Toxoplasmosis in Sheep Caused by Toxoplasma gondii Clonal Type I
Yurong Yang, Yiheng Ma, Kai Quan, Bingyan Guo, Yibao Jiang

TL;DR
This study reports the first isolation of a Toxoplasma gondii Type I strain from sheep in China, which was found to be non-lethal in mice.
Contribution
The first isolation and characterization of a viable T. gondii Type I strain from sheep with avirulence in mice.
Findings
A T. gondii Type I strain (TgSheepCHn15) was isolated from sheep tissues in China.
The isolated strain was avirulent in Swiss mice, with a survival time of 97 ± 31 days.
The overall T. gondii prevalence in sheep was 16.4% based on PCR testing.
Abstract
Toxoplasma gondii infects many warm-blooded animals, including sheep. Humans can be infected with T. gondii by consuming undercooked sheep tissues containing cysts. Type II (ToxoDB #3 or #1) and Type III (ToxoDB #2) varieties are the epidemic genotypes of viable T. gondii strains in sheep. Until now, no viable T. gondii Type I (ToxoDB #10) strain has been found in sheep tissues. Type I is usually associated with the highly virulent strain in mice. A viable T. gondii strain (TgSheepCHn15), ToxoDB #10, was isolated from the tissues of sheep at a veterinary clinic. This is the first report of a viable T. gondii clonal type I genotype that correlates with avirulence in mice. Ovine toxoplasmosis has a significant negative impact on sheep farming, and abortion is the main clinical manifestation. The objective of the present study was to survey ovine toxoplasmosis and assess its potential…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
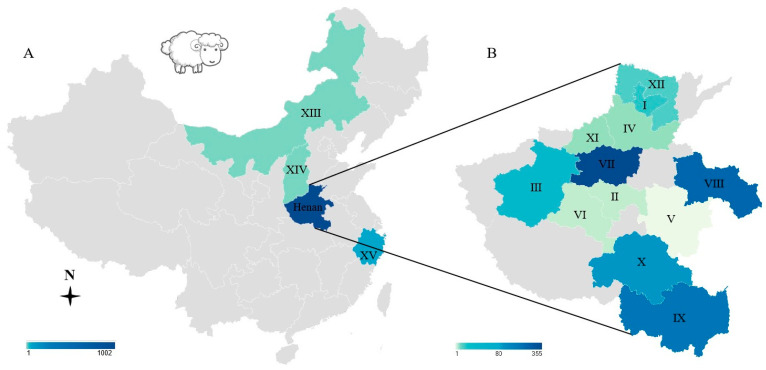 Figure 1
Figure 1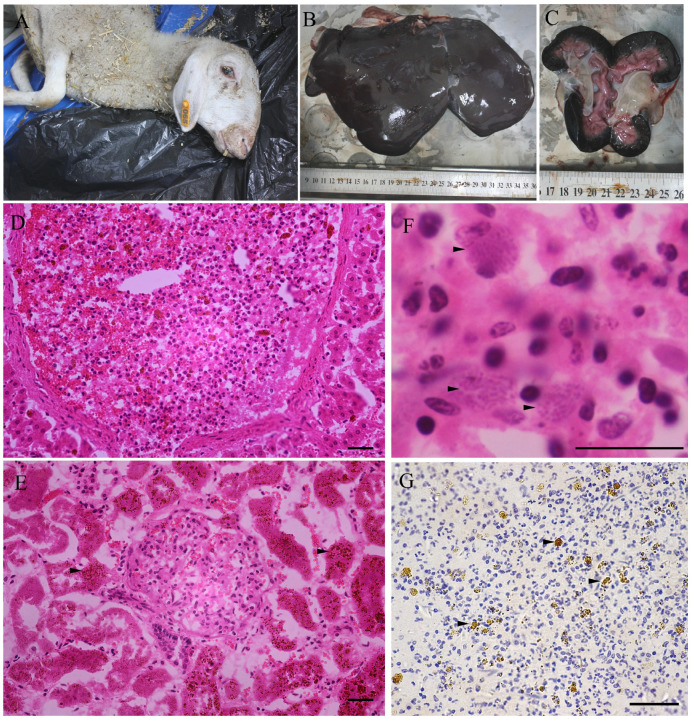 Figure 2
