL1~7) have been extensively studied, while less is known about the molecular and functional characteristics of the new member ELOVL8 [10,17,33]. In our present study, two elongase genes (named as elovl8a and elovl8b) of Chinese perch were identified, and their ORFs were 810 and 789 bp, encoding 270 and 263 aa, respectively. The ORF length of Chinese perch elovl8a is different from that in rabbitfish [11], implying that elovl8a may be variable in teleosts. In contrast, the ORF of elovl8b showed high consistency among various teleosts, encoding approximately 263 aa, such as in rabbitfish [11], hybrid grouper [19], and yellow catfish [20]. These findings suggest that elovl8b is more conserved in comparison with elovl8a in teleosts.
Multiple protein sequence alignments revealed that both Elovl8a and Elovl8b of Chinese perch possess some ELOVL family features (Figure 1), including four conserved regions that are related to catalysis, six predicted a-transmembrane regions, a histidine box (HXXHH), and an endoplasmic reticulum (ER) retention signal. These data are consistent with several previous reports [11,15,19,20], indicating that Elovl8 proteins may be able to participate in LC-PUFA biosynthesis in teleosts. Subsequently, similar 3D structures of Elovl8 proteins were observed in several representative fish species (Figure 2), suggesting that both Elovl8a and Elovl8b are highly conserved with potentially similar functions [33].
Our phylogenetic analysis showed that the elovl8 gene family is closely related to but distinctly different from the elovl4 gene family (Figure 3), which is consistent with previous studies in yellow catfish, rabbitfish, ayu sweetfish, and threadfin fish [11,20,21,33]. Among them, elovl8 can be further divided into elovl8a and elovl8b subclades, and the Chinese perch elovl8a and elovl8b shared close relationships with banded archerfish elovl8a and striped bass elovl8b, respectively. Our findings are in line with those reports in previous studies [11,20], implying that two paralogs may have commonly existed in teleosts, and this phenomenon may be generated by the teleost-specific whole genome duplication event [11]. Meanwhile, both elovl8a and elovl8b of Chinese perch originated after zebrafish elovl8s, indicating that the evolutionary process of elovl8 genes is consistent with species evolution [27,33,34,35]. However, elovl8a has been lost in some fish species, such as yellow catfish, African catfish, channel catfish, and threadfin fish (Eleutheronema rhadinum); this phenomenon is probably due to functional redundancy between elovl8a and elovl8b isotypes early in evolution [16,20,33]. Meanwhile, the ability of Elovl8 to prolong LC-PUFAs was first reported in African catfish by using a yeast heterologous expression assay [16]. Consistently, a previous study [11] has shown that rabbitfish Elovl8b possesses an ability to extend C18 (18:2n- 6,18:3 n-3, and 18:4n-3) and C20 (20:4n-6 and 20:5n-3) PUFAs to long-chain fatty acids, whereas Elovl8a is not involved in such LC-PUFA synthesis. However, several recent studies reported that both Elovl8a and Elovl8b may play a potential role in fatty acid biosynthesis in zebrafish and ayu sweetfish [17,18]. In addition, previous studies suggested that elovl8 may be only found in teleosts, but recent reports revealed that elovl8 may also be present in some amphibians, reptiles, and other chordates as well [11,17]. Interestingly, a recent study [34] stated that elovl8-like genes were identified in echinoderms, implying a complex evolutionary history of this gene family in various animals.
Gene structure comparison revealed that the number of introns and exons of elovl8a genes are variable (Figure 4A), while the structure of elovl8b genes was more conserved (both existed in seven coding regions; Figure 4B), which is consistent with those findings in rabbitfish and yellow catfish [11,20]. These results suggest that Elovl8b may play more important roles than its paralog Elovl8a. Meanwhile, genomic synteny further indicated that both elovl8a and elovl8b genes are conserved in teleosts (Figure 5). Interestingly, the TESK2-TOE1 and ZSWIM5-UROD flanking genes were found in mammalian humans and mice, and both gene clusters are associated with the elovl8a gene; the flanking of the teleost elovl8b gene was also present, indicating that the elovl8 may have ever extensively existed in vertebrates, and two isoforms in teleosts may be due to the teleost-specific genome duplication event [20,36,37].
