Studies of Vietnamese Pteridophyte Flora 3

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3 Figure 4
Figure 4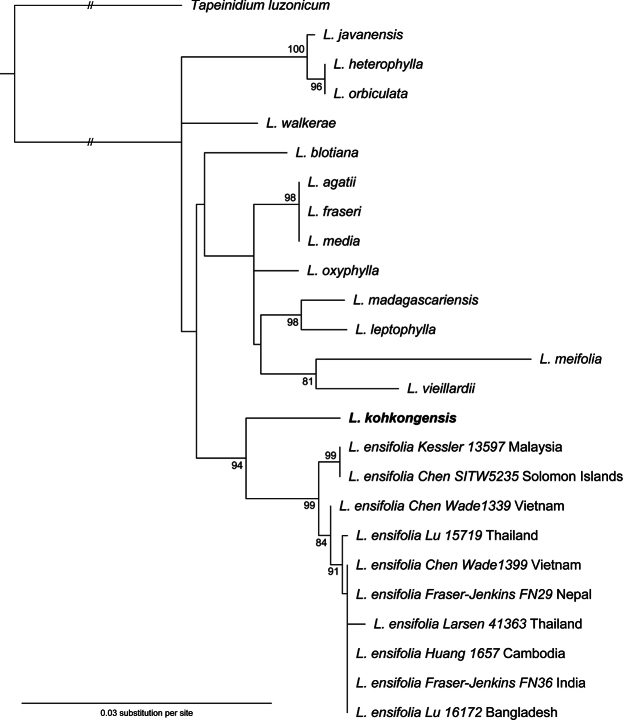 Figure 5
Figure 5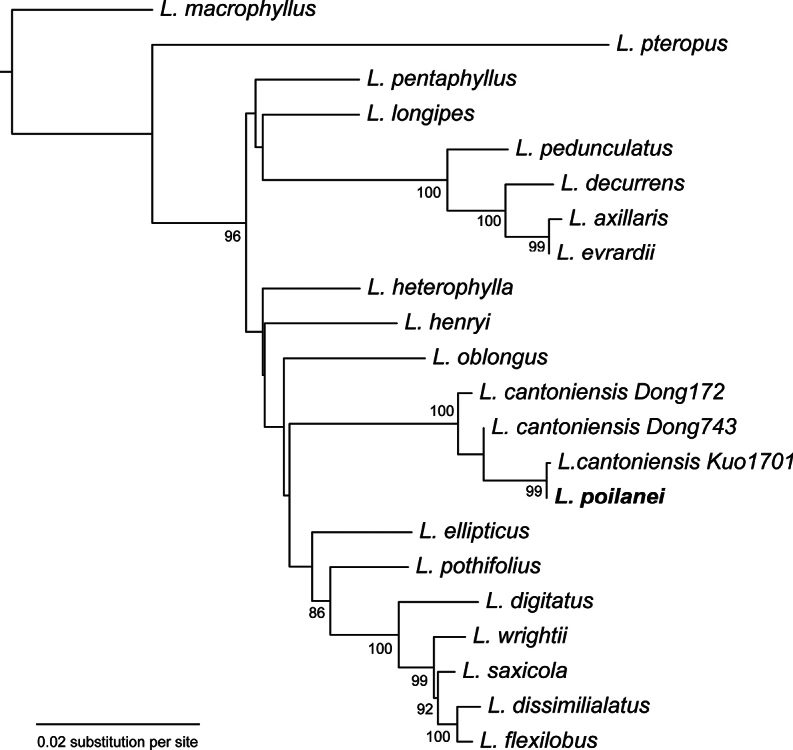 Figure 6
Figure 6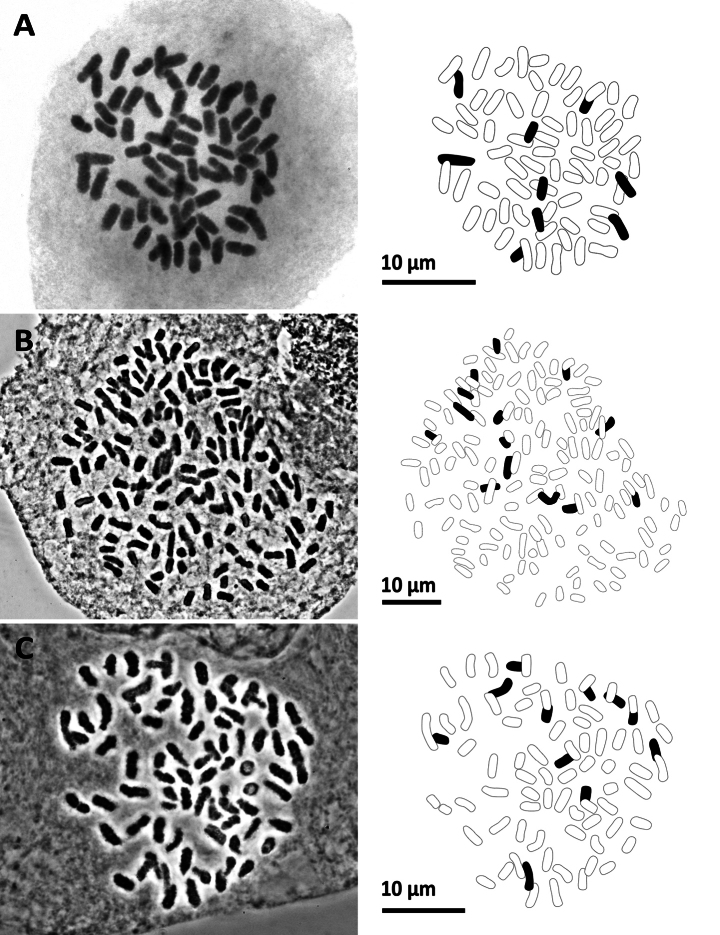 Figure 7
Figure 7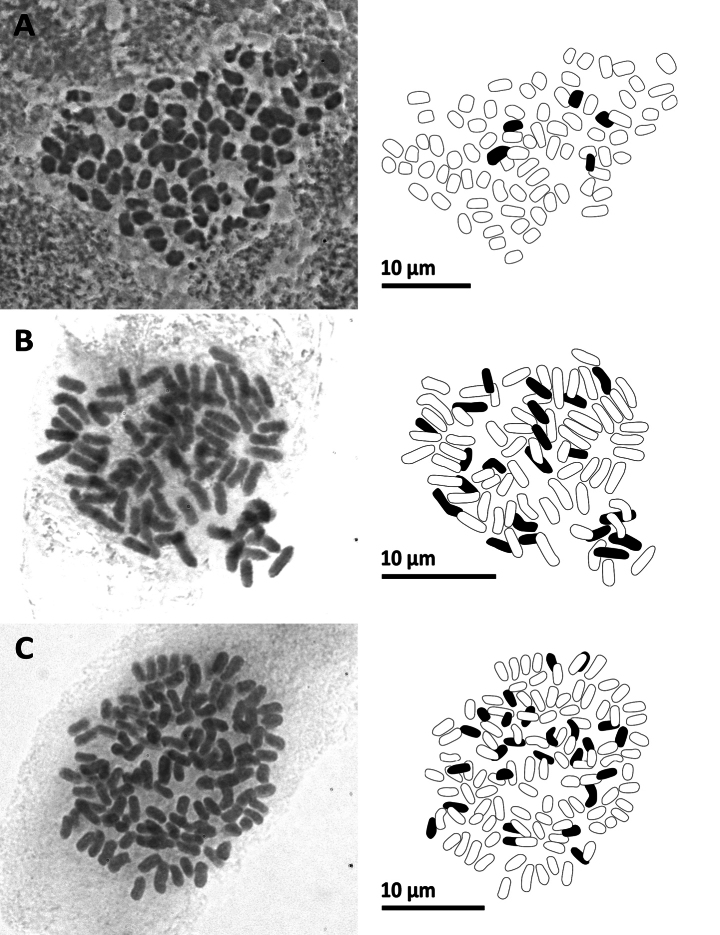 Figure 8
Figure 8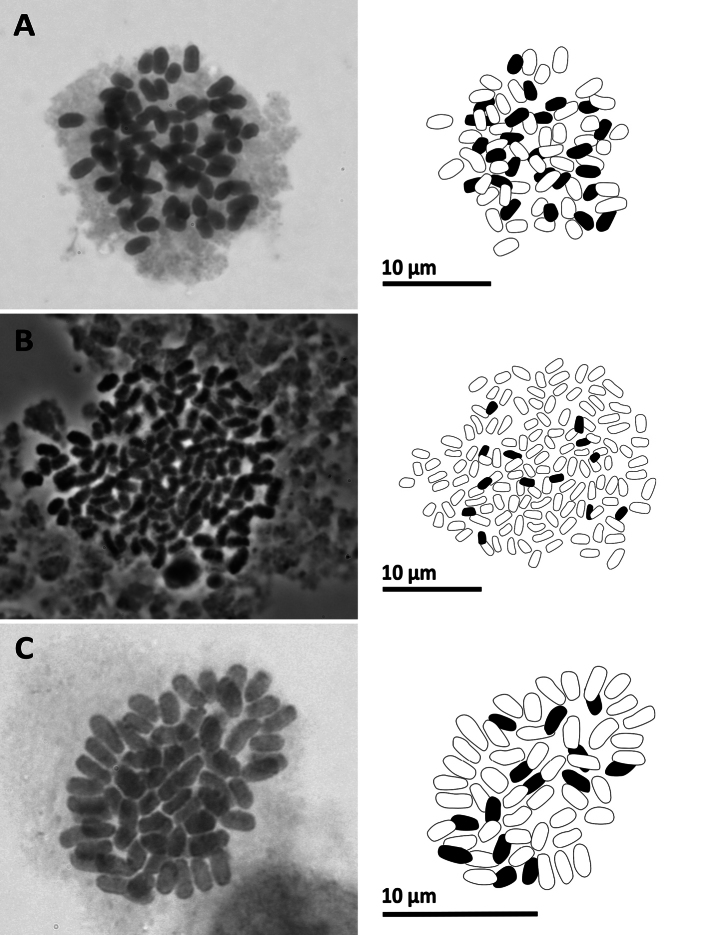 Figure 9
Figure 9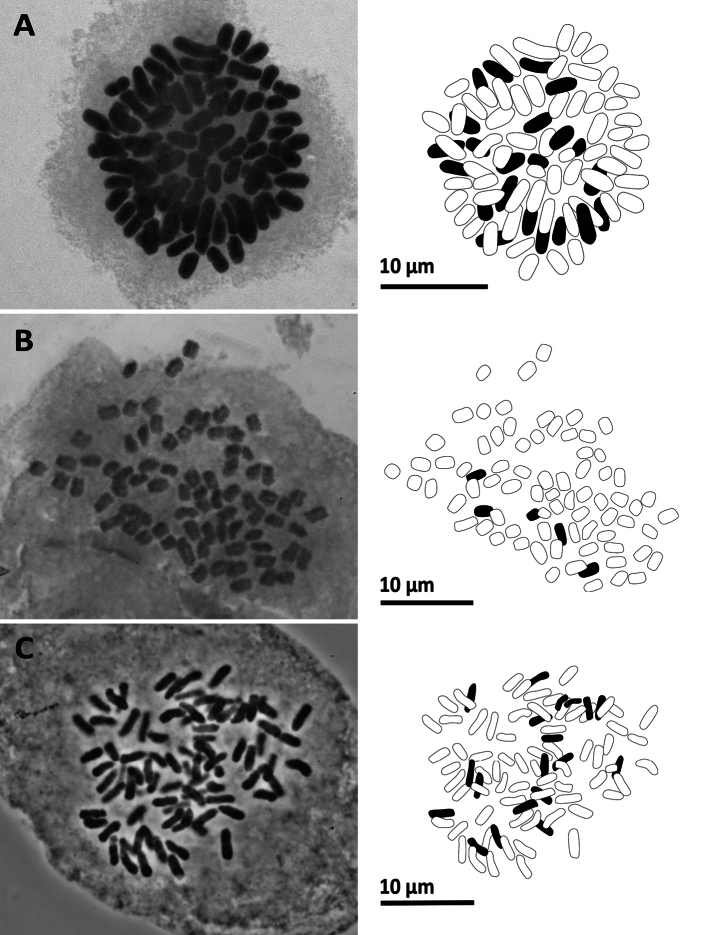 Figure 11
Figure 11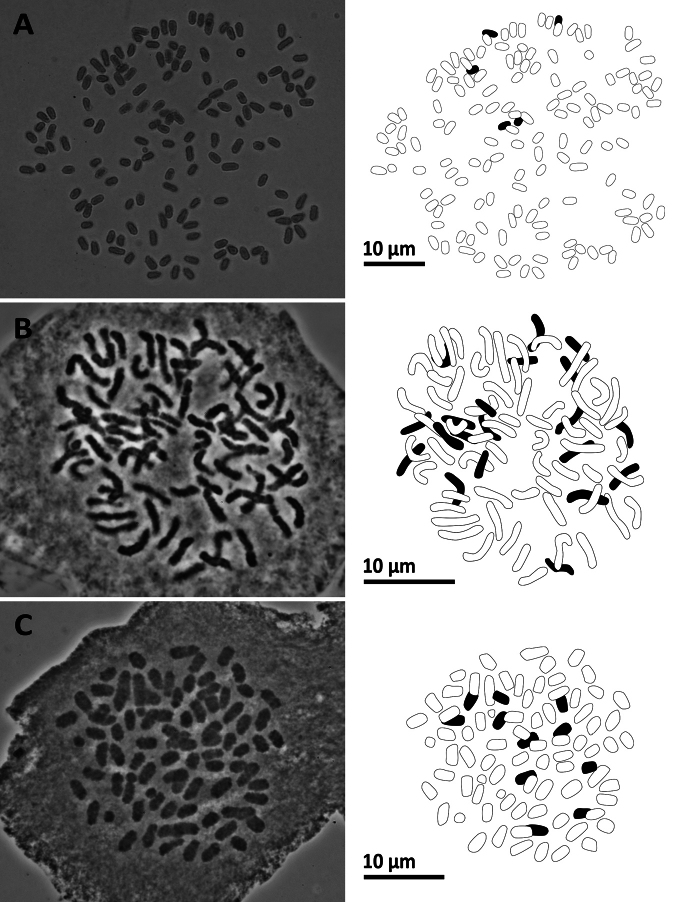 Figure 10
Figure 10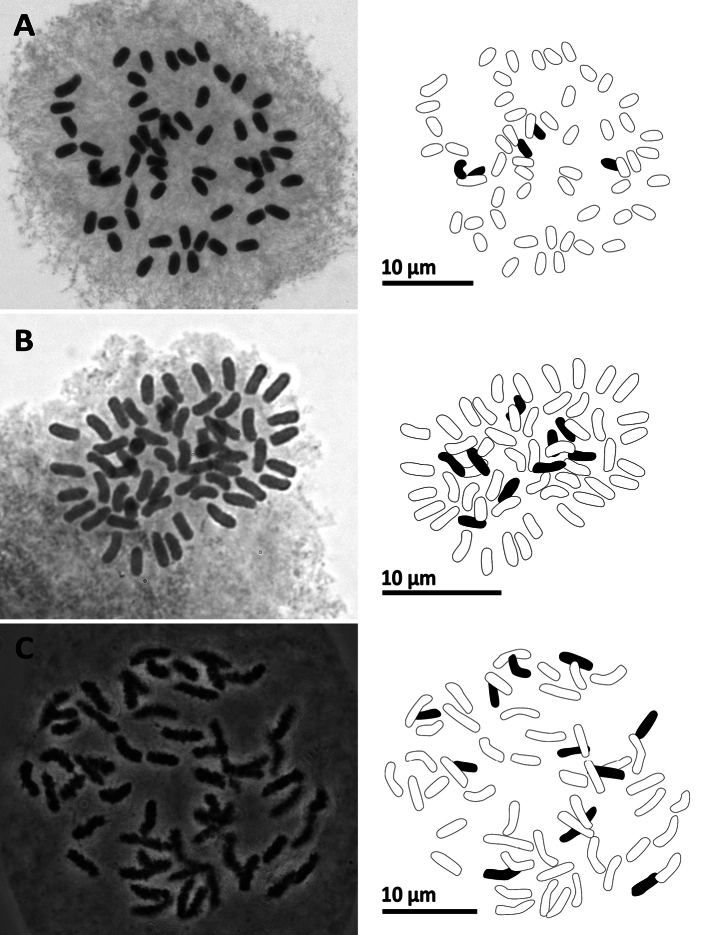 Figure 12
Figure 12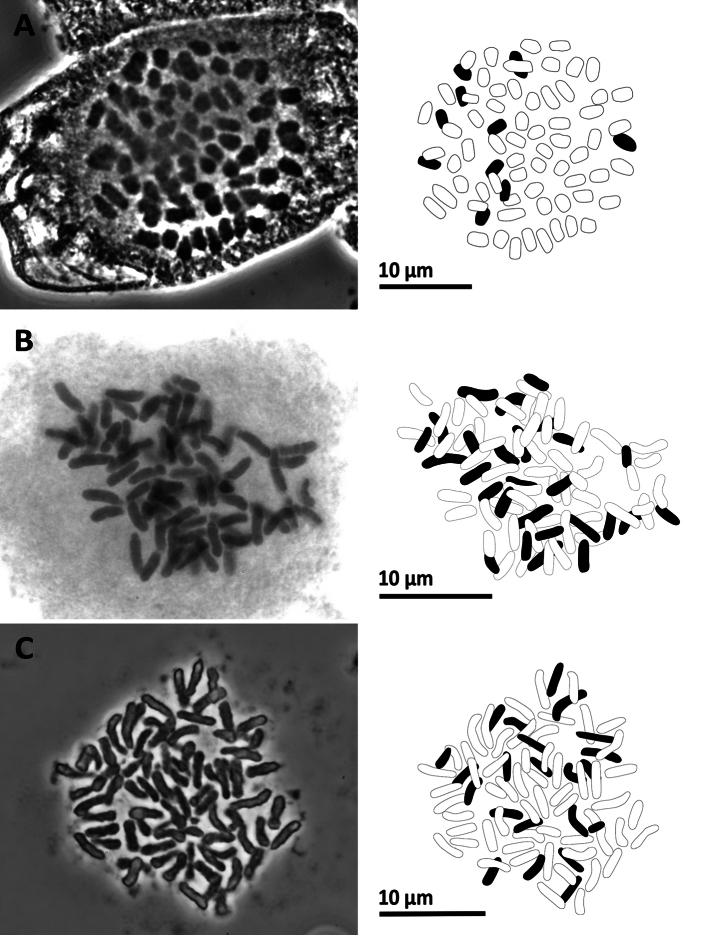 Figure 13
Figure 13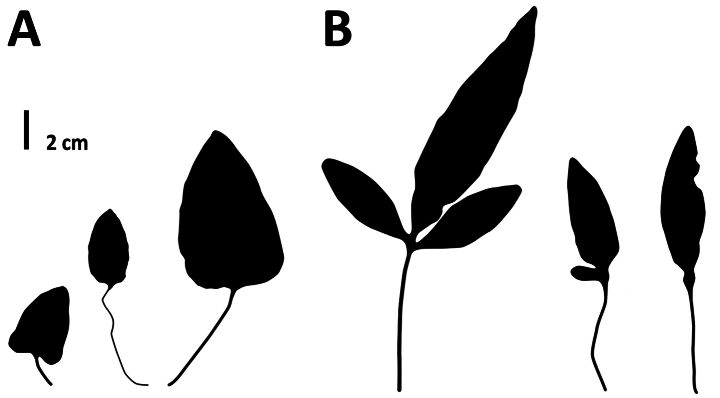 Figure 14
Figure 14| Region | Name | Sequence 5’ to 3’ | Reference |
|---|---|---|---|
|
| FERN matK fEDR | ATTCATTCRATRTTTTTATTTHTGGARGAYAGATT |
|
|
| DeLin matK rNRD | CTACGCAAYSCATCYCGATTT |
|
|
| 1F | ATGTCACCACAAACAGAAAC |
|
|
| 1379R | TCACAAGCAGCAGCTAGTTCAGGACTC |
|
|
| rps5 | ATGTCCCGTTATCGAGGACCT |
|
|
| trnSR | TACCGAGGGTTCGAATC |
|
|
| trnH | CGCGCATGGTGGATTCACAATCC |
|
|
| psbA3’f | GTTATGCATGAACGTAATGCTC |
|
|
| FernL1Ir1 | GGYAATCCTGAGCCAAATC |
|
|
| F | ATTTGAACTGGTGACACGAG |
|
| Taxa | Voucher | Chromosome no. | Ploidy | s/s | Figure |
|---|---|---|---|---|---|
|
| 2 | 2x | 64 | 7A | |
|
| 2 | 4x | 64 | 7B | |
|
| 2 | 2x | — | 7C | |
|
| 2 | 2x | 64 | 8A | |
|
| 2 | 2x | 64 | 8B | |
|
| 2 | 3x | — | 8C | |
|
| 2 | 2x | 64 | 9A | |
|
| 2 | 2x | 64 | 9B | |
|
| 2 | 2x | — | 9C | |
|
| 2 | 4x | 64 | 10A | |
|
| 2 | 2x | 64 | 10B | |
|
| 2 | 2x | 64 | 10C | |
|
| 2 | 2x | — | 11A | |
| 2 | 3x | 32 | 11B, 11C | ||
|
| 2 | 2x | 64 | 12A | |
|
| 2 | 2x | 32 | 12B | |
|