Figure 2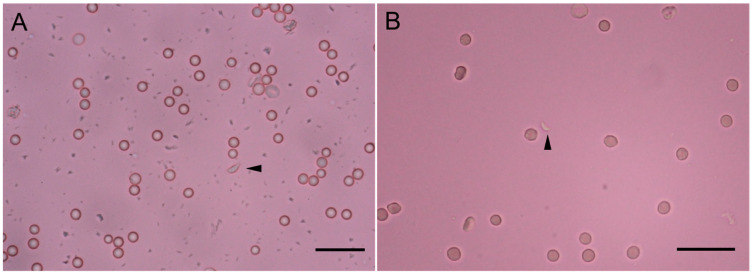 Figure 3
Figure 3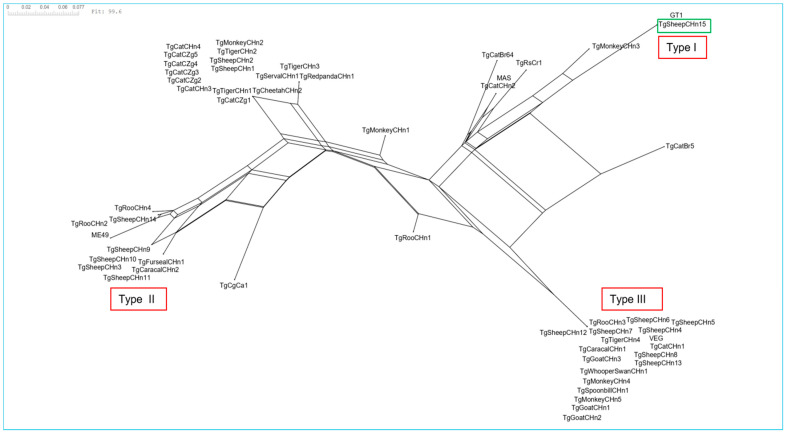 Figure 4
Figure 4- —Henan Province modern agricultural industrial technology system
- —Agriculture in Henan Province for fine breed research project
- —Henan Province International Scientific and Technological Cooperation Projects
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsToxoplasma gondii Research Studies · Parasitic Infections and Diagnostics · Rabies epidemiology and control
1. Introduction
Toxoplasma gondii infects many warm-blooded animals, including sheep. Ovine toxoplasmosis can cause serious reproductive disorders worldwide [1]. Studies have found that ovine toxoplasmosis infection routes include horizontal transmission (ingestion oocysts) and vertical transmission (exogenous or endogenous transplacental transmission) [2,3,4]. Hence, ovine toxoplasmosis can be preserved in sheep herd generations without definitive hosts (felids) [2,4].
In sheep worldwide, Type II (ToxoDB #3 or #1, 63%) and Type III (ToxoDB #2, 9%) are the epidemic genotypes of viable T. gondii strains [5]. Until now, no viable T. gondii Type I (ToxoDB #10) strain has been found in sheep tissues. Usually, Type I is associated with a highly virulent mouse strain, such as the RH strain or GT1 strain [6,7]. In this study, the T. gondii clonal type I genotype that correlates with avirulence in mice was found in sheep. China currently ranks the highest in sheep and goat meat production (animal production: 5166 kilotons in 2022). However, there has been limited research on ovine toxoplasmosis. The objective of the present study was to survey ovine toxoplasmosis and assess its potential harm to the host.
