Isolation and Genomic Characteristics of a Novel Pathogenicity Type I Feline Coronavirus in Mainland China
Yuanhong Wang, Junna Wang, You Zhao, Yun Liu, Miao Zhang, Xiaoying Deng, Jie Zhu, Guoxin Li, Guangqing Liu

TL;DR
Researchers in China isolated a new pathogenic feline coronavirus strain, HL2019, which causes severe disease in cats and shows genetic similarities to human coronaviruses.
Contribution
The study reports the isolation and genomic characterization of a novel pathogenic FCoV I strain, HL2019, in mainland China.
Findings
HL2019 is a recombinant strain derived from FCoV I China/ZJU1709 and FCoV I Netherlands/UU16.
Two out of three cats infected with HL2019 developed severe symptoms and died within 28 days.
The virus showed high viral load in the duodenum, indicating broad tissue tropism.
Abstract
Feline coronavirus (FCoV) is an enveloped, positive-sense RNA virus, which is widespread among feline populations, and can cause a fatal serious disease called feline infectious peritonitis (FIP). According to the differences of antigen and genetic composition, FCoV consists of two genotypes, FCoV I and FCoV II. In this study, we have isolated and identified a FCoV I strain named HL2019. Based on the complete genome of HL2019, phylogenetic analysis showed that HL2019 strain formed in the cluster FCoV I which is more closed to human coronavirus 229E (HCoV 229E) and HCoV NL63, while the FCoV I stains is distantly related to FCoV II strains. Analyzing with RDP4 and Simplot software showed that the virus HL2019 is recombinant by the FCoV I China/ZJU1709 and FCoV I Netherlands/UU16 strains. Furthermore, the pathogenicity of HL2019 was evaluated in 9–12-month-old cats. Two of three challenged…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