| 2 | 2x | 32 | 12C | |
|
| 2 | 2x | — | 13A | |
|
| 2 | 2x | — | 13B | |
|
| 2 | 2x | 64 | 13C |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsFern and Epiphyte Biology · Plant Diversity and Evolution · Plant and Fungal Species Descriptions
Introduction
This marks the third paper in our series on Vietnam’s pterido-flora (Chen et al. 2021; 2023). This series of studies aims to provide updated knowledge of the country’s pteridophyte flora, based on recent expeditions as well as the studies of herbarium specimens and relevant literature. Here, we reported three new additions to Vietnam’s flora: Haplopterisyakushimensis C.W.Chen & Ebihara (Fig. 1), Lindsaeakohkongensis I.C.Hwang, M.O.Moon & B.Y.Sun (Fig. 2), and Pterispseudowulaiensis Y.S.Chao (Fig. 3) based on our new collections.
Haplopterisyakushimensis C.W.Chen & Ebihara (Chen Wade6952) A habit B rhizome scales C adaxial lamina D abaxial lamina E stipes F gametophytes. Photographed by C.-W. Chen.
Lindsaeakohkongensis I.C.Hwang, M.O.Moon & B.Y.Sun (Chen Wade5034) A habit B rhizome C adaxial lamina D abaxial lamina E venation F rhizome scales. Photographed by C.-W. Chen.
Comparison of Pterispseudowulaiensis C.M.Kuo (A, B, C, Chao 3509 and 3517) and P.oshimensis Hieron. (D, Bon 4758) A habit B lamina (backlight) C specimen D specimen, CC BY © The Trustees of the Natural History Museum, London. Photographed by C.-W. Chen and Y.-S. Chao.
Additionally, we performed phylogenetic analyses to elucidate the placements of Lindsaeakohkongensis (Fig. 2) and Leptochiluspoilanei (C.Chr. & Tardieu) Liang Zhang & Li Bing Zhang (Fig. 4). In 2018, we collected an unknown Lindsaea specimen from Phu Quoc Island of southern