Comparative Analysis of Metopograpsus quadridentatus (Crustacea: Decapoda: Grapsidae) Mitochondrial Genome Reveals Gene Rearrangement and Phylogeny
Dan-Dan Bian, Sheng Tang, Song-Nan Wang, Qiu-Ning Liu, Bo-Ping Tang

TL;DR
This study uses the mitochondrial genome of a marine crab to clarify its evolutionary relationships and gene arrangement patterns.
Contribution
The study reveals a conserved gene order with a unique trnH translocation in Metopograpsus quadridentatus, aiding taxonomic classification.
Findings
The mitochondrial gene arrangement of M. quadridentatus is mostly unchanged, similar to ancient crustaceans.
Phylogenetic analysis confirms M. quadridentatus belongs to the genus Metopograpsus and family Grapsidae.
Gene order conservation is observed in several Brachyura families, including Grapsidae.
Abstract
The marine crab Metopograpsus quadridentatus is categorized in the family Grapsidae, but its classification remains unclear because of limited differences in appearance among species. In this research, we examined its mitochondrial genome, which showed a gene order similar to that of ancient crustaceans, with the exception of the translocated trnH gene. Analysis of 13 protein-coding genes from 40 Brachyura species indicated that it belongs to the genus Metopograpsus and family Grapsidae, clarifying its evolutionary relationships and demonstrating the importance of mitochondrial genomes in taxonomic classification. The taxonomy of the genus Metopograpsus is still a matter of debate because its species have limited morphological differences. Mitochondrial genomes, which are highly informative and easily accessible genetic markers, have been widely used to study molecular evolution and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
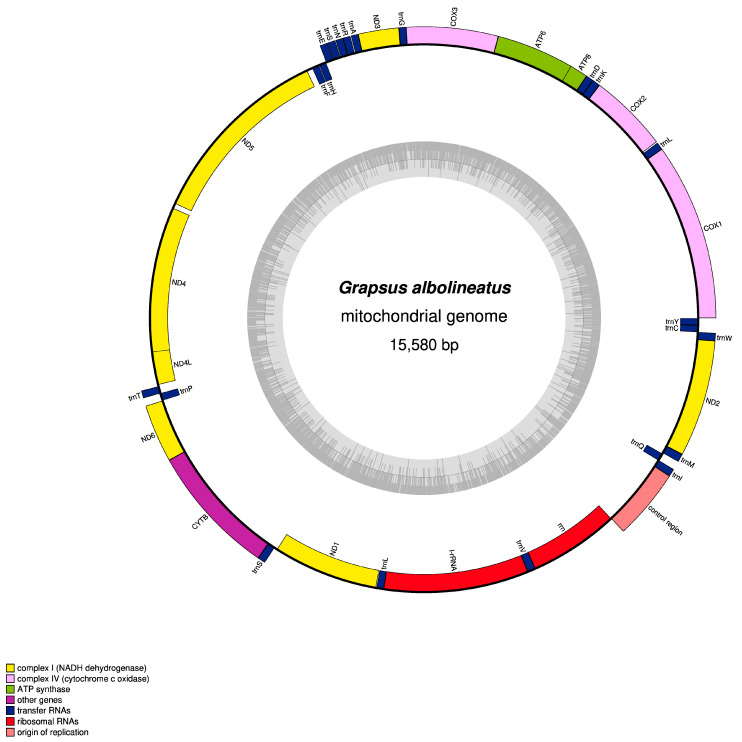 Figure 1
Figure 1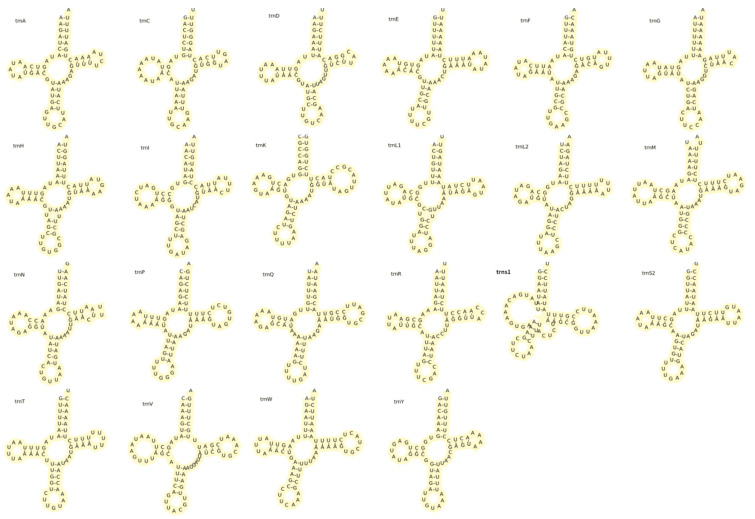 Figure 2
Figure 2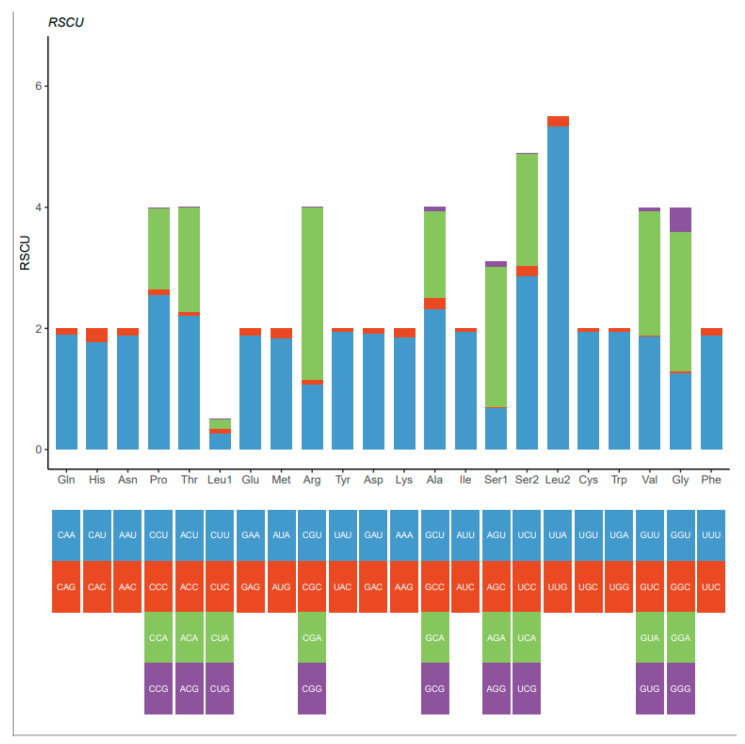 Figure 3
Figure 3 Figure 4
Figure 4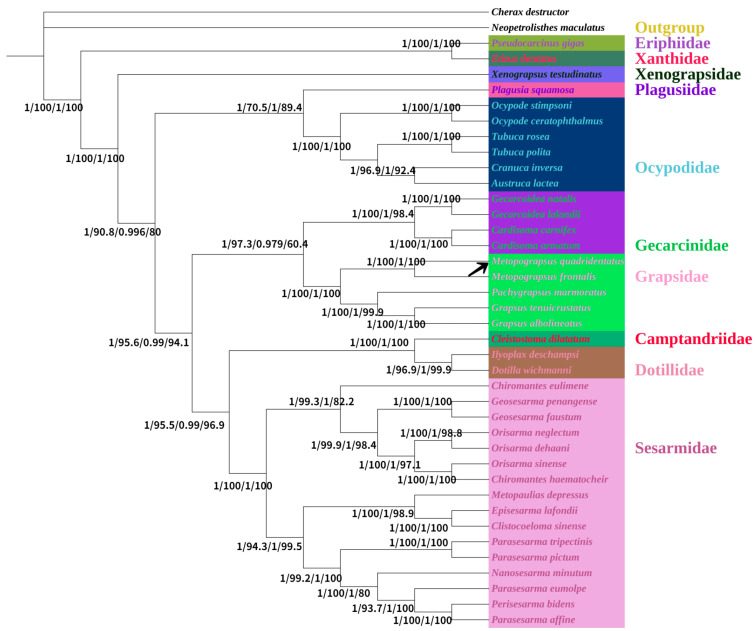 Figure 5
Figure 5- —National Natural Science Foundation of China
- —Jiangsu Province
- —YCTU