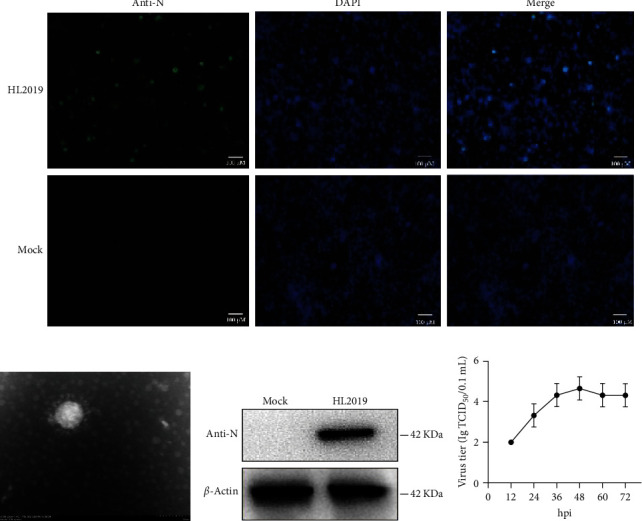 Figure 1
Figure 1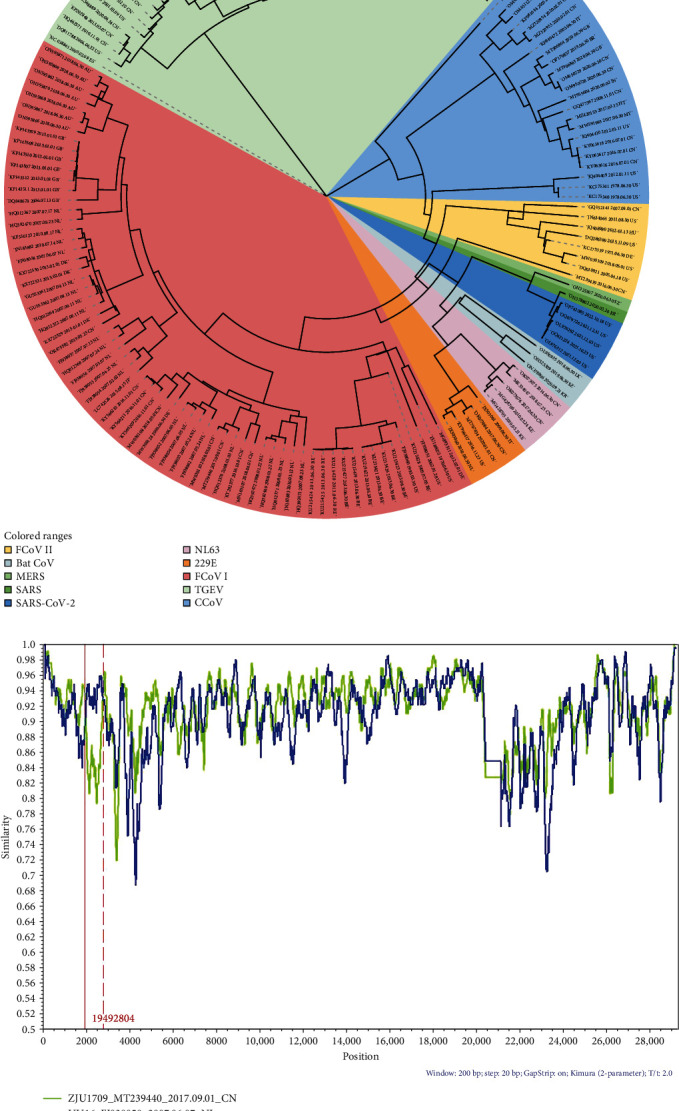 Figure 2
Figure 2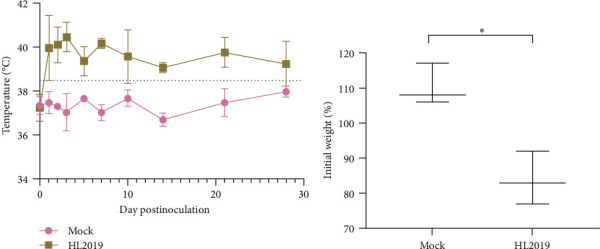 Figure 3
Figure 3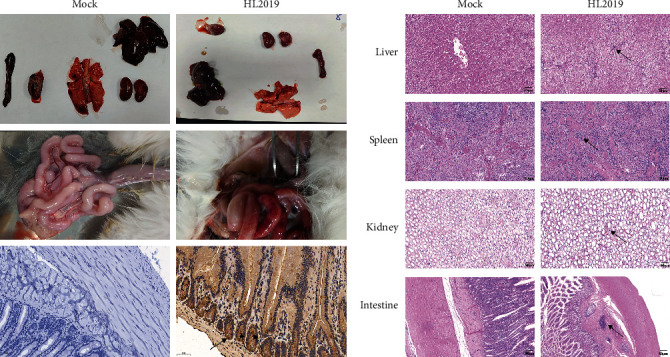 Figure 4
Figure 4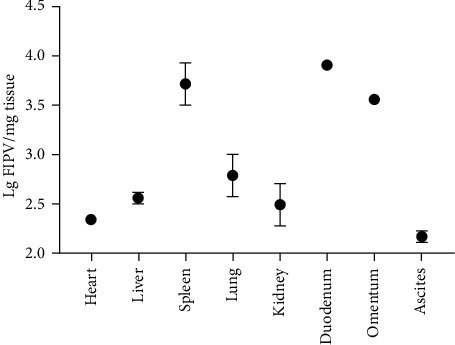 Figure 5
Figure 5- —Science and Technology Commission of Shanghai Municipality
- —National Natural Science Foundation of China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAnimal Virus Infections Studies · Virus-based gene therapy research · Viral gastroenteritis research and epidemiology
1. Introduction
Feline coronavirus (FCoV) is an enveloped, positive-sense RNA virus, which belongs to the genus Alphacoronavirus of the subfamily Orthocoronavirinae, family Coronaviridae [1]. FCoV infection is widespread among feline populations, especially within multicat households and catteries, which can exhibit upward of 96% seropositivity [2, 3]. Based on biological characteristics, FCoV is typically divided into feline enteric coronavirus (FECV) and feline infectious peritonitis virus (FIPV). Most cats with FECV do not show any or only mild symptoms, such as diarrhea and anorexia [4]. But FIPV could cause a more serious disease called feline infectious peritonitis (FIP), which results in fatal disease in cats once clinical signs develop [5].