2. Materials and Methods
2.1. Information on Samples and Sources
From 2018 to 2024, samples from 1035 sheep were obtained from Henan, Shanxi, Inner Mongolia, and Zhejiang provinces (Figure 1, Table 1). Samples included tissues (84 myocardial tissues from the slaughterhouses; the heart, liver, spleen, lungs, kidney, and lymphonodus of 12 lambs from the slaughterhouses; the heart, liver, spleen, lungs, kidney, and lymphonodus of 38 dead sheep or aborted fetuses from veterinary clinics) from 134 sheep, as well as heart juice (n = 134), sera (528 sera from the farms, 323 sera from the slaughterhouses, 20 sera from veterinary clinics, n = 871) and blood (n = 30, from veterinary clinics) from 1035 sheep. The samples were sent to the laboratory (Veterinary Pathology, Henan Agricultural University, Zhengzhou, Henan Province, China) for etiological diagnosis, the evaluation of meat quality, and to check the Brucella spp. infection status. The sheep were divided into three groups according to the sample source (farms, veterinary clinics, and slaughterhouses). For the supervision of Brucella spp. infection status, we only obtained serum samples from alive sheep in farms, with ages ranging from 1 month to 6 years old; these samples were defined as having a “farm source”. For meat quality evaluation, tissues or serum samples were obtained from slaughterhouses using animals ranging in age from 8 to 10 months and showing good clinical health. For etiological diagnosis, tissues or serum samples were collected from sick sheep, or aborted fetuses, or alive sheep with the clinical suspicion of disease, with the age of animals ranging from 0 days to 6 years old; these samples were defined as having a “veterinary clinics source”. The survey of ovine toxoplasmosis in these sheep was also allowed. These samples were not collected according to the epidemiological survey software; they were scattered.
2.2. Detection of T. gondii Antibodies in Sheep
For T. gondii, the postmortem detection of antibodies in meat juice samples is a useful alternative to serum examination [8,9]. In this study, for samples from slaughterhouses or dead cases, blood was removed from each heart and centrifuged to obtain the supernatant, which was termed “heart juice”. Heart juice (n = 134) and serum samples (n = 901) were tested for T. gondii IgG antibodies by a modified agglutination test (MAT) [10,11,12]. Titers ≥ 1:100 indicated T. gondii infection, as compared to viable parasite isolation from sheep tissues [11,12,13]. Formalin-fixed T. gondii RH tachyzoites were provided by the University of Tennessee (Knoxville, TN, USA). *T. gondii-*infected mice serum was used as a reference.
2.3. T. gondii Nucleic Acid Detection in Sheep
DNA was extracted from blood (30 sheep) or tissue samples (134 sheep) by a DNA extraction kit (Tiangen Biotec Company, Beijing, China, DP304). T. gondii was examined using the primer pair TOX-5 (5′-CGC TGC AGA CAC AGT GCA TCT GGA TT-3′) and TOX-8 (5′-CCC AGC TGC GTC TGT CGG GAT-3′) [14]. Target DNA was amplified using the following program: initial denaturation at 94 °C for 5 min, denaturation at 94 °C for 1 min, annealing at 60 °C for 1 min, and extension at 72 °C for 1 min. Following 35 cycles, a final extension step of 10 min was added at 72 °C. The PCR product was 450 bp, and negative (negative samples were from Toxoplasma gondii nucleic acid detection kit, Product ID: 25T, Shanghai Yan Qi Biological Technology Company, Shanghai, China) and positive (VEG strain infected mice brain was provided by Dr. Dubey) [15] controls were included in every batch.
2.4. Morphological Observation of T. gondii Parasites in Sheep Tissues
Tissue samples (only myocardial tissues from 84 sheep; heart, liver, spleen, lungs, kidney, and lymphonodus of 12 lambs; heart, liver, spleen, lungs, kidney, and lymphonodus of 38 dead sheep or aborted fetuses) from 134 sheep were fixed using formalin, embedded in paraffin, stained with hematoxylin and eosin, and subjected to immunohistochemistry (IHC) for T. gondii [1,16]. T. gondii-infected mouse tissues and T. gondii-free mouse tissues were used as positive and negative controls, respectively.
2.5. Isolation Viable T. gondii from Tissues of Sheep
Mouse bioassays were performed on blood or tissue samples of sheep (Table 1) . From 134 and 30 blood samples, 22 tissues and 2 blood samples were selected to isolate T. gondii by a mouse bioassay. Via a mouse bioassay, we used serologies or etiologies positive for T. gondii as selection references for most samples. Some samples with serologies or etiologies negative for T. gondii were also selected to isolate viable strains.
