The Cellular and Molecular Characteristics of Postnatal Human Thymus Stromal Stem Cells
Josipa Skelin Ilic, Ildikó Bódi, Lidija Milkovic, Zsolt Prodan, Dražen Belina, Darko Heckel, Lipa Cicin-Sain, Danka Grčević, Domenico Vittorio Delfino, Delfa Radic Kristo, Maja Matulić, Mariastefania Antica

TL;DR
This study identifies and characterizes stem cells in the human thymus that can form epithelial cells, revealing new insights into T-cell development.
Contribution
The study uncovers previously unknown characteristics of epithelial progenitor cells in the human thymus.
Findings
Neonatal and adult thymuses produce similar numbers of spheroids, indicating consistent progenitor potential.
Spheroids contain cells expressing genes of both mature and immature epithelial cells.
Treatment with 2-deoxyguanosine improved progenitor cell yield, and Cyr61 and Interleukin-22 affected spheroid size.
Abstract
Background: The thymus is the central hub of T-cell differentiation, where epithelial cells guide the process of their maturation. Objective: Our goal was to identify and describe progenitor cells within the human thymus that can differentiate into epithelial cells. Methods: When we plated enriched thymic cells in 3D culture conditions, rare individual cells capable of self-renewal and differentiation formed spheroids. Results: Both neonatal and adult thymuses produced similar numbers of spheroids, suggesting that progenitor potential remains consistent across age groups. Some cells within the spheres express genes typical of mature epithelial cells, while others express genes associated with the immature compartment active during thymic organogenesis. However, there were also cells expressing PDGFRβ. We treated the tissues with 2-deoxyguanosine before digestion, which improved the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
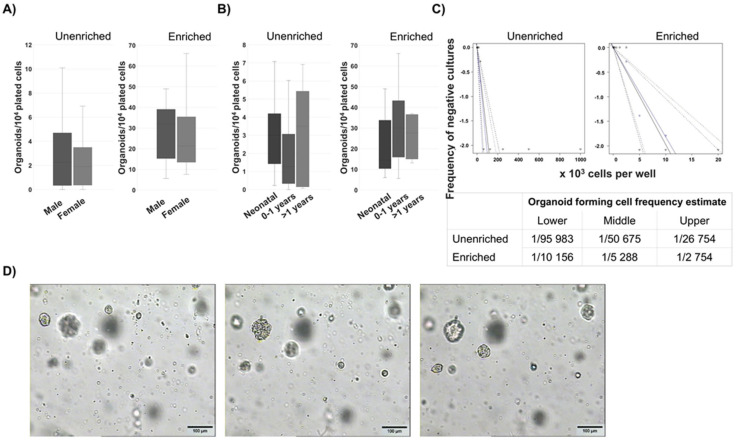 Figure 1
Figure 1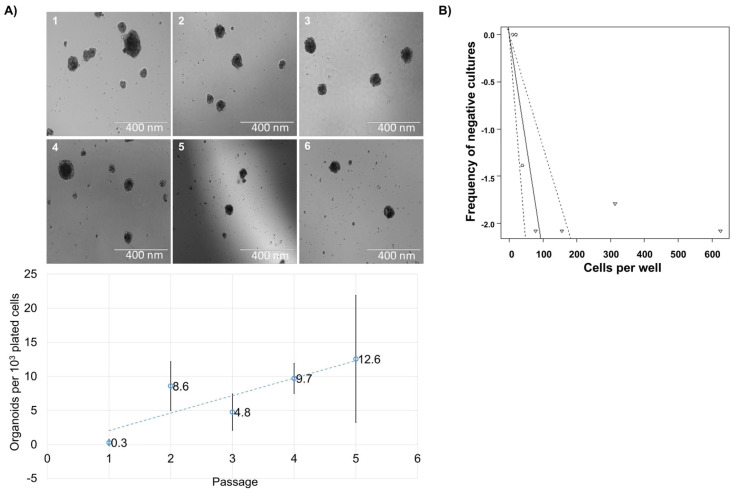 Figure 2
Figure 2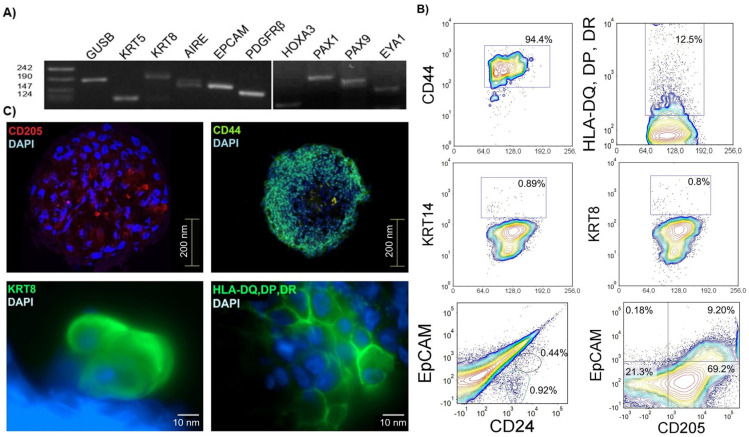 Figure 3
Figure 3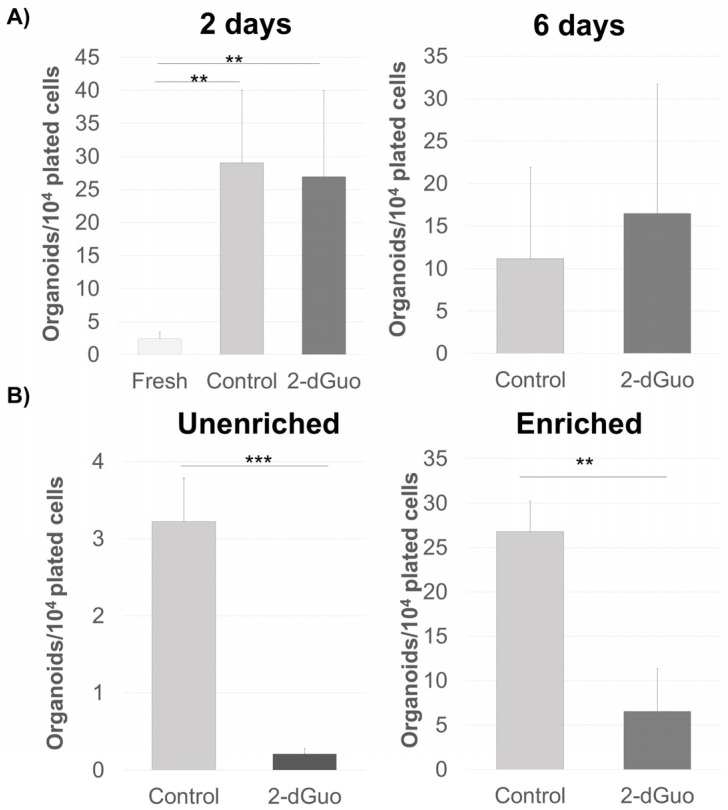 Figure 4
Figure 4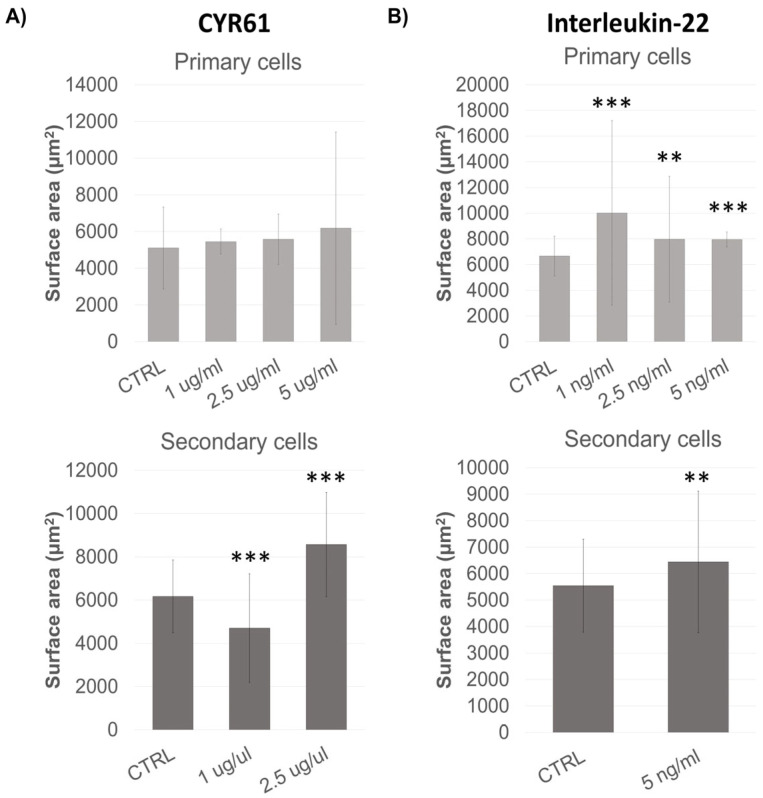 Figure 5
Figure 5- —Croatian Science Foundation