Vietnam. This specimen closely resembles L.ensifolia Sw. but being much smaller and with an unusual subaquatic habit. Specimens from Cambodia and Malaysia with the same morphology had recently been described as Lindsaeakohkongensis (Yun et al. 2023) but without molecular data. Here, we conducted a phylogenetic analysis to test whether L.kohkongensis is a distinct species or an eco-form of L.ensifolia.
Leptochiluspoilanei (C.Chr. & Tardieu) Liang Zhang & Li Bing Zhang (Chen Wade6804) A habitat B habit C rhizome scales D sterile frond E fertile frond F cross section of rhizome. Photographed by C.-W. Chen.
Leptochiluspoilanei was described by Christensen and Tardieu-Blot (1939) under the genus Colysis (≡ Leptochilus), based on specimens collected from Annam, which is now part of central Vietnam. Following its initial description, the species remained largely unstudied until Zhang et al. (2018) reclassified it under Leptochilus. To our knowledge, this species is only known from its type collection, and our new collection (Fig. 4) from Nui Chua National Park represents the second collection of this species. It can be readily distinguished from other congeneric species in Vietnam by its strongly dimorphic fronds: fertile fronds are mostly linear and simple (rarely tripartite), while sterile ones are simple to pinnatifid.
Lastly, we presented new chromosome counts of Vietnamese fern species and explored the implications of our findings in the context of systematics. Chromosome numbers have long been recognized as important information in plant systematics (Rice et al. 2015). This significance is particularly pronounced for ferns, as they exhibit the highest frequency of