Ontogenesis from Embryo to Juvenile in Threadsail Filefish, Stephanolepis cirrhifer
Liming Liu, Xuanhan Liu, Yanqing Wu, Jun Zeng, Wengang Xu

TL;DR
This study describes the early development stages of the threadsail filefish to help develop methods for its artificial breeding.
Contribution
The study provides detailed morphological data on ontogenesis from embryo to juvenile in a threatened marine species.
Findings
Embryonic development is divided into eight distinct phases from cleavage to hatching.
Two inflection points in growth curves correlate with metamorphosis and feeding habit changes.
Morphological markers like fin differentiation and pigmentation were identified at specific developmental stages.
Abstract
This study details and evaluates the morphological characteristics of the early embryonic development and ontogenesis of Stephanolepis cirrhifer, commonly known as the threadsail filefish. This species has experienced a considerable reduction in numbers in recent decades to the point of being listed as a threatened species in 2017, yet techniques for developing artificial propagation of the species have remained elusive. Our findings probably define the morphological characteristics of ontogenesis from embryo to juvenile. The embryonic development was observed and divided into eight phases, which were cleavage, blastocyst, gastrula, neurula, organogenesis, muscular contraction, heart pulsation, and hatching. Additionally, we found that the weaning period occurred from 40 dph, and two inflexion points occurring in the growth curves of larvae and juveniles were found to be associated with…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
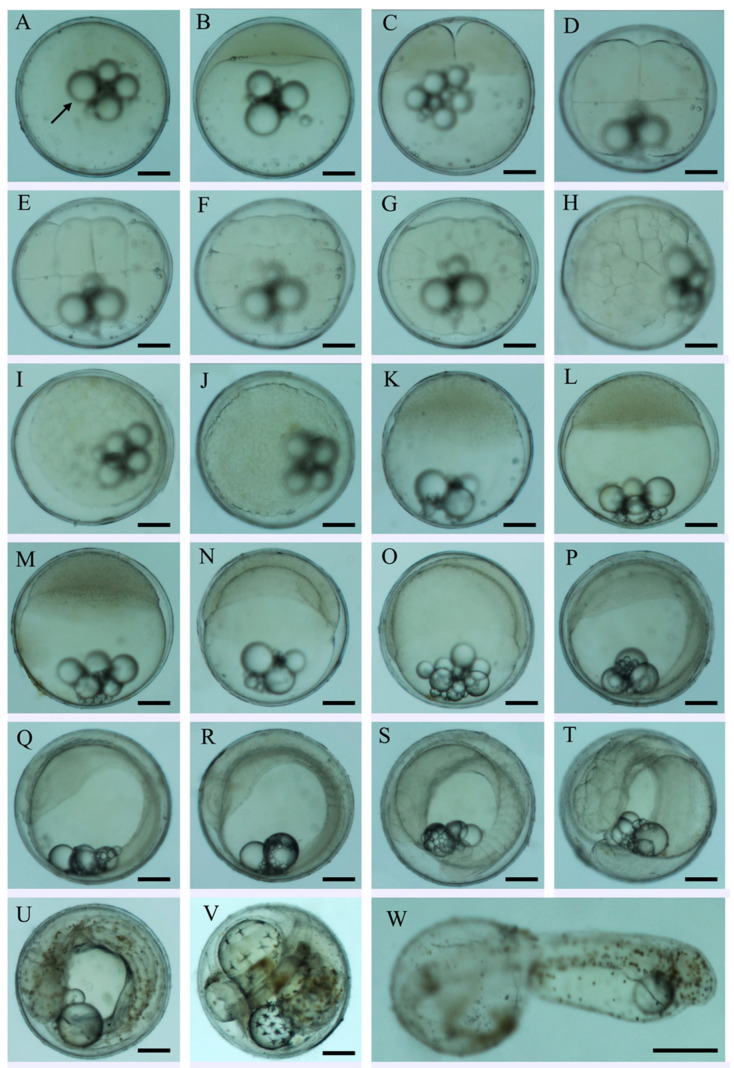 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3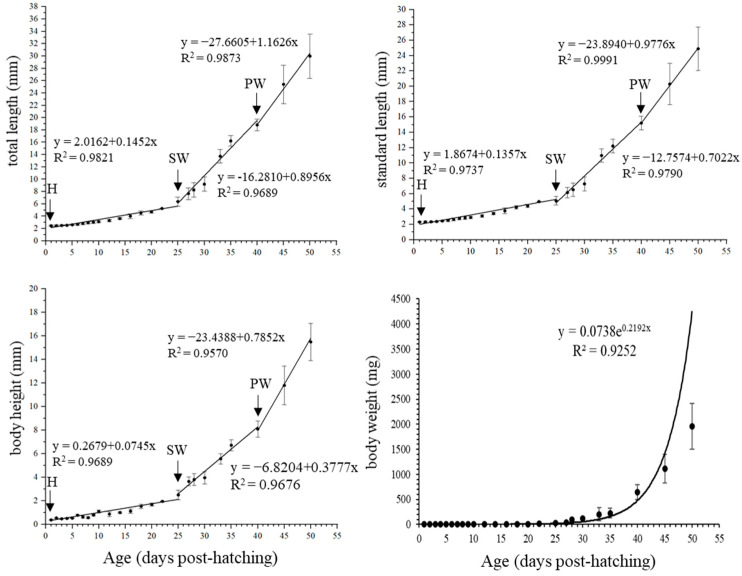 Figure 4
Figure 4- —Guangxi Academy of Sciences Special Project for Reform and Development
- —Fangchenggang Science and Technology Innovation Development Plan
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsReproductive biology and impacts on aquatic species · Aquaculture Nutrition and Growth · Aquatic life and conservation
1. Introduction
Filefish belong to the Monacanthidae family, comprising approximately 102 species and 32 genera distributed worldwide, with most species inhabiting coastal reefs, coral reefs, and sand sea areas [1]. In particular, the threadsail filefish, Stephanolepis cirrhifer (Temminck and Schlegel, 1850), is mainly distributed in the Western Pacific waters from Hokkaido to the East China Sea [2,3,4] and is an important commercial species in Asian countries, especially in Japan and South Korea [5,6,7,8]. In Japan, the liver and sashimi of S. cirrhifer are commonly used as premium ingredients [9], whereas in South Korea, dried S. cirrhifer fish products are popular [10]. Furthermore, S. cirrhifer prey on jellyfish larvae, with the potential to inhibit jellyfish booms [4,11].