Tissue distribution patterns showed that both elovl8a and elovl8b genes were expressed in almost all examined tissues in Chinese perch (Figure 6). The elovl8a was mainly distributed in the liver, pyloric cecum, eye, and intestine (Figure 6A), whereas elovl8b was relatively high in the gill and pyloric cecum (Figure 6B), suggesting that the elovl8a and elovl8b genes in Chinese perch may play different physiological roles in the same tissues. These distribution patterns of Chinese perch, however, are different from those in rabbitfish and yellow catfish [11,20], implying that the distribution patterns of elovl8 genes may be species-specific, which could be caused by various inhabiting environments. These findings indicate that both elvol8a and elovl8b are involved in crucial physiological processes in various tissues, while functional differentiation may also exist between the two paralogs. Notably, both elovl8a and elovl8b have high expression in the liver, which may be due to the fact that the liver is the main organ for lipid metabolism and synthesis [11,20,33,38,39,40].
Previous studies have shown that different levels of ingested nutrients or different nutritional statuses can affect the biosynthesis of LC-PUFAs [39,41,42,43]. In Senegalese sole, refeeding significantly decreased the expression of hepatic Δ4fad and elovl5 genes after long-term starvation [40]. In yellow catfish, however, long-term starvation caused a significant decrease in hepatic fad6 gene expression with no significant change after refeeding [44]. In our present study, the Chinese perch hepatic elovl8a expression was significantly decreased after long-term starvation, with no significant change in expression after refeeding either (Figure 7A), indicating that long-term food deprivation may restrict the process of HUFA biosynthesis, but rescue with food supplements cannot obviously return the HUFA biosynthesis after this long-term food deprivation [44]. Differently, long-term fasting did not alter the haptic elovl8b expression, but refeeding significantly decreased its expression after refeeding (Figure 7B), implying that elovl8b may play important roles in maintaining normal physiological activity during starvation, and this is functionally different from elovl8a in Chinese perch.
Meanwhile, intraperitoneal injection experiments were conducted to further explore the functional traits of Elovl8s in Chinese perch. Our results showed that the expression levels of hepatic elovl8a and elovl8b genes were significantly increased at 0, 1, and 3 h after administration with FO and VO (Figure 8), suggesting that both FO and VO administration can stimulate the LC-PUFAs biosynthetic pathways in Chinese perch and thus compensate for the lower levels of essential LC-PUFAs in the commercial diet [20]. Besides, the expression pattern of the elovl8a gene (Figure 8A) was similar to that of the elovl8b gene (Figure 8B), implying that the elovl8a gene is also involved in the LC-PUFAs biosynthesis pathway in Chinese perch [17]. Notably, loss of the elovl8a gene in some teleost genomes is probably due to functional redundancy between elovl8a and elovl8b isotypes, such as in yellow catfish, African catfish, channel catfish, and threadfin fish [11,16,20,33]. Generally speaking, the variations in elovl8 in various teleosts may be due to the loss of genes or functions resulting from the overlapping of LC-PUFAs biosynthesis pathways in vivo as a result of the combined effects of different nutritional levels, feeding habits, and ecological habits [5,20,45].