- —Seventh Framework program of the European Community for research and technological development (FP7)
- —bilateral program between the Republic of Croatia and the Peoples Republic of China
- —Terry Fox Zagreb run and Croatian League Against Cancer
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAdrenal Hormones and Disorders · Childhood Cancer Survivors' Quality of Life · Neuroblastoma Research and Treatments
1. Introduction
The adaptive immune system, the development, maturation and function of T lymphocytes, depends on the thymus. Thymic cortical (cTEC) and medullary (mTEC) epithelial cells, along with other cells of the stromal compartment, guide the differentiation and the selection of competent T lymphocytes through a complex multi-step process involving tests of functionality, sensitivity and self-tolerance [1,2]. With ageing, the thymus goes through a process of involution, during which epithelial cells are replaced by connective tissue, fibroblasts and adipocytes [3,4]. In addition to this natural process, the thymus is extremely sensitive to damage caused by chemo-, radio- or antibodytherapy. The degeneration of the stromal compartment inevitably leads to a loss in the number of naive T-cells produced and a decreased ability to fend off infections [5,6]. The search for an epithelial progenitor/stem cell capable of differentiating into both cortical and medullary epithelial cells needed for T-cell selection has been the focus of the field. Some groups have found that the adult thymus contains self-renewing, bipotent epithelial progenitors. However some studies dispute this, suggesting that the 3D spheroids originate from the mesenchymal lineage and do not support T-cell development [7,8,9]. Most of the published work focuses on the mouse model, which we cannot completely translate to humans, and both mouse and human thymic stromal progenitor cells remain to be precisely characterized. Here we show, on a substantial number of postnatal samples, that the human thymus, indeed, contains rare progenitor/stem cells with a spheroid-forming and self-renewal capacity, expressing genes involved in early thymic epithelial development as well as those typical for mature thymic epithelial cells.