As companion animal, people are increasingly concerned about the physical health of cats. Previous studies have shown that severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) can replicate in the cats' nose and throat and caused inflammatory pathology deeper in the respiratory tract, and airborne transmission did occur between pairs of cats [6]. In addition, a Malaysian study has identified a novel canine–feline recombinant alphacoronavirus named CCoV-HuPn-2018 which can infect human [7]. This suggests the possibility of cross-species transmission of CoV between human and feline. Tracking the origin of emerging pathogens is crucial for controlling their spread, and research on the transmission of pathogens with different species helps prevent pandemics [8].Therefore, it is necessary to figure out the epidemiology and biological characteristics of FCoV.
In this study, we first isolated a FCoV I strain named HL2019 in mainland China. We provide molecular characterization and pathogenicity of the virus HL2019. The results showed that this virus was highly pathogenic in cats and replicated efficiently in the duodenum. And this study will be helpful for understanding the evolution and pathogenicity of FCoV in mainland China.
2. Materials and Methods
2.1. Ethics Statements
We performed the animal experiment following the guidelines of the Institute of Shanghai Veterinary Research, Chinese Academy Agriculture of Sciences, using protocols approved by the Institutional Laboratory Animal Care and Use Committee (SV-20230728-01).
2.2. Sample Collection
Clinical samples (cat spleen and ascites) were collected from the British shorthair cat in Anhui province, China, in 2019. The cat showed a typical FIP signalment, including effusion, hyperproteinemia, hyperglobulinemia, hypoalbuminemia, and hyperbilirubinemia.
2.3. Isolation and Identification of HL2019 Strain
To isolate FCoV, we aseptically treated the effusion and inoculated it into a monolayer of FCWF-4 cells (American Type Culture Collection (ATCC): B184268), which were growing in Eagle's Minimum Essential Medium (EMEM; Hyclone, USA) supplemented with 5 μg/mL 0.25% (w/v) trypsin (Gibco, Australia) in 5% CO_2_ at 37°C. After ~80%–90% of the cells exhibited cytopathic effect (CPE), we conducted repeated freezing and thawing, and then we eliminated cell debris by centrifugation at 1500 × g for 5 min at room temperature. The supernatant containing the mature viral particles was stained with 2% phosphotungstic acid and observed with electron microscope (EM).
2.4. Western Blot and Immunofluorescence Assays
After 24 h postinfection, we fixed FCWF-4 cells infected with the virus HL2019 in 6-well plates at a one multiplicity of infection (MOI) with 4% paraformaldehyde. Subsequently, we used the FCoV I N-protein polyclonal antibody (prepared in our laboratory) as the primary antibody (1:100), followed by fluorescent isothiocyanate (FITC)-labeled goat antimouse secondary antibody (1:10,000) (Beijing, ZSBio) to examine fluorescence. We counterstained with 4′,6-diamidino-2-phenylindole (DAPI) at room temperature for 5 min and used a fluorescence microscope (Nikon, Tokyo, Japan) for examination. And we also performed blank FCWF-4 cells in immunofluorescence assay (IFA) as a negative control. In addition, the primary antibody FCoV I N-protein polyclonal antibody [9] and the secondary antibody horseradish peroxidase (HRP)-conjugated goat antimouse immunoglobulin G (IgG; Beijing, ZSBio; 1:5000) were subjected to the western blot assay.
2.5. Next-Generation Sequencing (NGS)
The HL2019 P3 was harvested for the extraction of viral RNA. Illumina sequencing and library construction were performed at the Shanghai Tanpu Biotechnology Co., Ltd (Shanghai, China). Specifically, the NEBNext Ultra II RNA Library Prep Kit (NEB, Ipswich, MA, USA) was used for library construction. After adapter ligation, 10 cycles of polymerase chain reaction (PCR) amplification were used to enrich the sequencing target. The libraries were pooled in equimolar amounts, denatured, and diluted to the optimal concentration before sequencing. The Illumina NovaSeq 6000 System (Illumina, San Diego, CA, USA) was used for sequencing to generate 150-bp pair-end reads.
2.6. Sequence Datasets
The sequences we have used in this study were obtained from the NCBI GenBank database (https://www.ncbi.nlm.nih.gov/), and 73 FCoV strains were downloaded (accessed on June 31, 2023). Specifically, 24 strains of canine coronavirus (CCoV), 46 strains of transmissible gastroenteritis virus (TGEV), 5 strains of human coronavirus (HCoV) NL63, 5 strains of HCoV 229E, and 5 strains of bat alphacoronavirus (BatCoV) were used to understand the relationship between Alphacoronavirus. To be specific, we downloaded five strains of SARS-CoV-2 which are from Betacoronavirus (Table 1).