Owing to overfishing, habitat destruction, and marine environmental pollution, wild catches of S. cirrhifer have sharply decreased from a collective output in excess of 250,000 tons in the Northwest Pacific in 1985 to below 350 tons in 2002 [12]. Similarly, in 2009, the annual S. cirrhifer catch dropped to 1500 tons, representing a 99% decrease from 1990 to 2009 in South Korea [13] and a corresponding 99% decrease in wild catch output. Consequently, in 2017, S. cirrhifer was placed on the IUCN Red List of Threatened Species [14].
Over the past decade, studies on S. cirrhifer have focused on its external morphology [15,16], genetic characteristics, population genetic structure [17,18], acoustic target strength [19], and feed nutrient levels [20,21]. This has led to the development of artificial culture and breeding techniques to increase production and maintain resource balance [22,23]. For example, one study reported that artificially cultured S. cirrhifer grew rapidly, reaching the commercial standard within 1 year, with a higher-quality liver than wild species [8]. In addition, S. cirrhifer exhibit cage-cleaning properties by feeding on attached algae and small animals. Hence, improved S. cirrhifer farming has important implications for human consumption and cage-cleaning processes. However, given that the artificial breeding technology has not yet achieved complete success, the main source of fish larvae depends on wild catches [18], and large-scale cultivation of S. cirrhifer remains to be realized.
Our previous study considered the changes in the growth and feeding characteristics during early ontogenesis in S. cirrhifer [24]. To improve the survival rate of larvae, the optimization of the processes of early embryonic development and ontogenesis after hatching is critical to fish culture. To date, few studies have provided a detailed characterization of these life stages for S. cirrhifer. Meanwhile, related processes have been thoroughly investigated in rockfish [25] and flatfish [26], both of which have been successfully cultivated on a large scale. Therefore, studies on the early development of fish species are needed to understand their basic biology, feeding requirements, and environmental preferences to develop successful artificial breeding strategies [27,28]. Accordingly, herein, we describe the morphological development and growth of S. cirrhifer throughout the early life stages.