2. Materials and Methods
2.1. Thymus Tissue
All of the thymuses used in experimental procedures were obtained from children who underwent a thymectomy as a part of the heart surgery at the University Hospital Centre Zagreb. The oldest patient from whom a thymus was obtained was 10 years old. However, we use the term “adult thymus”, as the thymus is considered to be fully developed right after birth in humans. Written informed consent was given by the children’s guardians in accordance with the Helsinki Declaration of 1975, as revised in 2008. The ethics committees from both the Rudjer Boskovic Institute (approval number BP-5865/1-2013) and the University Hospital Zagreb (Class: 8.1.-13/88-2, approval number: 02/21-BTB) have positively evaluated and approved this research.
2.2. Tissue Digestion and Thymic Stromal Cell Isolation
The thymuses were cleaned from blood clots and fatty tissue, weighed and chopped into small pieces. A single cell suspension was obtained by following a modified digestion protocol by Stoeckle [10]. Briefly, tissue fragments were digested during four incubation cycles with 1 mg/mL collagenase A (Worthington Biochemical Corporation, Lakewood, NJ, USA) and 0.1 mg/mL DNase I (Roche, Basel, Switzerland) in αMEM medium (Gibco, Thermo Fisher Scientific, Waltham, MA, USA), each lasting 15 min at 37 °C. Following the collagenase treatment, we incubated the tissue further, this time for 15 min with Versene (Lonza, Basel, Switzerland). As stem cells reside deep within the tissues, we used the cells obtained in the final digestion for further analysis. We counted the cells obtained from thymus digestion using trypan blue staining after each digestion cycle and collagenase change. The cells from the final digestion cycle (unenriched fraction), as well as the enriched cells after depletion and prior to FACS analysis, showed >95% viability in every thymus analyzed.
2.3. Thymic Stromal Cell Enrichment
The cells obtained during the final digestion cycle were washed and then labeled for 30 min with anti-human- CD3, CD4, CD8a, CD11b, CD14, CD16, CD19, CD56 (eBioscience, San Diego, CA, USA). The cells were then incubated for 30 min with Dynabeads goat anti-mouse IgG (Life Technologies, Carlsbad, CA, USA) and separated using a magnet. The obtained enriched stromal population of cells was used in all further experiments.
2.4. Spheroid Culture
The obtained enriched populations were grown in ultralow attachment flasks (Corning, New York, NY, USA) in a slightly modified medium for sphere cultivation, previously described by Ucar et al., 2014 [7]. αMEM (Gibco Thermo Fisher Scientific, Waltham, MA, USA) was supplemented with B27 supplement (Gibco, Thermo Fisher Scientific, Waltham, MA, USA), 0.5 µg/mL hydrocortisone (Sigma, St. Louis, MO, USA), 5 µg/mL insulin (Gibco, Thermo Fisher Scientific, Waltham, MA, USA), 4 µg/mL heparin (Sigma-Aldrich, St. Louis, MO, USA), 20 ng/mL bFGF (Peprotech, Thermo Fisher Scientific, Waltham, MA, USA) and 20 ng/mL EGF (Peprotech). The density of the plated cells was 10^5^ enriched cells/mL. We monitored the cultures for spheroid growth and fresh medium added every 3 days. The biological differences between samples were assessed by counting spheroids plated in quintuplicates in a 96-well plate 13 days after plating. The significance of the differences in sphere numbers between groups we determined by unpaired t-test or one-way ANOVA.