Further in vivo experiments showed that the expression profiles of the Chinese perch hepatic elovl8a and elovl8b genes were regulated by dietary FO or VO supplementation (Figure 9), which is similar to those reports in rabbitfish and yellow catfish [11,20]. In our present study, dietary FO supplementation significantly improved the expression levels of both hepatic elovl8a and elovl8b genes, whereas dietary VO supplementation had no effect in Chinese perch (see Figure 9A,B). These findings are consistent with previous studies, which have shown that the elovl8 genes prefer to extend n-3 LC-PUFA substrates rather than n-6 LC-PUFA substrates in rabbitfish and African catfish [11,16]. In our current study, the FO diet contained a higher content of n-3 LC-PUFA substrates and thus significantly induced the transcription of elovl8 genes, suggesting that both elovl8 genes may have the ability to prolong LC-PUFAs in Chinese perch. Similar results were observed in zebrafish, since knockdown of its elovl8a and elovl8b genes resulted in accumulation of C20 LC-PUFAs in vivo and impaired synthesis of C22 LC-PUFAs [18].
5. Conclusions
In summary, two fatty acid elongase genes (elovl8a and elovl8b) were identified from Chinese perch in our present study. Bioinformatics analyses revealed that both elovl8 genes were highly conserved, implying their evolutionary and functional conservation in Chinese perch. Species-specific tissue distribution patterns suggest functional diversity of elovl8 genes in various teleosts. Meanwhile, different nutritional statuses and dietary fatty acid supplementation can affect the transcription of both elovl8 genes and thus regulate the LC-PUFA biosynthesis in Chinese perch. All in all, our findings provide novel insights into the evolutionary and functional properties of elovl8 genes in teleosts, as well as lay a solid foundation for a better understanding of the regulatory mechanisms of LC-PUFAs in vertebrates.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Castro L.F.C. Tocher D.R. Monroig O. Long-chain polyunsaturated fatty acid biosynthesis in chordates: Insights into the evolution of Fads and Elovl gene repertoire Prog. Lipid Res.201662254010.1016/j.plipres.2016.01.00126769304 · doi ↗ · pubmed ↗
- 2MonroigÓ. Lopes-Marques M. Navarro J.C. Hontoria F. Ruivo R. Santos M.M. Venkatesh B. Tocher D.R. Castro L.F.C. Evolutionary functional elaboration of the Elovl 2/5 gene family in chordates Sci. Rep.201662051010.1038/srep 2051026856376 PMC 4746653 · doi ↗ · pubmed ↗
- 3Oboh A. Kabeya N. Carmona-Antoñanzas G. Castro L.F.C. Dick J.R. Tocher D.R. Monroig O. Two alternative pathways for docosahexaenoic acid (DHA, 22: 6n-3) biosynthesis are widespread among teleost fish Sci. Rep.20177388910.1038/s 41598-017-04288-228634391 PMC 5478668 · doi ↗ · pubmed ↗
- 4Oboh A. Navarro J.C. Tocher D.R. Monroig O. Elongation of very long-chain (> C 24) fatty acids in Clarias gariepinus: Cloning, functional characterization and tissue expression of elovl 4 elongases Lipids 20175283784810.1007/s 11745-017-4289-328856549 PMC 5613102 · doi ↗ · pubmed ↗
- 5Keim S.A. Branum A.M. Dietary intake of polyunsaturated fatty acids and fish among US children 12-60 months of age Matern. Child Nutr.20151198799810.1111/mcn.1207724034437 PMC 4745899 · doi ↗ · pubmed ↗
- 6Sargent J.R. Tocher D.R. Bell J.G. The lipids Fish Nutrition Elsevier Amsterdam, The Netherlands 200318125710.1016/B 978-012319652-1/50005-7 · doi ↗
- 7Monroig O. Tocher D.R. Castro L.F.C. Polyunsaturated fatty acid biosynthesis and metabolism in fish Polyunsaturated Fatty Acid Metabolism Elsevier Amsterdam, The Netherlands 2018316010.1016/B 978-0-12-811230-4.00003-X · doi ↗
- 8Guillou H. Zadravec D. Martin P.G. Jacobsson A. The key roles of elongases and desaturases in mammalian fatty acid metabolism: Insights from transgenic mice Prog. Lipid Res.20104918619910.1016/j.plipres.2009.12.00220018209 · doi ↗ · pubmed ↗