2.7. Phylogenetic Analysis
For comparing and sorting the complete genome, ModelFinder software was used to select best model which is generalized time-reversible (GTR)+F+G4 and assume an uncorrected relaxation clock (lognormal) [10, 11]. The chain length for each calculation is set to 5 × 10^9^ generations, obtained every 50,000 generations, with a total of three independent operation sets. After burn-in (10%), the Tracer software (V1.7.1) was used to estimate the data. Parameters with an effective sampling size of more than 200 were accepted [12]. FigTree software (V1.4.4) was using to display the final maximum clade credibility (MCC) tree.
2.8. Recombination Screening
For understanding the potential recombination events, the occurrence of potential recombination events between complete genome sequences of 163 strains in this study was analyzed using seven algorithms of the RDP4 software package, including RDP, GENECONV, Bootscan, MaxChi, Chimaera, SiScan, and 3Seq [13]. The recombination event identified by at least four of the seven methods was accepted in this report; the recombination events were confirmed by at least four methods with a p-value cutoff of 0.05. Furthermore, the SimPlot 3.5.1 software was used to confirm the potential recombinant events occurred in HL2019 by using the neighbor-joining method in the Kimura 2-parameter (K2P) model [14]. The window size and step were set to 200 bp and 20 bp, respectively, with a p-value cutoff of 0.05.
2.9. Animal Experiments
In this study, for the cats used in this experiment, we have used reverse transcription PCR (RT-PCR) to confirm feline calicivirus (FCV) and FCoV were pathogen-free and PCR to confirm feline panleukopenia virus (FPV) and feline herpesvirus (FHV) were pathogen-free (Table S1). Then the six 9–12-month-old cats were randomly divided into two groups: HL2019 group (n = 3) and Mock group (n = 3). All animals were housed in the P2 laboratory. Cats in HL2019 group were inoculated orally with isolated virus (10^5^ TCID_50_/cat), while cats in the Mock group were inoculated orally with phosphate-buffered saline (PBS). According to Thayer et al. [15] which have published the Diagnosis Guidelines in 2022, the clinical signs and symptoms were monitored daily. Two of the cats inoculated with the virus died after 3 weeks. The copy numbers of the heart, liver, spleen, lung, kidney, duodenum, and ascites were obtained by N EvaGreen real-time RT-PCR [5]. The duodenum was also analyzed for hematoxylin and eosin (H&E) and immunohistochemistry (IHC) stain.
3. Results
3.1. Sequencing and Virus Isolation
After 72 h, FCWF-4 cells exhibited cytopathic effects, including shrinking, rounding, lighting, and disruptive morphological characteristics (Figure 1A). And the typical crown particle was observed under transmission electron microscopy (TEM) with a diameter of about 100 nm (Figure 1B). Meanwhile, western blot analysis results indicated that the isolated virus (named HL2019) could specifically react with FCoV I N-protein polyclonal antibody (Figure 1C). Multistep replication curves revealed that the mean virus titer of HL2019 was the highest at 48 hpi (Figure 1D).
Furthermore, we have sequenced the complete genome of HL2019 P3; the results showed that the complete nucleotide sequence of HL2019 consisted of 29,044 nt (excluding the poly [A] tail) and contained 11 open reading frames (ORFs), which encoded four structural proteins (S, E, M, and N), five accessory proteins (7a and 7b, 3a, 3b, and 3c), and two nonstructural proteins (1a and 1b). The poly (A) tail of FIPV had 20 A's at least on either side of the coding region of the genome are 5′ untranslated region (5′UTR) and 3′UTR, which are 294 nt and 247 nt long, respectively.
Our data confirmed that we have successfully isolated a FCoV I HL2019 strain. The genomic sequence of HL2019 has been submitted to GenBank, and its accession number is OR475582.
3.2. Potential Recombination Events in HL2019 Strain
In this study, RDP4 was used to detect recombination and confirmed that there has recombinational event that existed among strains; we found that MT239440 (FCoV I ZJU1709) and FJ938058 (FCoV I UU16) might represent the parent lineages of OR475582 (FCoV I HL2019). Similarity plot and bootscanning analyses predicted the potential breakpoints in HL2019 to be at ORF1a(nt 2021 and 2821) (Figure 2A). ORF1a encodes a papain-like protease (PLpro) which is essential for coronaviral replication [16]. The occurrence of recombination and mutation events in ORF1a region may be a crucial strategy employed by FCoV to evade innate and adaptive immunity.