For the single-cell origin assay, enriched cells, obtained in the previously described manner, were mixed with cold (liquid) Mebiol^®^ gel (Cosmo Bio USA, Carlsbad, CA, USA) dissolved into the medium. An amount of 50 μL of the cell suspension was then plated in one well of a low attachment 96-well plate (Corning, New York, NY, USA) warmed to 37 °C. The temperature of the plate caused the gel to solidify and thus immobilize cells suspended in it. The gel was then overlaid with 150 μL of medium and the cells were cultured for 13 days prior to counting and photographing.
To assess the reformation ability of the sphere cells, we collected the spheres on a 20 µm strainer (pluriSelect, Leipzig, Germany) and dissociated them by incubating for 25 min with Versene (Lonza, Basel, Switzerland). The cells were then counted, replated in medium and monitored for reformation. The procedure was repeated every week as secondary cultures.
2.5. Growth Factor Treatment
The influence of Interleukin-22 (Il-22) and Cyr61 was assessed by adding the growth factors to primary and secondary spheroid cultures. Il-22 was used at 1, 2.5 and 5 ng/mL, while Cyr61 was used at 1 and 2.5 µg/mL. When used together, Il-22 was used at 5 ng/mL and Cyr61 at 2.5 µg/mL. Concentrations of 2 × 10^4^ of enriched cells (primary culture) or sphere cells (secondary cells) were plated in triplicates in 96-well plates. Spheroids were grown for 13 days when they were counted and photographed. Spheroid size was determined by ImageJ software (version 1.52a, National Institute of Health, Bethesda, MD, USA) [11]. The significance of the differences between treatments was determined by unpaired t-test or one-way ANOVA.
2.6. Limiting Dilution Assay
In order to determine the stem cell frequency in the thymus, as well as in the formed spheres, enriched cells (starting from 2 × 10^4^), unenriched cells (starting from 10^6^) and sphere cells (starting from 2 × 10^4^) were plated in quintuplicates in a two-fold series of dilutions, with dilutions positive for spheroids checked 13 days later. The series of dilutions were analyzed using the extreme limiting dilution analysis (ELDA) software (Version 4.7.1.2571 vom 16.01.2014) [12]. The interval of active cell frequency was calculated for at least three independent experiments.
2.7. 2′-Deoxyguanosine Treatment
We sought to optimize the stem cell isolation protocol and to simultaneously remove the bulk of thymocytes, so we utilized 2′-deoxyguanosine, previously known to selectively kill developing T-cells [13]. The thymic tissue was cleaned and prepared as previously described and then placed on a 40 μm filter (MiliporeSigma, Burlington, MA, USA) laid on a gelatinous sponge (Gelfoam, Pfizer, New York, NY, USA) immersed in αMEM medium (Gibco, Thermo Fisher Scientific, Waltham, MA, USA) containing 5% FCS (Gibco, Thermo Fisher Scientific, Waltham, MA, USA) and 1.35 mM 2′-deoxyguanosine (Sigma, St. Louis, MO, USA). A control tissue sample was incubated in the same manner but without 2′-deoxyguanosine. The tissue was incubated for 2 and 6 days, followed by digestion as described. The cells obtained by digestion were counted and their viability determined by using trypan blue staining. Although the cell viability obtained after digestion of fresh tissue was >95%, there was a decrease in viability after incubation, either with or without 2′-dGuo (consistently decreased to about 30% viable).
We plated the cells from the final digestion cycle at 2 × 10^4^ cells/mL and the number of spheres assessed 13 days later.
To see whether 2′-deoxyguanosine could hasten sphere formation and enhance their size, we incubated cells in spheroid medium as previously described, but with the addition of 1.35 mM 2′-deoxyguanosine, and analyzed for sphere formation and growth. The treated spheres and controls were counted 13 days after plating.
2.8. Multiplex PCR
Spheroids were collected 21 days after plating and lysed by snap freezing. RNA was transcribed into cDNA with transcript-specific primers by using MMLV reverse transcriptase (Applied Biosystems, Thermo Fisher Scientific, Waltham, MA, USA). The first PCR reaction was performed with a Taq polymerase (Sigma, St. Louis, MO, USA) during 15 cycles in a single tube with a mix of F1 and R primers, while the second PCR reaction was performed in separate tubes for each gene (F2 and R primers) for 42 cycles. The reaction was semi nested with common R primers. All of the primers were designed by Primer-BLAST and checked for specificity and dimer formation in silico and with known samples (Appendix A Table A1) [14]. At least one of the primers spans an exon–exon junction. The method was adapted for a small number of cells from the single cell multiplex PCR by Monteiro et al. (2004) [15,16].