2. Materials and Methods
2.1. Acquisition of Fertilized Eggs
The present study was conducted at Yantai Laizhou Aquatic Co., Ltd. in Shandong, China, in 2020. One hundred female and one hundred male wild parental S. cirrhifer caught in the Yellow Sea were selected for indoor ripening cultivation in November 2019. The average body weights of female and male fish were 520.17 ± 56.30 g and 401.15 ± 46.19 g, respectively. The fish were fed with fresh mussel Mytilus edulis and oyster Ostreidae daily, together with live sandworms Nereis succinea to promote gonadal maturation. Male and female fish were naturally mated and spawned after 3 months in February 2020. Polyethylene sheets were placed at the bottom of the pond and used as attachment bases for the collection of the fertilized eggs. Once several eggs were attached to the sheets, they were immediately removed and placed in an incubator.
2.2. Egg Incubation
The fertilized eggs were incubated in a 20 m^3^ concrete pool with gentle aeration and circulating seawater. The seawater was purified through two sand filters (DS-1200, Yixing Bohui Environmental Protection Technology Co., Ltd., Yixing, China) connected in series, and the salinity was 33 ± 0.5 psu. The water temperature and pH were maintained at 23.6 ± 0.5 °C and 7.8–8.2, respectively. The light intensity at the water surface was maintained at 5000–8000 lx using fluorescent lamps (E27, Opple Co., Ltd., Zhongshan, China). Furthermore, a flow rate of 40–50% seawater exchange/day was applied in this study.
2.3. Culture of Larvae and Juveniles
From hatching to 3 days post-hatching (dph), the larvae mostly rely on their own yolk sac as nutrition for survival. From 3 dph, they started to open their mouths for feeding. Based on the larval size, they were fed the rotifer (Brachionus plicatilis) at a density of 1–2 ind/mL in water from 3 to 25 dph, Artemia salina nauplii at a density of 0.5–1 ind/mL from 25 to 40 dph, and formula feed after 40 dph. During the changes in the feeding regimes, the proportion of two types of food was half each.
The cultivation condition was shaded, and 20 W LED lamps were used to adjust light intensity. A portable water quality detector (Hengxin, 86031, AZ Instrument Corp., Taiwan, China) was used daily to measure water temperature, salinity, dissolved oxygen, and pH. The water quality parameters were as follows: temperature, 24.1–25.2 °C; salinity, 31.4–32.2 psu; pH, 8.00–8.27; dissolved oxygen levels, 5.0–5.9 mg/L. The aeration was maintained for 24 h, and seawater exchange rate was gradually increased from 10% to 150%/d at the settling stage of juveniles.
2.4. Observation of Embryonic Development and Larval/Juvenile Morphology
At each developmental stage of embryos and larvae, 20 specimens were sampled daily from the incubation and larval rearing cement pool, respectively. The morphological characteristics and duration of development stages of larvae and juveniles were observed and photographed using a light microscope (Nikon SMZ800, Nikon, Tokyo, Japan). A digital CCD camera (Nikon US300) and VImage 2014 software (Vezu Technology Co., Ltd., Guangzhou, China) was used to capture images and measure the diameters of the oil globules and yolk sacs. The growth indices of each sampled larva, such as total length (TL), standard length (SL), body height (BH), were measured. The body weight (BW) of larva was measured using an electronic balance (ME55; Mettler-Toledo International Inc., Zurich, Switzerland) with a sensitivity of 0.01 mg.
2.5. Image Processing and Statistical Analysis
The larvae were placed in a culture dish, and photographs of the developmental stages were implemented using Photoshop CS6 (Adobe Systems, San Jose, CA, USA). All data were analyzed using SPSS v26.0 software (IBM, Armonk, NY, USA). Data are expressed as the mean ± standard deviation (SD). The TL, SL, BH, and BW were plotted against age. Linear and nonlinear regressions were performed using OriginPro 9.1 (OriginLab Corp., Northampton, MA, USA). The inflexion points were calculated according to Van Snik et al. [29].
3. Results
3.1. Embryonic Development
The total 120,000 fertilized eggs of S. cirrhifer were spherical, sticky, and demersal, and the diameter of eggs was 0.62 ± 0.01 mm (Figure 1A). The yolk was transparent and contained multiple oil globules of varying sizes. The ooplasm was evenly distributed, and the membrane was thick, comprising 4–6 oil globules (100–142 μm) and 8–12 smaller oil globules (<20 μm). Unfortunately, we failed to count the survival rate of embryos at each developmental stage from fertilized egg to hatching in this study.