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCrustacean biology and ecology · Genomics and Phylogenetic Studies · Genetic diversity and population structure
1. Introduction
Mitochondria are organelles found in eukaryotic cells that house their own genetic material, known as the mitochondrial genome, characterized by a high mutation rate, lack of gene recombination, a high replication rate, and maternal inheritance [1,2]. In metazoans, the mitochondrial genome is a small double-stranded circular molecule typically 16–20 kbp in size [3]. It contains 13 protein-coding genes, including subunits of NADH dehydrogenase, cytochrome b, cytochrome c, and ATP synthase, along with transfer RNA genes, ribosomal RNA genes, and a non-coding control region that regulates the mitochondria genome’s transcription and replication. Mitochondrial DNA (mtDNA) in animals exhibits remarkable organizational precision [4]. It contains 13 protein-coding genes: ND1-6 and ND4L encode core catalytic subunits of Complex I, Cyt b acts as the electron transfer hub of Complex III, COI-III constitute the oxidative center of Complex IV, and ATP6/8 drive proton gradient conversion in Complex V. This compact arrangement, interspersed with 22 tRNA genes, reflects evolutionary optimization for genomic compression and expression regulation. Notably, newly identified regulatory elements within the D-loop control region—such as termination-associated sequences (TASs) and conserved sequence blocks (CSBs)—participate in replication initiation through secondary structure formation, with their polymorphic features now serving as vital molecular markers in population genetics [5].
The infraorder Brachyura consists of 98 families and nearly 7000 described species [6]. Metopograpsus quadridentatus is a type of marine crab commonly found in rock crevices or under rocks at low tide. It is found in various locations, including Malacca, Java, Bali, Kalimantan, New Guinea, the Indian Ocean, Guangdong, Fujian, Zhejiang, Shandong, and other areas in China [7]. Currently, Metopograpsus quadridentatus is classified under Crustacea, Decapoda, Brachyura, Grapsidae, genus Metopograpsus. Due to advancements in sequencing technology, especially next-generation sequencing, the time