polyploid speciation among vascular plants (Wood et al. 2009). Despite the importance of chromosome number data, research on Vietnamese ferns in this regard has been limited. To date, no systematic survey has been conducted. Instead, available data remains sporadic and dispersed throughout various articles, often in the context of new species’ descriptions (e.g., Chen et al. 2023), new species records (e.g., Ding et al. 2013), or cytological studies focused on specific taxa, including specimens collected from Vietnam (e.g., Nayak and Singh 1989). By reporting the chromosome numbers of Vietnamese fern species and demonstrating how these data can be integrated into systematic studies, we aim to raise awareness of chromosome studies among local botanists, thereby encouraging further related research in the future.
Materials and methods
Specimen identification and distribution
We conducted field expeditions in Bidoup Nui-Ba National Park, Cuc Phuong National Park, Phia Oac-Phia Den National Park, Phu Quoc National park, and Nui Chua National Park during 2018–2023. To ensure the correct identifications of our newly collected specimens, we compared them with herbarium specimens, relevant literature, including published papers, checklist, and flora from neighboring countries (see notes under each species), and original protologues and types through the Biodiversity Heritage Library (https://www.biodiversitylibrary.org/) and JSTOR Global Plants (https://plants.jstor.org/). To confirm the known distribution of each species, we searched the names against World Ferns (Hassler 1994–2024) and GBIF (https://www.gbif.org/) and manually examined the available specimen images.