2.9. Flow Cytometry
Spheroids were dissociated with Versene (Lonza Basel, Switzerland) and stained with mouse anti-human primary antibodies for 20 min, after which they were analyzed on FACSCalibur flow cytometer (Becton Dickinson BD, Franklin Lakes, NJ, USA). The antibodies used were antiCD44, (Tonbo Biosciences, San Diego, CA, USA), anti HLA-DQ, DP, DR (Becton Dickinson, BD), KRT8 (Becton Dickinson BD), KRT14 (Merck, Darmstadt, Germany), CD24 (eBioscience San Diego, CA, USA), EpCAM (Exbio, Praha, Czech Republic) and CD205 (eBioscience San Diego, CA, USA). The obtained results were processed with FlowJo version 10 (Becton Dickinson Company, Franklin Lakes, NJ, USA) and Weasel version 2.3 software, https://www.frankbattye.com.au/about.html.
2.10. Fluorescent and Confocal Microscopy
Spheroids were fixed and permeabilized with ice cold methanol (for intracellular antigens; KRT7/8) or BD Cytofix/Cytoperm (for surface antigens; CD205, CD44 and HLA DQ, DP, DR) for 20 min when unspecific binding was blocked for 20 min with Image-iT™ FX Signal Enhancer ReadyProbes™, Thermo Fisher Scientific, Waltham, MA, USA). After overnight incubation, the spheroids were stained with antibodies for an hour followed again by two washing cycles. The antibodies used were anti CD205 (eBioscience San Diego, CA, USA), rat anti-human CD44 (FITC, Tonbo Biosciences, San Diego, CA, USA), mouse anti-human KRT7/8 (Becton Dickinson, BD, Becton Dickinson BD, Franklin Lakes, NJ, USA) and mouse anti-human HLA-DQ, DP, DR (FITC, BD Becton Dickinson BD, Franklin Lakes, NJ, USA), while the secondary antibodies used Goat Anti-Mouse lgG Alexa Fluor™ 488 and Alexa Fluor™ 647 conjugates (Thermo Fisher Scientific, Waltham, MA, USA). Nuclei were stained with NucBlue™ Live ReadyProbes™ Reagent, Thermo Fisher Scientific, Waltham, MA, USA) for 30 min. Spheroids were suspended in 50 µL of PBS with 2% FCS and immobilized on glass slides by centrifugation.
3. Results
3.1. Spheroid Formation Potential of Cells from Human Thymus
Cells with spheroid forming potential were reliably isolated from human thymus samples and discernible spheroids were formed, on average, 7 days after plating. Based on the ability to form spheroids, the progenitor/stem cell presence was not limited by sex (Figure 1A) or age (from neonatal to 10 years of age) (Figure 1B) of the patient, as the spheroids were present in all of the samples analyzed, indicating a resident progenitor population. Interestingly, a larger variance in stem cell potential was found in the thymuses of patients older than one year. The stem cell frequency was determined by limiting dilution assay and it was estimated to be one in 50,675 unenriched thymic cells and one in 5288 enriched cells. These data confirm our method of stem cell enrichment (Figure 1C). Culturing cells in a semi-solid medium which inhibited cell aggregation yielded spheroids and showed that they were, indeed, derived from single cells (Figure 1D).
3.2. Reformation Ability of Cells from Human Thymus
One of the main prerequisites of classifying a cell as a progenitor or precursor is the ability to self-renew. To test this ability, we enzymatically dissociated spheroids and replated the obtained cells. In all examined instances, cells reformed new spheroids (Figure 2A). This dissociation was repeated up to six times (up to the 7th spheroid) but it was noticed that the total cell number declined. This observation led us to believe that not all of the cells within the spheroid retain the true self-renewal capacity of progenitor/stem cells. We performed a limiting dilution assay with cells from dissociated primary spheroids, and found that, in secondary spheroids, one in every 43.2 sphere cells has the capability of reformation (Figure 2B). Even though the cell numbers declined with reformation, the number of spheres per 10^3^ cells plated increased, albeit irregularly (Figure 2). We postulate that the cells formed by the division of the initial spheroid-forming cell are likely directed towards terminal differentiation and thus lose the ability to divide independently of cell–cell or cell–matrix contact.
3.3. Expressional Analysis of Thymus Spheroid Cells
Deducing from cell enrichment and limiting dilution experiments, we hypothesized that the spheroid-initiating cells reside within the non-hematopoietic population of the thymus. To determine their precise identity, we reverse transcribed and amplified mRNA from spheroids by multiplex PCR [16], thus insuring amplification specificity from limited material (Figure 3A). Spheroids proved positive for genes vital for thymus function. The expression of EpCAM confirmed the presence of differentiated epithelial cells. Specific keratins show the presence of medullary (KRT5) and cortical (KRT8) thymic epithelial cells [17,18]. We also detected the expression of AIRE, a transcription factor found to be fundamental for the proper expression of tissue-specific antigens by medullary epithelial cells [19,20]. We also analyzed the expression of FOXN1 in spheroid samples from different thymuses and have indeed detected the expression of FOXN1. However, the expression was not consistent in all samples. To check whether mesenchymal cells contribute to spheroids, we analyzed the expression of the platelet-derived growth factor receptor beta (PDGFRβ), found on a variety of cells but mostly on those of mesenchymal origin [21]. Interestingly, some spheroids were positive for PDGFRβ.