and cost required for sequencing have substantially decreased, making it simpler to acquire the mitochondrial genome sequence [8]. Mitochondrial gene-based molecular phylogenetic analyses first confirmed the monophyly of all six Metopograpsus crab species, revealing high intraspecific genetic variation and a pronounced phylogeographic structure in M. thukuhar and M. quadridentatus, suggesting the presence of undescribed evolutionarily significant units (ESUs) that require further taxonomic validation via nuclear DNA and morphological investigations [9]. Mitochondrial genome analysis is widely used in phylogenetic analysis, biogeography, population genetics, medicine, and ecology [10,11,12,13,14]. The mitochondrial genome of Helice wuana and its relatives offer crucial evidence on the origin, germline evolution, and unique genetic structure of crab [15]. The mitochondrial genome contains 37 genes, which theoretically have a high rearrangement potential. However, according to existing results, gene rearrangements are not very common. Similar gene rearrangements among different species are thought to be unlikely due to convergent evolution but instead suggest a certain phylogenetic relationship among species [16]. As a result, analyzing gene rearrangement is more appropriate for determining the superior phylogenetic relationship [17].
In this research, we analyzed the complete mitochondrial genome of M. quadridentatus and compared it with other brachyura species. By examining the amino acid and nucleotide sequences of 13 protein-coding genes, we built phylogenetic trees. Additionally, we studied the gene rearrangement patterns to determine the evolutionary placement of M. quadridentatus within the brachyura.
2. Materials and Methods
2.1. Sampling and DNA Extraction
Mature crabs (M. quadridentatus) were gathered from Xiamen, Fujian Province, China. Prior to DNA extraction, the specimen was kept in aerated tap water at a temperature of 21 ± 1 °C for a period of 1 week. DNA was extracted from muscle tissue samples using an Aidlab Genomic DNA Extraction Kit (Aidlab, Beijing, China), and the extracted DNA was then preserved at −20 °C until amplification.