Cytological observations
To determine chromosome numbers, root tips were collected either from the field or transplanted plants in greenhouses. These root tips were treated with a 1:1 mixture of hydroxyquinoline and cycloheximide (Sigma-Aldrich, USA) for approximately 16 hours at 18 °C. Following this, the root tips were fixed in a 3:1 mixture of 95% ethanol and 45% acetic acid for about 12 hours at room temperature. Subsequently, the root tips were macerated using a 1:1 mixture of cellulase (Yakult, Japan) and pectolyase (Sigma-Aldrich, USA) for 1 hour at 37 °C. Finally, the treated root tips were squashed in 2% acetocarmine and observed under a microscope (Zeiss Axio Imager A1, Germany). To determine the ploidy of our observed chromosome numbers, we consulted two online databases, Chromosome Counts Database (CCDB, Rice et al. 2015) and Index to Plant Chromosome Numbers (IPCN, Goldblatt and Lowry 2011), and considered the lowest sporophytic counts known for a genus as the diploid.
Furthermore, we counted the spore numbers per sporangium and observed the spore shape regularity to determine the reproductive modes whenever possible. We also compared the spore sizes of Leptochiluscantoniensis (Baker) Ching and L.poilanei by measuring 30 spores from each species.
Molecular phylogenetic analysis
To elucidate the phylogenetic placement of Lindsaeakohkongensis and Leptochiluspoilanei, we conducted two phylogenetic analyses each based on three chloroplast markers including matK, trnH-psbA, and trnL-F for Lindsaea and rbcL, rps4-trnS, and trnL-F for Leptochilus. These markers were chosen to integrate our newly generated sequences into previous studies (Lehtonen et al. 2010; Zhang et al. 2019; Chen et al. 2020a; Fujiwara et al. 2023). We extracted genomic DNA from fresh fronds using Qiagen DNeasy Plant Mini Kit (Hilden, Germany), following the manufacturer’s protocol. We conducted PCR to amplify the five DNA markers using the primers listed in Table 1.
For Lindsaea, we included the sequences of 26 specimens as listed in Appendix 1: Table A1. Among these, the sequences of 14 specimens were downloaded from GenBank. To test the monophyly of L.ensifolia, this sampling included 1) L.ensifolia from a broad geographic range, including Bangladesh, Brunei, Cambodia, India, Malaysia, Nepal, Solomon Islands, Thailand, and Vietnam; and 2) closely related species identified by Lehtonen et al. (2010). For Leptochilus, in addition to our newly sequenced L.poilanei, we download the sequences of another 21 specimens from GenBank. In total, 22 specimens representing 19 species were included in the phylogenetic analysis as listed in Appendix 1: Table A2. This sampling covered all the major clades found in previous studies (Zhang et al. 2019; Chen et al. 2020a).
For our newly generated sequences, we first manually inspected raw reads, removing any ambiguous bases using BioEdit (Hall 1999). Subsequently, we aligned the sequences of each marker using MUSCLE (Edgar 2004) with default settings. These individual marker alignments were then concatenated into a single alignment, because chloroplast genome is non-recombining and therefore has a single evolutionary history. We then conducted maximum likelihood (ML) analyses using IQTREE (Minh et al. 2020) each with five partitions (three codon positions for matK and rbcL, and rps4-trnS, trnH-psbA and trnL-F, each treated as a single partition for simplification although they contain partly coding regions). The best-fit model for each partition and the best-fit partition scheme were determined by ModelFinder (Lanfear et al. 2012; Kalyaanamoorthy et al. 2017) as implemented in IQTREE (Minh et al. 2020). To assess branch support, we performed 1000 ultrafast bootstrap replicates using UFBoot (Hoang et al. 2018). The two concatenated alignments and the resulting phylogenetic trees are available on the Dryad Digital Repository (Chen et al. 2024).
Results
Phylogenetic placement of Lindsaeakohkongensis
The concatenated three-marker alignment contains 1832 bp including 85 parsimony informative sites and 39.2% missing data (17,973 bp). ModelFinder merged all the five partitions into one alignment with the best-fit model identified as K3Pu+F+R2. The phylogram generated by the ML analysis based on the concatenated alignment is shown in Fig. 5. The inferred species relationships closely align with the phylogeny reconstructed by Lehtonen et al. (2010) although the nodes are poorly supported in general. However, Lindsaeakohkongensis is strongly supported as the sister group to a clade comprising all the L.ensifolia specimens included in this analysis.
Phylogram of Lindsaea reconstructed from the maximum likelihood analysis of the concatenated plastid dataset (matK, trnH-psbA, and trnL-F). The branch length to the outgroup Tapeinidium is not to scale for better visualization. Support values below 80 are not shown on the nodes. L.kohkongensis is indicated in bold.
Phylogenetic placement of Leptochiluspoilanei
The concatenated three-marker alignment contains 3331 bp including 248 parsimony informative sites and 17.8% missing data (13077 bp). ModelFinder subset the alignment into two partitions, the first including first and third codon position of rbcL, and the second including the second codon position of rbcL along with rps4-trnS and trnL-F. The best-fit models determined for the two partitions were TNe+I+G4 and K3Pu+F+R2, respectively. The phylogram generated by the ML analysis based on the concatenated alignment is shown in Fig. 6. The inferred species relationships are similar to previous studies (Zhang et al. 2019; Chen et al. 2020a) as expected, although poorly supported in general. Leptochiluspoilanei is nested in a highly supported clade containing three specimens of L.cantoniensis.
Phylogram of Leptochilus reconstructed from the maximum likelihood analysis of the concatenated plastid dataset (rbcL, rps4-trnS, and trnL-F). Support values below 80 are not shown on the nodes. L.poilanei is indicated in bold.
Both L.cantoniensis (based on Hsu 3399) and L.poilanei (based on Chen Wade6804) produce 64 spores in each sporangium with normal morphology. The spore size of L.cantoniensis and L.poilanei is 44.3 ± 3.4 and 63.0 ± 4.9 µm, respectively. The spores of L.poilanei are significantly larger than those of L.cantoniensis (t-test, p < 0.001).
Chromosome number of 20 species
We successfully counted the chromosome numbers of 20 species, each determined by multiple cells. Their chromosome numbers and ploidy levels inferred from the known lowest base number are shown in Table 2. Figs 7–13 illustrated representative cells for all the 20 species. Among these, chromosome numbers were recorded for the first time for seven species: Gymnosphaerasalletii (Tardieu & C.Chr.) S.Y.Dong, Lepisorusspicatus (L.f.) Li Wang, Leptochiluspoilanei, Pteridryscostularis Li Bing Zhang, Liang Zhang, N.T.Lu & X.M.Zhou, Pterislatipinna Y.S.Chao & W.L.Chiou, Pyrrosiaeberhardtii (Christ) Ching, and Tectariasetulosa (Baker) Holttum. Furthermore, we reported the new cytotypes for three species: Diplaziumdoederleinii (Luerss.) Makino, Pterisesquirolii H.Christ, and Tectariaharlandii (Hook.) C.M.Kuo.
Mitotic chromosomes with explanatory illustrations AAspleniumnormale (2n = 72) BAspleniumtenerum (2n = 144) CCtenitiseatonii (2n = 82).