At 23.6 °C, it took about 48 h for the eggs to develop into larvae. The embryonic characteristics and duration of development in each stage of S. cirrhifer are shown in Figure 1 and Table 1, referred to from our recent study focusing on the species of Senegalese sole Solea senegalensis [30]. The hatching rate was about 70–85%.
3.2. Morphological Characteristics of Larvae and Juveniles
The morphological characteristics of S. cirrhifer post-embryo from 0 to 50 dph are shown in Figure 2, and its characteristics are described in detail in Table 2. The yolk sac of newly hatched larvae was visible in the abdomen, with an average long diameter of 0.53 ± 0.03 mm and short diameter of 0.28 ± 0.03 mm. An oil globule was observed in front of the yolk sac, with an average diameter of 0.18 ± 0.01 mm. The mouth and anus were closed; the larvae were unable to swim and were suspended upside down in the water, constituting the pre-larval phase (Figure 2a–d). At 4 dph, the yolk sac was completely absorbed, and the oil globules were almost invisible. The oral fissure formed, and the digestive tract was connected, constituting the post-larval phase. Subsequently, fins appeared and gradually differentiated, gradually improving the movement and predation abilities (Figure 2e–p). After 22 dph, in the juvenile phase, fish were evenly distributed in the water and successfully swam against the current of water. Juveniles were gradually fed compound feed (Figure 2q–v). During this phase, the juveniles completed metamorphosis, with body and fin morphologies similar to those of adult fish (Figure 2w). At 50 dph, young fish exhibited strong swimming and avoidance, with an ~80% individual survival rate. Their morphological structures and ecological habits were the same as those of adult fish (Figure 2x).
3.3. Growth Pattern of Larvae and Juvenile
From 0 to 50 dph, the TL, SL, BH, and BW on each day are shown in Table S1 (Supplementary Material). From 0 to 50 dph, the TL increased from 2.45 ± 0.06 to 29.97 ± 3.61 mm, and BW increased from 0.15 ± 0.02 to 1356.86 ± 461.81 mg. As shown in Figure 3, the linear regressions of TL, SL, and BH of the larvae and juveniles were plotted against age, with two inflexion points, respectively. However, the BW showed exponential growth. All correlation coefficients (R value) were greater than 0.90. The first inflexion point of TL, SL, and BH occurred at 25 dph with TL of 6.39 ± 0.73 mm, when weaning was initiated and growth accelerated. The slopes (mean growth rates) increased from 0.1452 to 0.8956 mm day^−1^ (TL), from 0.1357 to 0.7022 mm day^−1^ (SL), and from 0.0745 to 0.3777 mm day^−1^ (BH) (Figure 3, SW). The second inflexion point occurred at 40 dph with a TL of 18.81 ± 0.97 mm, when the larvae were fed the formulated feed and growth accelerated again, and the slopes increased to 1.1626 mm day^−1^ (TL), 0.9776 mm day^−1^ (SL), and 0.7852 mm day^−1^ (BH) (Figure 3, PW).
4. Discussion
4.1. Characteristics of Fertilized Eggs
The fertilized egg of S. cirrhifer was a demersal egg with stickiness, which is similar to that reported in the grass pufferfish Takifugu niphobles [31]. However, compared to S. cirrhifer, the stickiness of T. niphobles eggs was not strong. In this study, we observed that the initial eggs of S. cirrhifer had strong stickiness, which decreased after hatching, similar to that of the black scraper Thamnaconus modestus [32]. Furthermore, the first fertilized egg of S. cirrhifer contained multiple oil globules, while newly hatched larvae contained only a single oil globule. It was observed that the oil globules were initially distributed in the center of the egg, and they gradually moved towards the vegetal pole during the process of cleavage. They began to fuse during muscular contraction and melted into a larger oil globule during heart pulsation. Similar phenomena were observed in other species. For example, in the fat greenling Hexagrammos otakii [33], multiple oil globules melted into 1–5 oil globules, while they melted into 1–3 globules in the rabbitfish Siganus oramin [34]. Furthermore, in the medaka Oryzias melastigma, the fusion of oil globules into a single globule occurred during the transition from the blastocyst to gastrula stages [35]. In the T. modestus, oil globules began to fuse during the tail-bud stage, and they fused into a large oil globule during the muscular contraction stage [32], which was slightly earlier than in the S. cirrhifer observed in this study. Therefore, the fusion period of an oil globule may be regarded as an important characteristic during the embryonic development of the S. cirrhifer.