2.2. Sequencing and Analysis
Prior to mitochondrial genome amplification of M. quadridentatus, commercially prepared universal primers were obtained from Sunbiotec (Beijing, China) [18]. Genomic DNA amplification was conducted following standardized protocols provided with the Aidlab Extraction Kit (Beijing, China). Polymerase chain reactions were executed in 50 μL volumes comprising 5 μL of 10× Taq Plus Buffer (Mg^2+^ supplemented), 4 μL dNTP mixture, 2 μL forward/reverse primers (each), 2 μL template DNA, 34.5 μL molecular-grade ddH_2_O, and 0.5 μL Taq DNA polymerase RED. Thermal cycling parameters included an initial denaturation period at 94 °C for 3 min, followed by 35 cycles of denaturation at 90 °C for 30 s, primer-specific annealing (49–58 °C for 35 s, with temperature optimized per primer pair), and extension at 72 °C (30 s to 4 min duration, scaled by target fragment size), concluding with terminal elongation at 72 °C for 10 min. Electrophoretic separation was conducted using 1% agarose gels (w/v) for PCR product visualization, followed by purification with Aidlab’s DNA Gel Extraction System (Beijing, China). The processed amplicons were subsequently cloned into T-vector systems for sequencing analysis (Sangon Biotech, Shanghai, China).
2.3. Bioinformatics Analysis
Mitochondrial genome annotation of M. quadridentatus was performed via NCBI BLAST homology searches (https://blast.ncbi.nlm.nih.gov/Blast.cgi). Sequence assembly and refinement were conducted using DNASTAR’s SeqMan module version 7 (DNAStar Inc., Madison, WI, USA) [19]. Genome architecture was visualized through OrganellarGenomeDRAW (OGDRAW) to generate a graphical genome map [20]. Transfer RNA identification and secondary structure prediction (cloverleaf conformation) were achieved using the tRNAscan-SE algorithm [21]. Nucleotide composition, synonymous codon usage bias, and compositional skewness (AT skew = [A − T]/[A + T]; GC skew = [G − C]/[G + C]) were quantified using MEGA 12 [22,23].
2.4. Phylogenetic Analysis
Mitogenomic sequences from Brachyura species and two outgroup taxa (Cherax destructor and Neopetrolisthes maculatus) were retrieved from NCBI GenBank (accessions in Table 1). Protein-coding genes (13 PCGs) were aligned using MAFFT v7 under the invertebrate mitochondrial genetic code [24,25], with ambiguously aligned regions trimmed via Gblocks [26]. Consolidated alignments were generated using PhyloSuite’s sequence concatenation function [27,28]. Evolutionary model selection was optimized with PartitionFinder2 [29], followed by partitioned phylogenetic analyses using MrBayes v3.2 and IQ-TREE [30,31]. The GTR + I + G and MtRev + I + G + F models were chosen as the best models for nucleotide and amino acid phylogenetic analysis. The MrBayes software v3.2 underwent ten million generations with four chains, sampling every 100 generations, with a burn-in step of 5000 generations. The average standard deviation of the split frequency was less than 0.01, indicating convergence. The IQ-TREE software version 2 was run with 1000 bootstrapped replicates, and results were analyzed using Tracer v1.6. The effective sample size (ESS) value was over 200, showing convergence of the chains in the Markov chain Monte Carlo. Phylogenetic trees were visualized using the online tool Interactive Tree Of Life [32].
3. Results and Discussion
3.1. Mitogenome Organization and Nucleotide Composition
The mitochondrial genome of M. quadridentatus exhibits a covalently closed circular configuration with double-stranded topology, spanning 15,523 base pairs (Figure 1). Consistent with typical Brachyuran mitochondrial architecture, it harbors 37 canonical genes: 13 protein-coding genes (PCGs), 22 transfer RNA (tRNA) genes, 2 ribosomal RNA (rRNA) genes, and a non-coding AT-rich regulatory region (Figure 2). Gene strand distribution reveals 14 tRNA genes (trnL2, trnK, trnD, trnG, trnA, trnR, trnN, trnS1, trnE, trnT, trnS2, trnI, trnM, and trnW) and all PCGs encoded on the minority strand, while the remaining genes (8 tRNAs, 2 rRNAs, and the AT-rich region) reside on the majority strand (Table 2). Genomic nucleotide composition demonstrates marked AT bias, with adenine (34.4%) and thymine (36.3%) collectively constituting 70.7% of the total bases, contrasted by cytosine (19.6%) and guanine (10.3%). The AT-rich region accounts for 70.4% of its sequence. Calculated compositional skews (AT skew = [A − T]/[A + T] = −0.025; GC skew = [G − C]/[G + C] = −0.314) indicate moderate adenine depletion and pronounced guanine deficiency. Comparative analysis across Brachyura reveals that M. quadridentatus shares the negative GC skew trend, while only 10 congeners exhibit marginally positive AT skew values (Table 3).