Mitotic chromosomes with explanatory illustrations ADidymochlaenatruncatula (2n = 82) BDiplaziumdoederleinii (2n = 82) CDiplaziumdonianum (2n = 123).
Mitotic chromosomes with explanatory illustrations AGrypothrixsimplex (2n = 72) BGymnosphaerasalletii (2n = 138) CLepisorusspicatus (2n = 70).
Mitotic chromosomes with explanatory illustrations APteridryscostularis (2n = 82) BPteriscadieri (2n = 87) CPteriscadieri (2n = 87).
Mitotic chromosomes with explanatory illustrations ALeptochiluspoilanei (2n = 144) BPleocnemiawinitii (2n = 82) CPolystichumbiaristatum (2n = 82).
Mitotic chromosomes with explanatory illustrations APterisesquirolii (2n = 58) BPterislatipinna (2n = 58) CPterispseudowulaiensis (2n = 58).
Mitotic chromosomes with explanatory illustrations APyrrosiaeberhardtii (2n = 74) BTectariaharlandii (2n = 80) CTectariasetulosa (2n = 80).
Discussion
Systematic uniqueness of Lindsaeakohkongensis
When we collected this species from Phu Quoc island in 2018, we were hesitant to describe it. Morphologically, it closely resembles L.ensifolia, leading us to question if it might be an eco-form of the latter. In this study, we conducted a phylogenetic analysis based on three chloroplast regions, sampling L.ensifolia from a wide geographic distribution throughout Asia and the Pacific Islands. The results indicate that L.kohkongensis is sister to L.ensifolia rather than being embedded within it (Fig. 6). On one hand, this suggests that L.kohkongensis can be recognized as a distinct species based on the principle of monophyly. On the other hand, it could also be classified within a broadly circumscribed L.ensifolia. To our knowledge, L.ensifolia typically grows terrestrially in a diverse habitat from swampy forests, dipterocarp forests, to exposed grasslands (Kramer 1971; personal observation). In contrast, L.kohkongensis is exclusively found in shaded valleys as lithophytes and is frequently submerged in water (Yun et al. 2023; personal observation). This distinction suggests an ecological speciation event that needs to be tested in the future.
While identifying our specimen from Phu Quoc island, we came across a rheophyte form of L.ensifolia described as L.ensifoliavar.rheophila K.Iwats from Sumatra (Iwatsuki 1977). After examining the type materials deposited in KYO, we concluded that this is the same species as L.kohkongensis and should be treated as a synonym of it. As noted by Kramer (1967), L.ensifolia is one of the most widespread and variable species of the genus. Apart from var. rheophila, few other subspecies or varieties have been published, such as subsp. coriacea (Alderw.) K.U.Kramer from Bornean swamp forests. Furthermore, ploidy variations (diploid, triploid, and tetraploid) and different reproductive modes (sexual and apomixis) have been observed in L.ensifolia (e.g., Lin et al. 1996). Although we here tentatively recognize L.kohkongensis as a distinct species, the variability observed in L.ensifolia underscores the need for a systematic study of this complex species.
Hybrid origin hypothesis of Leptochiluspoilanei
Our phylogenetic analysis based on three plastid markers unambiguously resolves L.poilanei within a clade that includes three specimens of L.cantoniensis (Fig. 6). This result is unexpected, considering the clear morphological distinctions between these two species. Specifically, fully developed sterile fronds of L.poilanei are pinnatifid, while those of L.cantoniensis remain consistently simple (Fig. 14).
Comparison of the sterile fronds between Leptochiluscantoniensis and L.poilaneiAL.cantoniensis, based on Kuo 1701 (TAIF [509328], left), Cadière 158 (MICH [1191289], middle), and s.c., s.n. (K [000959730], right) BL.poilanei, based on Poilane 5373 (BM [000036782], all the three fronds).
Our examination of the reproductive mode indicates that both species undergo sexual reproduction, as evidenced by the production of 64 well-formed spores in each sporangium. The mitotic chromosome count for L.poilanei is 2n = 144 (Fig. 10A), suggesting it is a tetraploid, given the base number of the genus is x = 36 (Rice et al. 2015). Although the chromosome number of L.cantoniensis is presently unknown, its spores are significantly smaller than those of L.poilanei (44.3 ± 3.4 vs. 63.0 ± 4.9 µm). Considering the widely found correlation between ploidy and spore size in ferns (Barrington et al. 1986), we propose that L.cantoniensis is likely a diploid species.
Building on the evidence from our study, we propose that L.poilanei has a hybrid origin. We suggest that L.poilanei originated from hybridization between L.cantoniensis (as the maternal parent, considering chloroplast inheritance in ferns, e.g., Gastony and Yatskievych 1992) and an unidentified diploid species, followed by polyploidization. The close similarity in chloroplast DNA markers between L.cantoniensis and L.poilanei suggests a relatively recent hybridization event. This hypothesis gains further support from the overlapping distribution of L.cantoniensis and L.poilanei in central Vietnam. While the identity of the other parent remains elusive, we anticipate it is likely a species with pinnatifid fronds, given that L.poilanei exhibits this characteristic. Future studies employing bi-parentally inherited nuclear markers and comprehensive sampling of the genus in Vietnam are essential to test our hypothesis.
New findings of chromosome number
In the following paragraphs, we discussed the systematic implications of our new finding (i.e., new species counts or new cytotypes) species by species, following alphabetical order.
Diplaziumdoederleinii (Luerss.) Makino—2n = 82, sexual diploid, Fig. 8B
Previous studies reported triploid (2n = 123) and tetraploid (2n = 164) specimens from Japan (Takamiya et al. 2001). Our discovery of a diploid specimen from Vietnam provides further support for the long-observed cytogeographic pattern: diploids are typically found in warmer tropics, whereas polyploids are prevalent in cooler temperate regions (e.g. Chang et al. 2013). As demonstrated by Takamiya et al. (2001), there is a little genetic divergence between triploids and tetraploids. Future studies could incorporate the newly discovered diploid into genetic analyses to further investigate the origins of polyploids.
Gymnosphaerasalletii (Tardieu & C.Chr.) S.Y.Dong—2n = 138, sexual diploid, Fig. 9B
The first chromosome count reported for the species. This Vietnam endemic species was recently reevaluated by Li et al. (2024), who described two new species previously been misidentified as G.salletii. Gymnosphaera is predominantly a paleotropical genus, comprising approximately 46 species (Dong and Zuo 2018). All four previously reported species with chromosome counts in the genus are diploid, namely G.capensis (L.f.) S.Y.Dong, G.gigantea (Wall. ex Hook.) S.Y.Dong, G.khasyana (T.Moore ex Kuhn) Ching, and G.podophylla (Hook.) Copel. (Manton and Sledge 1954; Löve et al. 1977; Kato 1999). Notably, there is a correlation between geographic distribution and the basic chromosome number: all Asian species exhibit x = 69, while African species have x = 70.