Studies on mouse thymic organogenesis have provided insight into the molecular mechanisms involved and have defined the genes indispensable for the proper development and organization of the thymic stromal compartment—HOXA3, PAX1, PAX9 and EYA1. The expression of these genes has been found to be conserved between mice and humans, with PAX1 having an even more restricted expression pattern in humans [22]. All of the aforementioned genes were found to be expressed in thymic spheroids obtained from the postnatal human thymus. These data strongly suggest that a population of immature cells should be capable of maintaining thymic stroma.
The protein expression was analyzed by flow cytometry (Figure 3B) and by confocal and fluorescence microscopy (Figure 3C). It was shown that the spheroid cells are positive for CD44, a glycoprotein expressed on a wide variety of cells. HLA-DQ, DP, and DR, detected by the common antibody for MHC II molecules and expressed on antigen presenting cells, were found to be present on 12.5% of spheroid cells [23]. Mature thymic epithelial cells were detected by KRT8 (cTEC) and KRT14 (mTEC) and were found to compose <1% of spheroid cells [18]. Cells positive for KRT8 were also found by fluorescent microscopy. CD205 was found to be expressed on 80% of the spheroid cells. Flow cytometry revealed that it was expressed with EpCAM (9.3%) and CD24 (9.4%). The two populations were mutually exclusive. EpCAM was positive on a small population of cells, confirming that the mature thymic epithelial cells represented a minority of spheroid cells. CD24 was considered as a possible specific progenitor marker due to its expression by human intestinal epithelial stem cells as well as by a number of cancer stem cells [24,25]. CD24 was found on 1.4% of spheroid cells.
3.4. 2-Deoxyguanosine Treatment of the Stromal Cell Population
To enrich the stromal cell population, and consequently the population of spheroid-forming cells, we utilized the property of 2′-deoxyguanosine (2-dGuo) to selectively kill T lymphocytes [13,26]. As the goal of different treatments was to increase the number of progenitors, our final readout was the number of spheroids formed under each treatment condition, as shown in Figure 4 for 2-dGuo. Shorter tissue incubation (2 days) with 1.35 mM 2-dGuo led to a significant increase in the number of spheroids formed in the final digestion cycle when compared with fresh tissue, but their number was comparable to those of cells grown without 2-dGuo. Extending the incubation period to six days led to a moderate increase in the number of spheroids formed, possible because of the tissue degradation during such prolonged incubation (Figure 4A). The outcome of the spheroid formation was also found to be dependent on the patient sample. We sought to ascertain whether this effect was due, as suspected, to the depletion of thymocytes from the tissue leading to the increased percentage of stromal cells in the final fraction or due to a stimulatory influence of 2-dGuo. We plated unenriched cells obtained from fresh tissue in spheroid medium with the addition of 1.35 mM 2-dGuo. We found that the direct presence of 2-dGuo in the medium inhibited spheroid formation (an 11- to 21.8-fold decrease in spheroid number) (Figure 4B). Therefore, the continuous exposure of cells to 2-dGuo had an unfavorable effect on the proliferative capabilities of the precursors. It cannot be excluded that cell debris from apoptotic T lymphocytes had an adverse effect on the growth of other cells in the culture.
3.5. Cyr61 and Interleukin-22 Impact on Spheroid Formation
To enhance the culture conditions of spheroid-forming cells, we cultured primary thymus stromal cells and dissociated spheroids (secondary cells) with a factor discovered to be implicated in the thymus epithelial cell expansion in mice, Cysteine-rich protein 61 (Cyr61) [27]. We also tested a cytokine which has been known to positively influence the proliferative capabilities of a variety of tissue-resident stem cells, Interleukin-22 (Il-22) [28,29,30].
We found that Cyr61 boosts the rate of spheroid cell divisions and influences the size redistribution in favor of larger spheroids, but these effects seem to be dependent on the patient sample. Cyr61 at 2.5 μg/mL impacted the proliferation of dissociated spheroid cells, but only in some samples and therefore was not found to be significant (Figure 5A).
The treatment of primary thymus cells with Il-22 (1 and 2.5 ng/mL) increased spheroid number and size, but only in some patient samples (Figure 5B).
When cells were treated simultaneously with 2.5 μg/mL Cyr61 and 2.5 ng/mL Il-22, no significant effect on spheroid size or number was seen.