3.2. Protein-Coding Genes
As shown in Table 2, the 13 protein-coding genes (PCGs) varied in length, ranging from 159 bp (atp8) to 1734 bp (nad5). The PCG region of the mitochondrial genome of M. quadridentatus was 11,172 bp in length and consisted of 13 genes (nad1–6, nad4L, cox1–3, atp6, atp8, and cytb). The start codon for 12 of the 13 protein-coding genes is ATG (T/G), while the start codon for the nad4 gene is CTG. The mitochondrial genome of M. quadridentatus exhibits standard termination codons (TAA or serine-encoding TCG) in ten protein-coding genes (PCGs), whereas cox1, cox2, and cytb feature incomplete T-nucleotide stop signals (Table 2). Codon utilization patterns across PCGs are summarized in Table 3, revealing a total of 3724 codons. Leucine (Leu, 15.9%) predominates as the most abundant amino acid, followed by serine (Ser, 9.80%) and phenylalanine (Phe, 8.83%). The highest-frequency codons include UUA (Leu), UUU (Phe), and AUU (isoleucine) (Table 4). Relative synonymous codon usage (RSCU) profiles for M. quadridentatus (Figure 3) corroborate these findings, validating codon bias trends observed in M. quadridentatus.
3.3. Transfer RNA and Ribosomal RNA Genes and Control Region
The mitochondrial genome of M. quadridentatus includes two ribosomal RNA genes, rrnL (1323 bp) and rrnS (832 bp), with a trnV gene positioned between them—a conserved feature among metazoans [55]. The genome encodes 22 tRNA genes (Table 2), ranging from 63 to 74 nucleotides (total combined length: 1475 bp). Eight tRNAs (trnH, trnF, trnP, trnL1, trnV, trnQ, trnC, and trnY) were encoded on the minority strand. tRNAscan-SE analysis predicted canonical cloverleaf secondary structures for all tRNAs (Figure 2), except trnS1, which exhibited an elongated dihydroxyuridine (DHU) arm and a distinctive auxiliary loop, a structural anomaly consistent with Brachyuran mitogenomes [56]. Genomic organization analysis identified 18 nucleotide overlaps (1–32 bp) and five intergenic spacers (2–7 bp), excluding the control region. A 568 bp control region, positioned between rrnS and trnI, harbors conserved sequences associated with replication origin and transcriptional regulation (Table 2).