Lepisorusspicatus (L.f.) Li Wang—2n = 70, diploid, Fig. 9C
The first chromosome count reported for the species. Found across the paleotropics, this species has often been confused with L.mucronatus (Fée) Li Wang. Following Hovenkamp (1998), we identify the species by the presence of entire rhizome scales with hyaline margins. Following Zhao et al. (2020), this species belongs to sectiob Belvisia (Mirb.) C.F. Zhao, R.Wei & X.C. Zhang, a small clade comprising eight species (Hovenkamp and Franken 1993). Lepisorusmucronatus is the only other species in this section with known chromosome numbers and has been determined as tetraploid (2n = 140) from Australia (Tindale and Roy 2002).
Leptochiluspoilanei (C.Chr. & Tardieu) Liang Zhang & Li Bing Zhang—2n = 144, sexual tetraploid, Fig. 10A
As discussed in previous sections, we hypothesize that this Vietnam endemic species originated from a hybridization between L.cantoniensis and an unidentified diploid species, followed by polyploidization. Leptochilus is predominantly a tropical Asian genus with approximately 51 species (Zhang et al. 2024). Prior to this study, chromosome numbers were known for only four species including: L.axillaris (Cav.) Kaulf., L.decurrens Blume, L.ellipticus (Thunb.) Noot., and L.pothifolius (Buch.-Ham. ex D.Don) Fraser-Jenk. (e.g., Mitui 1968; Löve et al. 1977; Manickam and Irudayaraj 1989). Among these four species, L.decurrens is the only one reported to exist in both diploids and tetraploids, while the other three species are known only as diploids.
Pteridryscostularis Li Bing Zhang, Liang Zhang, N.T.Lu & X.M.Zhou—2n = 82, diploid, Fig. 11A
The first chromosome count reported for the species. Pteridrys is a tropical Asian genus with ca. 22 species, and 11 of them are recorded in Vietnam (Zhou and Zhang 2019). So far, all the four species (the other three are P.australis Ching ex C.Chr. & Ching, P.cnemidaria (Christ) C.Chr.et.Ching, and P.syrmatica (Willd.) C.Chr. & Ching) with reported chromosome numbers are diploids (Manton 1954; Tsai and Shieh 1985; Bidin and Go 1995).
Pterisesquirolii H.Christ—2n = 58, sexual diploid, Fig. 12A
This species is reported from southern China, Taiwan, and Vietnam (Chao 2019). Prior to this study, the only reported chromosome count was 2n = ca. 90 (presumably an apogamous triploid due to the count of 32 spores per sporangium) from southern China (Lin et al. 2002). Although Chao (2019) found very little genetic divergence among populations from China and Taiwan, it may be worthwhile to include the Vietnamese population in future analyses, given their different ploidy level and reproductive mode. This is particularly important considering the prevalence of reticulate evolution observed in the genus (e.g., Chao et al. 2012, 2022).
Pterislatipinna Y.S.Chao & W.L.Chiou—2n = 58, apogamous diploid, Fig. 12B
This is the first chromosome count reported for this species. This species was firstly described from Taiwan (Chao et al. 2017) and later reported from China and Vietnam (Chen et al. 2020b). Our chromosome counts support the hypothesis proposed by Chao et al. (2022) that P.latipinna is a diploid, as indicated by flow cytometry. Our spore number counts also confirm that P.latipinna is reproduced through apogamous reproduction, which is common for the genus (Walker 1962).
Pyrrosiaeberhardtii (Christ) Ching—2n = 74, diploid, Fig. 13A
The first chromosome count reported for the species. This species is recorded in southern China, Thailand, and Vietnam, with its type locality in Vietnam. It has sometimes been grouped under a broadly defined P.lingua (Thunb.) Farw. alongside other closely related species such as P.oblonga Ching and P.heteractis (Mett.) Ching. (Hovenkamp 1986). This broader classification of P.lingua is further supported by Zhou et al. (2017), showing a close relationship among these species. The chromosome numbers of P.lingua have been reported from China and Japan, and both are diploids (e.g., Takei 1969; Kato 1999).
Tectariaharlandii (Hook.) C.M.Kuo—2n = 80, diploid, Fig. 13B
This species has been documented in the Ryukyu Islands, southern China, northern Vietnam, and Taiwan. Tetraploid populations (n = 80) have been reported in the Ryukyu Islands (as Hemigrammadecurrens (Hook.) Copel., Mitui 1976) and Taiwan (Tsai and Shieh 1985). In our study, we provide the first record of a diploid form Vietnam. Notably, this species has been proposed as the maternal parent of the triploid (2n = 120) sterile hybrid species T.×hongkongensis (Zhao and Dong 2016), with T.zeilanica (Houtt.) Sledge as the paternal parent. It is reasonable to assume that T.×hongkongensis originated from the hybridization between diploid and tetraploid parents. Given that both diploids and tetraploids are now confirmed in these two parental species (Manton and Sledge 1954; Tsai and Shieh 1985), it would be interesting to investigate whether there is a bias in the direction of hybridization based on ploidy level.
Tectariasetulosa (Baker) Holttum—2n = 80, sexual diploid, Fig. 13C
The first chromosome count reported for the species. Initially described from Ba Vi mountain range in northern Vietnam, this species has since been documented in southern China, Indochina, and Peninsular Malaysia. Additionally, a variety raciborskii (Alderw.) Holttum has been further identified extending to Java without known ploidy.
New records from Vietnam
Haplopteris
yakushimensis
Taxon classificationPlantaePolypodialesPteridaceae
C.W.Chen & Ebihara, Phytotaxa 156(4): 232. 2014.
9C99E1EE-788C-56A3-AB12-8D65D377F4FF
Type.
Japan. • Kagoshima Pref., Yakushima Island, Nakabase River, 20 Aug 1982, Nakaike s.n. (holotype TNS [VS-456666!]).
Distribution and ecology.
This species was previously recorded only in Japan and Taiwan (Chen et al. 2014, TPG 2019). In Vietnam, H.yakushimensis is found in damp evergreen broadleaf forests at elevations of 1200–1360 m, where it grows on rocks with thick compost near streams.
Specimens examined.
Vietnam. • Cao Bang Province: Phia-Oac Phia-Den National Park, 6 Dec 2013, Zhang et al. 6755 (MO, TAIF [499240!], VNMN). Phia-Oac Phia-Den National Park, Nguyen Binh District, Thanh Cong Ward, 22.589539°N, 105.880403°E, 1353 m, 8 Nov 2023, Chen Wade6952 (TAIF!, VNMN!). • Ha Giang Province: Vi Xuyen District, Cao Bo Ward, 22.767500°N, 104.880394°E, 1200–1360 m, 12 Sep 2000, Harder et al. 5514 (UC [1763099!]).