In Table 3, we compare the characteristics of embryo development between S. cirrhifer and other filefish species. In the present study, the S. cirrhifer egg diameter was similar to that of T. modestus [32], 23.2% smaller than that of Japanese inflator filefish Brachaluteres ulvarum [36], and 18.9% larger than that of leatherjacket Paramonacanthus japonicus and White-spotted pygmy filefish Rudariu ercodes [37]. The embryonic incubation time for S. cirrhifer was 4% shorter than that of T. modestus, with the shortest and longest times observed in P. japonicus and B. ulvarum, respectively. Among these five species, the embryonic developmental characteristics were most similar between S. cirrhifer and T. modestus.
4.2. Embryonic Development
In this study, the eye primordia of S. cirrhifer appeared before the formation of sarcomeres, which is similar to the American shad Alosa sapidissima [38] and rock bream Oplegnathus fasciatus [39]. However, differently, the eye primordia of pufferfish T. obscurus appeared after the formation of sarcomeres [40]. After 15 h 10 min of fertilization during the formation of segments, the Kupffer’s vesicle appeared above the tail of the embryo with a transparent spherical cyst. After 25 h 40 min, the tail bud of the embryo became dissociated, and the Kupffer’s vesicle disappeared. Studies have shown that the early appearance of the Kupffer’s vesicle may be related to the digestion and absorption of nutrients, which may promote the absorption of yolk [41]. The cells of Kupffer’s vesicle may participate in the formation of the embryonic digestive tract during embryonic development [42]. There are also reports that the appearance of Kupffer’s vesicle initiated asymmetric development of the embryo on both sides of the symmetry axis of the embryo, creating conditions for the formation of various organs [43,44]. However, the function and mechanism of the Kupffer’s vesicle during embryonic development of S. cirrhifer still remain unclear in this study.
The embryonic development of fish is influenced by various factors, such as the quality of eggs and external environment [45]. Studies have shown that during the breeding season, the supply of nutrition for parent fish, especially lipid nutrition, may directly affect embryonic development [46,47,48]. Furthermore, in the fish that spawned in batches, the quality of eggs appeared different. In this study, the quality of eggs in the early and late stages of spawning was poor, and the hatching rate of eggs was also greatly lower than that during the peak period. Studies show that the embryo development of fish in the early stages is very sensitive to changes in the external environment [49]. In this study, it was observed that most fertilized eggs that were not successfully hatched showed arrested development during the stages of the cleavage and early gastrula. They displayed uneven cleavage, decreased viscosity and transparency, and the surface of the fertilized egg turning white. Studies show that that the optimal incubation temperature for silver rasbora Rasbora argyrotaenia eggs is 30 °C, which shows the best results of the hatching and survival rate of larvae [50]. Increased temperature is beneficial for accelerating the hatching rate of fertilized eggs in an appropriate range, but excessively high temperature may increase the malformation rate of embryos [51,52]. Further studies should focus on the effects of water temperature on the embryo development in S. cirrhifer.
4.3. Growth of Larvae and Juveniles
Weaning is a process to gradually replace live feeds with artificial diets in fish larvae [53]; therefore, this is not a natural process but rather is imposed artificially by the aquaculturist. In practice, the weaning of most temperate marine fish species such as yellowtail kingfish Seriola lalandi is usually commenced after metamorphosis [54]. In the present study, our feeding strategy initiated B. plicatilis from 3 to 25 dph, A. salina nauplii from 25 to 40 dph, and weaning to the formulated feed from 40 dph. In our previous study on S. senegalensis, the weaning period is also from 40 dph [30]. However, it suggested that the best weaning was 16–22 dph in the golden pompano Trachinotus ovatus [55]. Therefore, weaning presented a bottleneck in S. cirrhifer culture, which should be focused on in a future study.