3.4. Gene Arrangement
Mitochondrial gene rearrangement is a valuable tool for studying the evolutionary relationships of organisms at higher taxonomic levels [57]. In recent years, this technique has been used to elucidate the evolutionary history of various groups such as birds, marsupials, echinoderms, nematodes, and others [58]. When comparing the gene order of the entire mitochondrial genome of M. quadridentatus to the Pancrustacean ground pattern, we observed that, except for the translocation of trnH, the gene sequences were identical (Figure 4). The mitochondrial genome of Eucrate crenata exhibits a notable rearrangement involving the translocation of trnH-cac, which deviates from the conserved gene order typically observed in decapods. In most brachyuran crabs, trnH-cac is positioned between nad5 and nad4, a pattern considered ancestral for the group. However, in E. crenata, this tRNA gene is relocated between trnE-gaa and trnF-ttc, marking a significant structural divergence [59]. Such rearrangements, though uncommon in Brachyura, are increasingly recognized as phylogenetically informative markers [19]. The trnH translocation observed here may reflect lineage-specific evolutionary dynamics, potentially arising from mechanisms such as tandem duplication–random loss (TDRL) or recombination-mediated processes [42]. Additionally, when compared to the gene rearrangement of other brachyura species in our study, we found that the gene order was the same among families Eriphiidae, Grapsidae, Camptandriidae, Dotillidae, Plagusiidae, Ocypodidae, and Gecarcinidae. However, in Sesamidae, gene rearrangement was observed in the transposition of trnQ and trnI. The mechanism of mitochondrial genome rearrangement is believed to involve errors in light-strand replication and gene tandem duplication, which can occur due to replication errors such as incorrect start or stop signals and slipped mispairing [60]. The gene order of Varunidae, Macrophthalmidae, and one Pinnotheridae species, Tritodynamia horvathi, are the same. The gene rearrangement pattern of Homolidae, Raninidae, Orithyiidae, Oregoniidae, Majidae, Bythograeidae, Oziidae, Xanthidae, Leucosiidae, Matutidae, Eriphiidae, Menippidae, Carpiliidae, Ovalipidae, Geryonidae, Portunidae, and some Potamidae species are the same or similar [19]. This rearrangement gains further significance when contextualized within the phylogenetic framework [43]. While gene order conservation is often emphasized in mitogenomic studies, the trnH translocation in Brachyura underscores the utility of structural variations in resolving deep phylogenetic nodes, particularly within Decapoda, where traditional morphological classification faces challenges [25]. Our study also revealed frequent gene rearrangements in tRNAs, protein-coding genes, and rRNAs in Xenograpsus testudinatus and Etisus dentus, suggesting the occurrence of mitochondrial gene recombination [54].
3.5. Phylogenetic Analysis
Phylogenetic reconstruction was initiated by identifying taxonomically proximate Brachyura species through NCBI BLAST homology screening. Nucleotide and amino acid sequences of 13 protein-coding genes (PCGs) from Brachyuran taxa (Table 1) were employed to resolve the phylogenetic position of M. quadridentatus. Topological congruence between the BI and ML trees permitted integration into a consensus phylogeny (Figure 5). Both analyses robustly placed M. quadridentatus and M. frontalis within a monophyletic clade, supported by statistically robust nodal values (posterior probability ≥ 0.95; bootstrap ≥ 90%), strongly suggesting a sister-group relationship between these congeners. Both M. quadridentatus and M. frontalis were clustered in the same branch with Grapsidae species. We can infer from this that M. quadridentatus belongs to genus Metopograpsus, family Grapsidae, which is consistent with the findings of a previous study [19]. The phylogenetic trees based on amino acid and nucleotide sequences have different topologies. In the phylogenetic trees based on nucleotide sequences, the topology is ((((((Sesermidae + Gecarcinidae) + Xenograpsidae) + (Ocypodidae + Plagusiidae)) + (Camptandriidae + Dotillidae)) + Grapsidae) + (Eriphiidae + Xanthidae)). The topology of the phylogenetic trees based on amino acid sequences is (((((Sesermidae + (Camptandriidae + Dotillidae)) + (Grapsidae + Gecarcinidae)) + (Ocypodidae + Plagusiidae)) + Xenograpsidae) + (Eriphiidae + Xanthidae)). This robust support for their placement within Grapsidae aligns with broader efforts to clarify brachyuran systematics, similarly resolving Grapsidae as a cohesive lineage using mitochondrial PCGs [40]. However, the conflicting higher-level topologies between nucleotide- and amino acid-based trees—such as