3.3. Relationship Among Multiple Species of Coronavirus
In order to analyze the phylogenetic relationship between different species of coronavirus, we download some alphacoronavirus complete genomes for reconstructing MCC (Figure 2B). The results of the phylogenetic trees showed the HL2019 strains located in the clade of FCoV I and closed to the UG-FH8 which is isolated from Denmark. The clade of FCoV I stains was distantly related to clade of FCoV II strains, while the clade of FCoV II strains was closed to TGEV and CCoV strains. Interestingly, we found that the clade of FCoV I showed high confidence with HCoV NL63 and HCoV 229E. Further analyzed on the homology of HCoV NL63 (MG428705.1), HCoV 229E (JX503060.1), and FCoV I HL2019 (OR475582), it is showed that the homology of HCoV NL63 (MG428705.1) and HL2019 (OR475582) is up to 60.1%, and the homology of HCoV 229E (JX503060.1) and HL2019 (OR475582) reached to 61.2%. It indicated that the FCoV may have cross-species transmission potential.
3.4. HL2019 Is Highly Pathogenic to Cats
Prior to the challenge, all cats were healthy. After oral challenge with HL2019 (10^5^ TCID_50_/cat), two of the three cats showed typical sign associated with FIP including inappetence, anorexia, weight loss, fever, and diarrhea and died within 28-day postchallenge (dpc). Specifically, cats in the HL2019 group developed a persistent fever after virus challenge, while cats in the Mock group maintained normal (Figure 3A). Additionally, the average weight of cats in the HL2019 group (84% of initial weight) was significantly lower than the Mock group (110% of initial weight) (Figure 3B). On 28 dpc, all cats were euthanized. In the HL2019 group, the ascites were observed, with an intestinal bleeding and congestion (1/3), while the Mock group remain normal, and the IHC results further confirmed that viral antigen was detected within macrophages in the lesions of cats from the HL2019 groups (Figure 4A).
For H&E examination, distinct focal accumulations of macrophages were observed just under the capsule of the liver, spleen, kidney, and intestine in the HL2019 groups, while the Mock groups remain normal (Figure 4B).
3.5. Detection of Virus Load
FCoV antigens were detected in organs, such as the lung, heart, spleen, liver, kidney, duodenum, omentum, and ascites. And it is abundant in the duodenum and spleen which is reaching 10^4^ copies/mg (Figure 5).
4. Discussion
FCoV processes about 29 k-nucleotide positive-sense single-stranded genomic RNA, which is composed of 11 ORFs encoding 7 nonstructural proteins (1a, 1b, 3a, 3b, 3c, 7a, and 7b) and 4 structural proteins (the spike, envelope, membrane, and nucleocapsid proteins) [2, 3]. According the spike sequence, FCoV has divided into two kinds of serum type (FCoV I and FCoV II) [17, 18]. FCoV I accounts for the bulk (80%–90%) of natural infections in cats, while FCoV II (naturally occurring recombination between FCoV I and CCoV spike proteins) is far less prevalent (<10%) [19]. FCoV I is difficult to be cultured in vitro, while FCoV II is easier to grow in cell cultures. Previous studies have reported FCoV I to be isolated, but the titer has not been defined [20, 21]. In our study, we have successfully isolated a FCoV I strain on FCWF-4 cells, and its titer is reaching 10^5^ TCID_50_ which is important to study the pathogenic mechanism of FCoV I and development of related biological products against FCoV.
Recombinant analysis was performed to examine the evolution of the isolate HL2019 strain. The HL2019 is located in cluster FCoV I and most closely related to UG-FH8 which was isolated in Denmark. In addition, recombination analysis indicated that a potential recombination event had occurred within ORF1a. The primary function of PLpro is to process the viral polyprotein in a coordinated manner. PLpro has the additional function of stripping ubiquitin and ISG15 from the host cell proteins to aid coronaviruses in their evasion of the host innate immune responses [22, 23]. Of note, increasing studies point to that cats can be the intermediate hosts for some HCoVs, such as SARS-CoV-2 and HCoV-229E [24, 25]. In addition, feline-like CoV strains have been reported to infect humans [26], suggesting that FCoV has the potential for interspecies transmission or evolve as a new human strain. In this study, we have found that the FCoV I strains are more closed to HCoV, including HCoV 229E and HCoV NL63, suggesting cats play a potential role in interspecies transmission of coronaviruses. So it is necessary to conduct more in-depth research to dig out the viral variability and evolution of FCoV.