In brief, tissue (striated muscles or tissues containing T. gondii DNA, 50 g) was ground or pepsin-digested and used to inoculate Swiss mice (n = 2–5) or a C57BL/6 IFN-γ^−/−^ mouse subcutaneously [1]. Clinical signs were observed. T. gondii parasites were examined in the lungs, mesenteric lymph nodes, pleural effusion, ascites, or the brain of dead mice. Facial vein blood was obtained from mice 30 days post-inoculation (DPI) and 60 DPI, and 1:100 dilutions of serum were checked for T. gondii antibodies by MAT. If no antibodies and parasites were found, the homogenized mouse lungs, brain, spleen, and heart were subpassaged into a new mouse group subcutaneously. Only one round of mice was inoculated with negative samples.
2.6. Cell Cultivation and Genotyping
The mouse lungs were seeded into Vero cells (ThermoFisher, Waltham, MA, USA). DNA was extracted from T. gondii-infected mice tissues. Then, PCR-RFLP was used to check the genotype of the T. gondii isolate using 10 genetic markers [17]. The virulent genes ROP18 and ROP5 were also typified [18]. Eight T. gondii DNA (obtained from Dr. Chunlei Su, University of Tennessee) samples were used for reference.
2.7. Statistical Analysis
Data are expressed as the mean ± SEM using GraphPad Prism 8.0, followed by one-way ANOVA, with p < 0.05 considered to be statistically significant. Statistical analyses relating to organelles in the isolated T. gondii strain were performed using transmission electron microscopy (TEM). T. gondii genotype analysis was performed by SplitsTree CE 6.0.0 [19]. The original input consisted of 56 taxa and 57 standard sequences of length 62. The Show Splits method (default options) was used to obtain Split Network visualization. To explore the phylogenetic position of the strain studied, strains (n = 48) from Central China were selected, with the background of the strains shown in Table S1.
3. Results
3.1. Investigation of Ovine Toxoplasmosis in Sheep
In this survey, we checked heart juice (n = 134) or sera (n = 901) from 1035 sheep for T. gondii IgG antibodies by MAT. Overall, 7.2% (75/1035) (95% confidence interval 5.81–9.00%) of sheep had T. gondii antibodies (Table 1 and Figure 1). The titers of these samples ranged from 1:100 to 1:12,800, with a titer of 1:100 in 12 sheep, 1:200 in 20, 1:400 in 2, 1:800 in 7, 1:1600 in 15, 1:3200 in 11, 1:6400 in 5, and 1:12800 in 3.
The sheep were divided into three groups according to the sample source. Namely, farms, veterinary clinics, and slaughterhouses. The T. gondii prevalence in sheep was 9.8% (52/528, 95% CI: 7.57–12.70%) in farms, 7.1% (12/169, 95% CI: 4.00–12.11%) in veterinary clinics, and 3.3% (11/338, 95% CI: 1.76–5.80%) in slaughterhouses. Compared with sheep in slaughterhouses, the prevalence of T. gondii was higher in farms or veterinary clinics (p < 0.05), with an odds ratio of 3.248 or 2.272 (Table 1 and Table 2).
The prevalence of T. gondii in sheep was higher in the <1-year group (16.1%, 61/378) compared to the >1-year group (2.1%, 14/657) (p < 0.0001). When analyzed by geographic location, there were few samples from Inner Mongolia (n = 1), Shanxi (n = 1), and Zhejiang (n = 31), and none of them showed seroprevalence for T. gondii (Table 1 and Table 2).
According to the PCR results, T. gondii DNA was detected in 27 of 164 sheep (16.4%, 95% CI: 11.52–22.94%). These included whole blood (n = 11), multiple organs of two ewes, three lambs, three aborted fetuses, and one placenta from veterinary clinics; myocardium samples (n = 7) from slaughterhouses; and none of the sheep from farms (Table 1).