Sesarmidae clustering with Gecarcinidae/Xenograpsidae in nucleotide trees versus Camptandriidae/Dotillidae in amino acid trees—underscore persistent challenges in reconstructing deep brachyuran relationships. These discrepancies echo Shen et al., who noted that mitochondrial sequence data, while valuable, often struggle to resolve rapid radiations or ancient divergences due to homoplasy and saturation, particularly in nucleotide alignments [52]. The polyphyly of traditional superfamilies like Grapsoidea and Ocypodoidea in our study further reflects the unresolved nature of brachyuran classification. For instance, the close association of Potamoidea (Parathelphusidae + Potamidae) with Thoracotremata, rather than their marine relatives in Heterotremata, parallels ecological transitions observed in freshwater crabs like Potamon [19]. Such anomalies suggest that adaptive radiations in novel niches (e.g., freshwater habitats) may obscure phylogenetic signals, as seen in Potamon’s origin in Western Asia and subsequent diversification across the Mediterranean [61]. This ecological divergence complicates molecular phylogenetics, as convergent adaptations can mimic shared ancestry, necessitating integrative approaches that combine mitochondrial data with nuclear loci or morphological traits [62]. The incongruent placements of Sesarmidae and Gecarcinidae in our trees also align with broader debates in brachyuran systematics. Zhang et al. similarly reported unstable relationships among Sesarmidae, Gecarcinidae, and Xanthidae in mitochondrial phylogenies, emphasizing the need for expanded taxon sampling [35]. Shen et al. further demonstrated that mitochondrial genomes alone may fail to resolve deep nodes without comprehensive representation of major lineages, a limitation exacerbated by the uneven sampling of families like Camptandriidae and Xenograpsidae in our study [52]. The reason for such discrepancies is that the classification of Brachyura crabs was not perfect, and more phylogenetic studies of Brachyura crabs are needed in the future. Owing to the ongoing development of DNA sequencing technology and bioinformatics, molecular technology can now be used to classify phylogeny [1,61]. The computing power of computers is increasing, which can be used to process molecular data. Using bioinformatic technology to analyze the mitogenome in the future will aid in evolutionary biology studies to classify crabs.
4. Conclusions
In this study, we analyzed the whole mitogenome of M. quadridentatus. Using gene arrangement patterns and phylogenetic analysis, we suggested that M. quadridentatus belongs to the Grapsidae family. While mitochondrial sequences remain a cornerstone of brachyuran phylogenetics, their limitations in resolving deep nodes call for methodological innovation. By combining mitochondrial data with nuclear markers, time-calibrated models, and robust taxon sampling, future studies can unravel the evolutionary history of Brachyura, particularly for ecologically divergent lineages like Grapsidae and Potamoidea.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Tsang L.M. Schubart C.D. Ahyong S.T. Lai J.C.Y. Au E.Y.C. Chan T.Y. Ng P.K.L. Chu K.H. Evolutionary History of True Crabs (Crustacea: Decapoda: Brachyura) and the Origin of Freshwater Crabs Mol. Biol. Evol.2014311173118710.1093/molbev/msu 06824520090 · doi ↗ · pubmed ↗
- 2Ma H. Ma C. Li X. Xu Z. Feng N. Ma L. The complete mitochondrial genome sequence and gene organization of the mud crab (Scylla paramamosain) with phylogenetic consideration Gene 201351912012710.1016/j.gene.2013.01.02823384716 · doi ↗ · pubmed ↗
- 3Rackham O. Filipovska A. Organization and Expression of the Mammalian Mitochondrial Genome Nat. Rev. Genet.20222360662310.1038/s 41576-022-00480-x 35459860 · doi ↗ · pubmed ↗
- 4Zhang Y. Wei L. Liu B. Liu L. LüZ. Gong L. Two complete mitogenomes of Ocypodoidea (Decapoda: Brachyura), Cleistostoma dilatatum (Camptandriidae) and Euplax sp. (Macrophthalmidae) and its phylogenetic implications Acta Oceanol. Sin.202342819210.1007/s 13131-022-2054-9 · doi ↗
- 5Zhang Y. Gao Y. Gong L. Lu X. Jiang L. Liu B. Liu L. LüZ. Li P. Mitochondrial genome of Episesarma lafondii (Brachyura: Sesarmidae) and comparison with other sesarmid crabs J. Ocean Univ. China 2021201545155610.1007/s 11802-021-4779-z · doi ↗
- 6Guinot D. Jamieson B.G.M. Deforges B.R. Relationship of Homolidae and Dromiidae: Evidence from Spermatozoal Ultrastructure (Crustacea, Decapoda)Acta Zool.19947525526710.1111/j.1463-6395.1994.tb 01213.x · doi ↗
- 7Shen J. Dai A. Illustrated Fauna of China: Crustacea Science Press Beijing, China 1964 Volume 21142
- 8Metzker M.L. Sequencing Technologies—The Next Generation Nat. Rev. Genet.201011314610.1038/nrg 262619997069 · doi ↗ · pubmed ↗