In this study, we found two distinct inflexion points occurring in the growth curves of S. cirrhifer larvae, which may be related to metamorphosis, weaning, and transitions in feeding habits. Studies show that most fish have growth inflexion points in the early stages such as the grey mullet Chelon labrosus [56], as well as our previous studies on S. cirrhifer and T. modestus [57]. Furthermore, previous studies have shown that the phenomenon of inflexion points in the early stages of fish is related to their survival strategies and developmental mechanisms during long-term evolution, aiming to ensure that the most important organs for early survival are prioritized for development. For example, in the chum salmon Oncorhynchus keta [58] and C. labrosus [56], they prioritize the development of sensory, feeding, respiratory, and motor organs.
In this study, the inflexion points of total length, standard length, and body height appeared at 25 dph and 40 dph, respectively, with an accelerated growth in these stages. At 25 dph, with the development of the fin as the main swimming organ, the fish developed from the larval to juvenile stages, showing a dietary transition from B. plicatilis to A. salina nauplii, and the growth accelerated. This phenomenon was also observed in the larval rearing of T. modestus [32], rockfish Sebastes schlegeli [59], and yellow catfish Pelteobagrus fulvidraco [60]. At 40 dph, the main digestive organs of juveniles had developed well and were able to feed on formulated feed. The food intake increased sharply, and the second growth acceleration period occurred. Therefore, it is suggested that the bait and changes in feeding habits have a significant impact on the growth rate of larval fish in S. cirrhifer.
5. Conclusions
We derived the following results: (1) S. cirrhifer had spherical, sticky, and transparent fertilized eggs with multiple oil globules, and oil globules fused at the stage of muscular contraction and melted into a larger oil globule during heart pulsation; (2) the embryonic development of S. cirrhifer was divided into eight phases from pre-cleavage to hatching, lasting 48 h, and the overall process is similar to that of T. modestus; (3) the weaning period occurred from 40 dph with feeding changing from live feeds to artificial diets; and (4) two inflexion points occurring in the growth curves of larvae and juveniles were found to be associated with metamorphosis and transitions in feeding habits.
As the global output and quality of S. cirrhifer gradually decrease, these results may provide guidance to improve artificial propagation and early life stage success in filefish species. Further studies are essential for understanding the mechanisms of growth and reproductive regulation from hatching to maturation in this species.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Nelson J.S. Grande T.C. Wilson M.V. Fishes of the World John Wiley & Sons Hoboken, NJ, USA 2016
- 2Alien G.R. Amaoka K. Anderson W.D.Jr. Bellwood D.R. Bohlke E.B. Bradbury M.G. Carpenter K.E. Caruso J.H. Cohen A.C. Cohen D.M. A checklist of the fishes of the South China Sea Raffles Bull. Zool.20008569667 Available online: https://www.science.nus.edu.sg/wp-content/uploads/sites/11/2024/05/s 08rbz 569-667-1.pdf(accessed on 31 August 2022)
- 3Nakabo T. Fishes of Japan: With Pictorial Keys to the Species Tokai University Press Tokyo, Japan 2022 Volume 2Available online: https://www.nhbs.com/fishes-of-japan-with-pictorial-keys-to-the-species-3-volume-set-japanese-book(accessed on 26 July 2024)
- 4Miyajima-Taga Y. Masuda R. Yamashita Y. Larvae of the threadsail filefish Stephanolepis cirrhifer feed on eggs and planulae of the jellyfish Aurelia sp. under laboratory conditions Plankton Benthos Res.201611969910.3800/pbr.11.96 · doi ↗
- 5Youn C.H. Fishes of Korea with Pictorial Key and Systematic List Academic Seojeok Seoul, Republic of Korea 2002 Volume 448
- 6Kim P.D. Miniature Guide for Whole Korean Fish Seoul Poong Deung Press Seoul, Republic of Korea 2007
- 7Minami T. Kanemaru M. Iwata K. Kuwahara M. Amano K. Mizuta A. Maeda N. Nishiki I. Tue Y. Yoshida T. Pathogenicity of Streptococcus iniae and Lactococcus garvieae in farmed thread-sail filefish and efficacy of the formalin-killed vaccines against these bacteria Fish Pathol.201348818710.3147/jsfp.48.81 · doi ↗
- 8Mizuno K. Miura C. Miura T. Relationships between Temperature and Growth of Thread-sail Filefish Stephanolepis cirrhifer and Black Scraper Thamnaconus modestus Aquacult. Sci.201462233010.11233/aquaculturesci.62.23 · doi ↗