3.2. Assessment of Pathological Lesions, and T. gondii Antigen Distribution
Tissue samples were obtained from 134 sheep between 2018 and 2024 (Table 1). TgSheepCHn15 was isolated from the tissue of a ewe (Xinxiang, Henan, 11 October 2021) (Figure 2). The ewe had a moderate appetite, emaciation, and anemia, could ruminate, and died the next day. Necropsy showed that the lungs had adhered to the wall of the chest, with pulmonary congestion and white foam observed in the trachea. The liver was swollen, crisp, and dark purple in color, and the gallbladder was enlarged and full of bile. The spleen was mildly enlarged, soft, and black in color. The kidneys were atrophic and black, and the fat was yellow and jelly-like. The mesenteric lymph nodes were enlarged with striated black spots on the cut surface. Microscopically, the interlobular boundaries of the liver were unclear, and the hepatic cords were arranged in a disordered manner. The hepatic veins were filled with neutrophils. Necrotizing lymphadenitis was observed, with many macrophages in the lymph nodes with T. gondii-like parasites. T. gondii parasites were verified in this ewe by immunohistochemistry (Figure 2). Interstitial pneumonia and necrotizing splenitis were observed. Necrotizing atrophic renal tubular nephritis and huge hemoglobin deposits were observed in renal tubular epithelial cells. Pathological changes suggested that this ewe had chronic hemolytic anemia (copper poisoning, proven by mineral element testing), with a history of exposure to T. gondii.
T. gondii parasites were also observed in the lungs of a lamb from Zhumadian, Henan (13 September 2021) by IHC, while T. gondii DNA was also detected in the lungs and diaphragm of this ewe. T. gondii parasites were not observed in paraffin sections of 132 other sheep.
3.3. Isolation of the T. gondii Strain from Sheep Tissues or Blood
We selected tissue samples and the blood of 24 sheep to isolate T. gondii by mouse bioassay (Table 1). In the Tox#35-43 group (“Tox#35” indicates samples from sheep, “43” is the group number), three Swiss mice (M#628, M#629, and M#630) were inoculated with the digestive fluid of the skeletal muscles, liver, and spleen from the ewe (sampling from 11 October 2021, Xinxiang) (Figure S1). These three mice died at 15–27 DPI with clinical manifestations of depression and poor appetite, and tachyzoites were found in the lungs of the M #629 mice (Figure 3A). In the Tox#35-44 group, five Swiss mice (M#631-M#635) were inoculated with ground tissues from the lymph nodes and spleen of the ewe (sampling from 11 October 2021, Xinxiang). These mice died of bacteremia 2–4 DPI. Tox#35-45, Tox#35-46, and Tox#35-47 were inoculated with the myocardium, lungs, spleen, and lymph nodes of Tox#35-44 mice. T. gondii parasites were observed in the lungs of M #643 (Tox#35-46) at 19 DPI (Figure 3B). Tox#35-48 and Tox#35-49 were inoculated with the tissues of Tox#35-45 and Tox#35-47 mice, respectively. The 50% mice were seropositive for T. gondii (≥1:200) when using MAT in the Tox35-48 (M #654) and Tox35-49 (M #659) groups at 30 DPI. Tachyzoites were observed in the lymph nodes and lungs of M#654 at 32 DPI. The survival time for M#659 was 116 DPI, and no brain cysts were observed.
The brain, lymph nodes, and lungs of M#654 mice were homogenized and inoculated into the mice groups and seeded into Vero cells. This T. gondii strain could propagate slowly in cell culture at 16 DPI, and bradyzoites could be formed in the cell culture medium at 50 DPI. In vivo, only 42.9% of the inoculated mice groups (9/21) could infect T. gondii (25–50% mice infected with T. gondii in positive groups, verified by MAT or crescent-shaped parasites). This strain was designated as TgSheepCHn15.
In the Tox#35-41 group (lamb samples from 23 September 2021, Zhoukou), 1/5 mice (M#789) were seropositive for T. gondii, and parasites were observed in the IFN-γ^−/−^ mice M#727 lungs at 6 DPI. Unfortunately, none of the mice were exposed to T. gondii after being sub-passaged.
No viable T. gondii strain was isolated from other sheep (n = 22).