4. Discussion
Up to today there have been many different attempts to establish thymus-like conditions for T-cell differentiation. The first models used immunocompromised mice engrafted with human fetal thymus and liver tissue fragments and in vitro models with human–mouse chimeric fetal thymic organ cultures [31,32,33]. Montel-Hagen [34] generated artificial thymic spheroids (ATO) from human and murine hematopoietic stem and progenitor cells cultivated with murine stromal cells expressing Notch ligand. Many of the established models needed feeder layers or to prove their functionality in vivo.
In parallel, there were many attempts to cultivate and differentiate thymic stem cells which could reconstitute the thymus conditions in vitro. Most of these experiments were undertaken on murine thymic tissue. Bleul et al. established functional thymus initiated by a postnatal epithelial progenitor cells [35]. Furthermore, the existence of a bipotent epithelial progenitor population was identified in the murine adult thymus, and is capable of differentiation into both cortical and medullar epithelial cells. The identified progenitor cells were EpCAM and FOXN1+ [36,37]. Ocampo-Godinez et al. [38] established the culture of functional TECs and thymic interstitial cells from mice by cultivating them in DMEM/F-12 medium with 20% FBS. These cells were able to form thymospheres and expressed functional markers for T-cell development, such as Aire, Il25, CD80 and CCL21. Lim et al. [39] isolated functional thymic epithelial spheroids from adult murine thymus. These consisted of epithelial cells and were able to proliferate in a complex medium sustaining Wnt- and EGF-dependent signaling. They contained epithelial progenitors, which could be differentiated either in cTEC or mTEC. Hubscher et al. [40] managed to grow mouse TEC spheroids, which mediate T-cell development. They grew cells in defined spheroid-based medium on matrix-based hydrogel with the addition of FGF7. The culture conditions allowed spheroid formation from neonatal, but not adult TECs. On the other hand, there were numerous experiments in which thymic cells were differentiated from human and mouse embryonic stem cells or induced pluripotent stem cells. Additionally, iTEC cell-induced thymic epithelial cells were produced by direct reprogramming of mouse fibroblasts by FOXN1 [41]. Tetteh et al. [42] showed that neural crest-derived mesenchymal cells supported thymic reconstitution after lethal irradiation in mice.
Recently Campinoti et al. [8] identified epithelial cells in human thymuses which had the epithelial–mesenchymal phenotype and were capable of long-term proliferation and establishing a functional microenvironment for human T-cell differentiation. However, there are still different hypotheses about the origin of thymic stem cells, whether they are of epithelial or mesenchymal origin, and, accordingly, their markers. For these reasons, we approached the issue from a different angle and, considering the unknown phenotype of precursors, removed hematopoietic cells. Our efforts at improving the isolation and culture technique with 2-deoxyguanosine, Interleukin-22 and Cyr-61 did not provide definitive results. Tissue treatment with 2-dGuo yielded a greater proportion of stromal cells within the population obtained by digestion, but incubation is a lengthy process and the addition of 2-dGuo to the cell medium had a negative effect on spheroid formation. Interestingly, interleukin-22 and Cyr61 treatment had highly individualized effects. It could be possible that the human thymus contains several different types of stem/progenitor cell populations with different responses to the compounds used. Thus, the differing effects could be due to the varying proportions of spheroids derived from different stem cells.
By growing cells of the postnatal human thymus in low attachment conditions with the addition of growth factors [7], we continuously obtained 3D clusters of cells i.e., spheres or spheroids. Due to the presence of cells with phenotypic markers peculiar to mesenchymal cells, which are known for their high migratory potential, we needed to test whether the cells were of single-cell origin or cell aggregates by immobilizing them in a semi-liquid gel. However, more experiments are needed to completely prove that there was no cell migration present in our experimental conditions and test the proliferative activity of the cells in culture. Presuming that each sphere was formed from a single stem cell, the stem cell potential of postnatal human thymuses was shown to be independent of the age or sex of the patient. However, the differences between individual samples became bigger in samples from patients older than one year. The limited dilution assay shows that this sphere-forming population is rare, as stem cells usually are. A population of cells within the clusters has also been shown to possess the capacity for self-renewal. However, the cells that originated from the sphere-forming progenitors did not always retain self-renewal properties. Nonetheless, these findings together