Note.
Haplopteris, a fern genus comprising approximately 40 species, is primarily found in tropical Africa, Asia, and the Pacific Islands (Schuettpelz et al. 2016). In Vietnam, eight Haplopteris species were previously recorded: H.angustifolia (Blume) E.H.Crane, H.doniana (Mett. ex Hieron.) E.H.Crane, H.elongata (Sw.) E.H.Crane, H.ensata (Christ) C.W.Chen & S.Linds., H.ensiformis (Sw.) E.H.Crane, H.flexuosa (Fée) E.H.Crane, H.hainanensis (C.Chr. ex Ching) E.H.Crane, and H.sikkimensis (Kuhn) E.H.Crane (Chen et al. 2023). Haplopterisyakushimensis can be distinguished from these species by having fronds broader than 1 cm wide, costae that are grooved on the adaxial side and raised on the abaxial side, and the submarginal (ca. 1–2 mm away from the margins) sori lines.
Recent studies have identified two genetically distinct yet morphologically indistinguishable lineages in H.yakushimensis (Kuo et al. 2017; TPG 2019). With the discovery of new populations in Vietnam, we are currently working to clarify these lineages using an integrated approach that includes reproductive biology, genome size estimation, and nuclear markers. The results of this research will be published in a separate paper.
Lindsaea
kohkongensis
Taxon classificationPlantaePolypodialesLindsaeaceae
I.C.Hwang, M.O.Moon & B.Y.Sun, Korean J. Pl. Taxon. 53(4): 289. 2023.
4A3F447C-5DB1-5B35-8B5F-CB3066F0443B
Type.
Cambodia • Koh Kong, Thma Bang District, near Chamnar village, 22 Dec 2013, Sun et al. C5520 (holotype HIBR; isotypes HIBR, KB).
Distribution and ecology.
This species was recently described from Cambodia and Malaysia. In Vietnam, it grows along a valley in damp tropical forests as lithophytes and is frequently submerged in water.
Specimens examined.
Vietnam. • Kien Giang Province: Phu Quoc district, Phu Quoc National Park, 6 Mar 2018, Chen Wade5034 (TAIF [514046!]).
Note.
Morphologically, this species closely resembles Lindsaeaensifolia. Although L.kohkongensis is usually a much smaller species than L.ensifolia, we did not identify any qualitative trait to distinguish them. Currently, subaquatic habitat preference appears to be the most reliable characteristic for differentiating this species from L.ensifolia. Yun et al. (2023) described L.kohkongensis as having free venation, in contrast to the anastomosing venation found in L.ensifolia. However, this is not true. As demonstrated in Fig. 3G of their paper and Fig. 2F of the current study, the veins of L.kohkongensis are connected near the lamina margin, forming areoles in fertile fronds.
Pteris
pseudowulaiensis
Taxon classificationPlantaePolypodialesPteridaceae
Y.S.Chao, Taiwania 66(3): 314. 2021.
FFBC7C92-BCCB-5197-9143-06B00505EE9C
Type.
Taiwan • New Taipei, Mt. Pataoerh, 600–700 m, 29 Apr 2016, Hsu 8437 (holotype TAIF [497137!]; isotype TAIF [497138!]).
Distribution and ecology.
This species, recently described from Taiwan, was initially reported from southern China and Taiwan (Chao et al. 2021). In Vietnam, P.pseudowulaiensis was discovered in relatively dry evergreen broadleaf forests at lowland areas of Cuc Phuong National Park.
Specimens examined.
Vietnam • Ninh Binh Province: Cuc Phuong National Park, Muong Khu trail, Y.-S. Chao 3509, 3515, 3517, 3531 (TAIF!, VNMN!).
Note.
This species belongs to the Pterisfauriei Hieron. complex, a taxonomically challenging group due to its reticulate evolution involving hybridization, polyploidization, and apomixis (Chao et al. 2022). In Vietnam, five species from this complex have been recorded, namely P.arisanensis Tagawa (Chao et al. 2022), P.kawabatae Sa.Kurata (Chao et al. 2021), P.latipinna Y.S.Chao & W.L.Chiou (Chen et al. 2020b), P.oshimensis Hieron. (Liao et al. 2013), and P.pseudowulaiensis. Morphologically, P.pseudowulaiensis closely resembles P.oshimensis but differs by having broader pinnae, measuring 2–3.5 cm compared to less than 2 cm in P.oshimensis. According to Chao et al. (2022), P.pseudowulaiensis is an apogamous diploid originating from hybridization of P.wulaiensis C.M.Kuo (a species reported from Japan and Taiwan) and another unknown species.
Supplementary Material
XML Treatment for Haplopteris yakushimensis
XML Treatment for Lindsaea kohkongensis
XML Treatment for Pteris pseudowulaiensis
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Barrington DS Paris CA Ranker TA (1986) Systematic inferences from spore and stomate size in the ferns.American Fern Journal 76: 149–159. 10.2307/1547723 · doi ↗
- 2Bidin AA Go R (1995) Chromosome base numbers for Tectaria and allied genera in peninsular Malaysia.Natural History Research (Chiba)3: 115–122.
- 3Chang Y Li J Lu S Schneider H (2013) Species diversity and reticulate evolution in the Aspleniumnormale complex (Aspleniaceae) in China and adjacent areas.Taxon 62: 673–687. 10.12705/624.6 · doi ↗
- 4Chao YS (2019) New record of Pterisesquirolii in Taiwan, with two similar new species, Pterisrugosifolia and Pterissubesquirolii. Systematic Botany 44: 274–281. 10.1600/036364419 X 15562052252072 · doi ↗
- 5Chao YS Dong SY Chiang YC Liu HY Chiou WL (2012) Extreme multiple reticulate origins of the Pteriscadieri complex (Pteridaceae).International Journal of Molecular Sciences 13: 4523–4544. 10.3390/ijms 1304452322605994 PMC 3344230 · doi ↗ · pubmed ↗
- 6Chao YS Ebihara A Chiou WL Huang YM (2017) Pterislatipinna sp. nov. (Pteridaceae), a new species segregated from Pterisfauriei. Phyto Keys 85: 95–108. 10.3897/phytokeys.85.14884 PMC 562421529033663 · doi ↗ · pubmed ↗
- 7Chao YS Chiou WL Ebihara A Hsu TC Chang YH Lin CY (2021) Taxonomic and nomenclatural novelties in the Pterisfauriei group (Pteridaceae).Taiwania 66: 307–316.
- 8Chao YS Ebihara A Chiou WL Tsai JM Huang YW Ranker TA (2022) Reticulate evolution in the Pterisfauriei group (Pteridaceae). Scientific Reports 12: 9145. 10.1038/s 41598-022-11390-7PMC 915998735650209 · doi ↗ · pubmed ↗