3.4. Genotypes of TgSheepCHn15
The TgSheepCHn15 genotype was determined from the heart, lungs, and lymph nodes of M#654 (Tox#35-48), and it belonged to ToxoDB genotype #10 (Figure S2, Table S2). The ROP18/ROP5 allele type was 1/1 (Figure S2). Infection with the TgSheepCHn15 strain was in all of the inoculated mice. According to the survival time (97 ± 31) of infected mice (Table 3), the TgSheepCHn15 strain was avirulent to Swiss mice.
The genotyping of IFN-γ^−/−^ mouse M #727 (Tox35-41, lamb samples from 23 September 2021, Zhoukou) tissues using PCR-RFLP was unsuccessful. The neighbor-joining analysis based on PCR-RFLP data indicated that TgSheepCHn15 is disparate from other reported Chinese strains genotyped with PCR-RFLP 10 markers (Figure 4).
4. Discussion
A viable T. gondii TgSheepCHn15 strain was isolated from tissues of a ewe (heart juice titer = 1:25) in a veterinary clinic (Table 1). The method of grinding tissues (lymph nodes and spleen) was superior to pepsin digestion (skeletal muscles, liver, and spleen). T. gondii bradyzoites were observed in the mesenteric lymph node of the ewe (Figure 2), indicating that it might be in the acute stage of infection, supported by molecular biology evidence (T. gondii DNA was detected in the liver, lungs, and blood) and morphological evidence obtained by IHC.
The T. gondii TgSheepCHn15 genotype was ToxoDB #10 (Type I). In China, some researchers have reported the isolation and genotypes of T. gondii in sheep. Between 2019 and 2021, the T. gondii strain (ToxoDB #3) was isolated from sheep hearts [16]. From 2017 to 2019, 11 T. gondii isolates were obtained from sheep hearts. Seven isolates were ToxoDB#2 and four isolates were ToxoDB #4 [12]. From 2014 to 2016, two T. gondii strains were obtained from sheep hearts, both of which were ToxoDB #9 [20]. The genotype of the T. gondii strain in sheep from Qinghai was ToxoDB #3 [21]. T. gondii TgSheepCHn15 could grow and be sub-passaged in Vero cells and mice, but showed poor growth performance, possibly due to the characteristics of this strain. However, the exploration of the organelle structure by TEM was unsuccessful for a smaller number of T. gondii.
Toxoplasma gondii ToxoDB #10 strains and DNA seen worldwide are summarized in Table S3. Viable T. gondii ToxoDB #10 strains with high pathogenicity have been isolated from humans, goats, cats, and chickens in the USA, Brazil, Colombia, and China [1]. ToxoDB #10 nucleic acids have been found in aborted sheep fetuses from Iran [22]. Furthermore, ToxoDB #10 nucleic acids have been isolated from pigs, Microtus fortis, goats, cattle, donkeys, chickens, sparrows, foxes, raccoon dogs, and bats from China [1]. These findings were not based on viable parasites, and their virulence is unknown. However, some samples were from slaughterhouses or animals that appeared to be clinically healthy, indicating that they were avirulent for the hosts.
More than 80% of the 1/1 ROP18 and ROP5 alleles in T. gondii strains are lethal [18]. The ROP18 and ROP5 alleles of TgSheepCHn15 (1/1) were not lethal in mice (the survival time of TgSheepCHn15 infected mice was 97 ± 31 days), which is consistent with the same type of TgCtCo2 from Colombia [23]. Furthermore, no T. gondii TgSheepCHn15 cysts were found in the brain of mice (as identified by MAT and PCR). This may be related to the characteristics of the host and the inoculated parasites [24]. The genetic evolutionary distance of TgSheepCHn15 from other T. gondii strains isolated in Central China is significant, as shown in Figure 4. Overall, TgSheepCHn15 is distinctly different from the other ToxoDB #10 strains.