indicate the existence of stromal progenitors of the human thymus. Cells from the clusters express a plethora of markers specific for early (HOXA3, EXA1, PAX1 and PAX9) and late (EpCAM, KRT7/8, KRT14) thymic epithelial differentiation and epithelial cell function (HLA-DQ, DP, DR and AIRE) [18,20,22,23]. HOXA3 is expressed in the third pharyngeal pouch during embryonic development and its deletion leads to the inability of the pouch to differentiate into the thymus and parathyroid glands [43]. EYA1 is expressed in the same context and with the same consequences as HOXA3, while PAX1 and PAX9 continue the signaling cascade during thymic organogenesis [44]. The expression of these early genes reveals the presence of an extremely immature population of cells within the postnatal thymus. The expression of functionally important genes, such as HLA-DQ, DP, DR and AIRE, shows that some of the cells within the clusters have differentiated into competent keratinocytes. C205, which we found to be present on spheroid cells, is also expressed on antigen presenting cells, more specifically, on cortical epithelial cells and dendritic cells of the thymus [45]. The abundance of CD205 cells in our thymus cultures may suggest that the spheroids are predominantly cortical epithelium rather than medulla epithelium. We cannot rule out the possibility that our culture conditions favor cTEC development over mTEC development. Interestingly, CD44 and PDGFRβ, the markers more commonly connected to mesenchymal cells, were found to be abundantly present in the clusters [46]. CD24, a glycoprotein found on mesenchymal bone marrow stem cells, but also on intestinal epithelial stem cells and hematopoietic cells, was found on a small population within the clusters [47]. Taken together, our results are in accordance with those obtained by Campinoti et al. [8], showing that the human thymus contains epithelial–mesenchymal hybrid cells capable of expansion and differentiation into functional thymic stromal cells. We chose to call our clusters spheroids because of the heterogeneity shown. In view of the possibility that the cells’ presumed progenitors have a mixed epithelial–mesenchymal phenotype, it would be interesting to observe the localization of signals from epithelial and mesenchymal markers in spheroids simultaneously. Taking into account the differential response of spheroid-forming cells to Cyr61 and Il-22, and differences in spheroid-forming potential between patient samples, we propose the existence of two distinct populations of tissue-resident progenitor/stem cells in the postnatal human thymus. Their distinctions, specificities and precise roles in thymic stromal maintenance are yet to be determined. The future of the research described in this paper surely lies in a single cell approach and cell tracking, including the precise analysis of signals within the stem cell. In this paper we have described the very robust method that we used and the cells we obtained. The population we isolated and described here is ideal for a more thorough examination with antibodies to markers of mesenchymal phenotype, and this is the matter of our future work.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Boyd R.L. Tucek C.L. Godfrey D.I. Izon D.J. Wilson T.J. Davidson N.J. Bean A.G.D. Ladyman H.M. Ritter M.A. Hugo P. The Thymic Microenvironment Immunol. Today 19931444545910.1016/0167-5699(93)90248-J 8216723 · doi ↗ · pubmed ↗
- 2Kumar B.V. Connors T.J. Farber D.L. Human T Cell Development, Localization, and Function throughout Life Immunity 20184820221310.1016/j.immuni.2018.01.00729466753 PMC 5826622 · doi ↗ · pubmed ↗
- 3Steinmann G.G. Klaus B. Muller-Hermelink H.K. The Involution of the Ageing Human Thymic Epithelium Is Independent of Puberty. A Morphometric Study Scand. J. Immunol.19852256357510.1111/j.1365-3083.1985.tb 01916.x 4081647 · doi ↗ · pubmed ↗
- 4Palmer S. Albergante L. Blackburn C.C. Newman T.J. Thymic Involution and Rising Disease Incidence with Age Proc. Natl. Acad. Sci. USA 20181151883188810.1073/pnas.171447811529432166 PMC 5828591 · doi ↗ · pubmed ↗
- 5Nijhuis E.W. Nagelkerken L. Age-Related Changes in Immune Reactivity: The Influence of Intrinsic Defects and of a Changed Composition of the CD 4+ T Cell Compartment Exp. Clin. Immunogenet.199291952021364007 · pubmed ↗
- 6Chaudhry M.S. Velardi E. Dudakov J.A. van den Brink M.R.M. Thymus: The Next (Re)Generation Immunol. Rev.2016271567110.1111/imr.1241827088907 PMC 4837659 · doi ↗ · pubmed ↗
- 7Ucar A. Ucar O. Klug P. Matt S. Brunk F. Hofmann T.G. Kyewski B. Adult Thymus Contains Fox N 1- Epithelial Stem Cells That Are Bipotent for Medullary and Cortical Thymic Epithelial Lineages Immunity 20144125726910.1016/j.immuni.2014.07.00525148026 PMC 4148705 · doi ↗ · pubmed ↗
- 8Campinoti S. Gjinovci A. Ragazzini R. Zanieri L. Ariza-Mc Naughton L. Catucci M. Boeing S. Park J.E. Hutchinson J.C. Muñoz-Ruiz M. Reconstitution of a Functional Human Thymus by Postnatal Stromal Progenitor Cells and Natural Whole-Organ Scaffolds Nat. Commun.202011637210.1038/s 41467-020-20082-733311516 PMC 7732825 · doi ↗ · pubmed ↗