Ovine toxoplasmosis can cause serious economic losses worldwide [1,5,13]. Recently, the prevalence of T. gondii has been decreasing in sheep from China. However, T. gondii remains present in sheep herds and should not be ignored. The T. gondii prevalence in sheep from Henan Province from 2018 to 2024 was 7.5%, which was lower than that seen from 2017 to 2019 (25.3%) [12]. Both studies used 1:100 as a cut-off value of MAT. The low prevalence of T. gondii in sheep may be related with the development of the intensive management and health management of the sheep industry in China [25]. The seroprevalence of T. gondii in <1-year-old sheep (16.1%) was higher than that in older sheep (2.1%). This phenomenon may be related to the sample source (young sheep were mainly from veterinary clinics, and old sheep were mainly from slaughterhouses or farms) or the limited number of samples. Furthermore, the prevalence of ovine toxoplasmosis in farms or veterinary clinics was higher than that in slaughterhouses. Some unhealthy sheep enter veterinary clinics or pathology laboratories because of toxoplasmosis or other pathogens. Sheep infected with other pathogens or exhibiting sickness may also have an increased chance of infection with T. gondii. Generally, the T. gondii prevalence in sheep increased with age, indicating that ingestion oocysts constitute the main transmission route for sheep [3,5].
For chronic toxoplasmosis, the rate of isolation T. gondii or the number of tissue cysts was correlated with antibody titers in sheep and mice [11,12,26]. In this study, T. gondii DNA was found in 27 tissues or the blood of 164 sheep; only one sheep showed T. gondii antibodies (MAT titer = 1:200) from slaughterhouses, and the other 26 sheep were all <1:25 (6 sheep were from slaughterhouses, and 20 sheep were from veterinary clinics). MAT can detect IgG acting against T. gondii at the stage of chronic infection [10], indicating the need for additional examination for IgM antibodies or the etiological assessment of samples from veterinary clinics or sheep that may be in the acute stage of infection. T. gondii can be isolated from seronegative or etiological negative (low density of T. gondii parasites) animal tissues [27,28]. The findings of this study indicate that the parasite load density has no correlation with antibody titers in sheep at the stage of acute toxoplasmosis.
5. Conclusions
In the present study, one T. gondii clonal type I (ToxoDB #10) strain was isolated from adult sheep tissues. To the best of our knowledge, this is the first viable T. gondii ToxoDB #10 strain reported in sheep worldwide, and it is not lethal to mice. Further research is needed for the impact of TgSheepCHn15 and other isolated strains on sheep health and potential zoonotic risks.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Dubey J.P. Toxoplasmosis of Animals and Humans 3rd ed.CRC Press Boca Raton, FL, USA Taylor & Francis Group Abingdon, UK 20221542
- 2Klauck V. Pazinato R. Radavelli W.M. Custódio E. Bianchi A.E. Camillo G. Cezar A.S. Vogel F.F. Tonin A.A. Ferreira R. Toxoplasma gondii infection in dairy ewes: Vertical transmission and influence on milk production and reproductive performance Microb. Pathog.2016991011052753111810.1016/j.micpath.2016.08.012 · doi ↗ · pubmed ↗
- 3Innes E.A. Bartley P.M. Buxton D. Katzer F. Ovine toxoplasmosis Parasitology 2009136188718941999546810.1017/S 0031182009991636 · doi ↗ · pubmed ↗
- 4Hide G. Role of vertical transmission of Toxoplasma gondii in prevalence of infection Expert. Rev. Anti. Infect. Ther.2016143353442680749810.1586/14787210.2016.1146131 · doi ↗ · pubmed ↗
- 5Dubey J. Murata F. Cerqueira-Cézar C. Kwok O. Su C. Economic and public health importance of Toxoplasma gondii infections in sheep: 2009–2020 Vet. Parasitol.20202861091953297968210.1016/j.vetpar.2020.109195 · doi ↗ · pubmed ↗
- 6Dubey J.P. Mouse pathogenicity of Toxoplasma gondii isolated from a goat Am. J. Vet. Res.1980414274297369619 · pubmed ↗
- 7Sabin A.B. Toxoplasmic encephalitis in children J. Am. Med. Assoc.1941116801807
- 8Berger-Schoch A.E. Bernet D. Doherr M.G. Gottstein B. Frey C.F. Toxoplasma gondii in Switzerland: A serosurvey based on meat juice analysis of slaughtered pigs, wild boar, sheep and cattle Zoonoses Public Health 2011584724782182434810.1111/j.1863-2378.2011.01395.x · doi ↗ · pubmed ↗