Furthermore, our results showed that this virus was pathogenic in 9–12-month-old cats. Meanwhile, real-time PCR analysis showed that HL2019 has broad tissue tropism; FCoV was abundant in the duodenum which is up to 10^4^ copies/mg. In addition, the levels of viral genomic RNA paralleled the intensity of the inflammatory response, which in turn paralleled the numbers of infected macrophages in the tissues or effusion. Accoring to Kiss, Poland, and Pedersen [27] study, they used an avirulent type I strain of FIPV-UCD1 and challenge exposed to a highly virulent cat passaged type I strain FIPV-UCD8. They found a FIPV-UCD1 immunization induced only partial protection at best, as gauged against historical data. Animal-passaged FIPV-UCD8 usually kills from 90 to 100% of inoculated cats, almost always from effusive FIP. And in Terada et al. [28] study, they discovered C3663, a strain of FCoV I isolated from FIP cats that retained virulence despite adaptation in FCWF-4 cells. It could make three (75%) of four SPF cats developed FIP after infection with the C3663 strain, and in their further research, they found the cDNA clone for type I FCoV strain C3663. It seems the virulence of FCoV I strains varies greatly, and the specific mechanism still needs further study.
5. Conclusion
In a word, this is the first study to report the complete viral genome of FCoV I strain HL2019 isolated from FCWF-4 cells in the mainland of China. We provide evidence of the pathogenicity of the virus HL2019 which provided scientific evidence that HL2019 could be highly pathogenic to cats. These results provide important information on the evolution of FCoV in mainland of China and suggest that we need to continue surveillance studies in felines to monitor the spread and evolution patterns of FCoV and other CoV infections, as well as the detection of recombinant viruses.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Doki T. Yabe M. Takano T. Hohdatsu T. Differential Induction of Type I Interferon by Type I and Type II Feline Coronaviruses in Vitro Research in Veterinary Science 2018120576210.1016/j.rvsc.2018.09.0022-s 2.0-8505380922230265871 PMC 7111755 · doi ↗ · pubmed ↗
- 2de Cássio V. de Barros B. de Castro C. M. O. Pereira D. First Complete Genome Sequence of a Feline Alphacoronavirus 1 Strain from Brazil Microbiology Resource Announcements 20198101535151810.1128/MRA.01535-182-s 2.0-85064667236 PMC 640611430863824 · doi ↗ · pubmed ↗
- 3Neuman B. W. Buchmeier M. J. Advances in Virus Research Advances in Virus Research 20169612710.1016/bs.aivir.2016.08.0052-s 2.0-8499471614927712621 PMC 7112365 · doi ↗ · pubmed ↗
- 4Cook S. Castillo D. Williams S. Haake C. Murphy B. Serotype I and II Feline Coronavirus Replication and Gene Expression Patterns of Feline Cells—Building a Better Understanding of Serotype I FIPV Biology Viruses 202214710.3390/v 140713561356 PMC 932044735891338 · doi ↗ · pubmed ↗
- 5Wang G. Hu G. Liang R. Establishment of Full-Length c DNA Clones and an Efficient Oral Infection Model for Feline Coronavirus in Cats Journal of Virology 2021952110.1128/JVI.00745-21e 0074521 PMC 851346234406859 · doi ↗ · pubmed ↗
- 6Shi J. Wen Z. Zhong G. Susceptibility of Ferrets, Cats, Dogs, and Other Domesticated Animals to SARS-Coronavirus 2 Science 202036864941016102010.1126/science.abb 701532269068 PMC 7164390 · doi ↗ · pubmed ↗
- 7Tortorici M. A. Walls A. C. Joshi A. Structure, Receptor Recognition, and Antigenicity of the Human Coronavirus C Co V-Hu Pn-2018 Spike Glycoprotein Cell 20221851322792291.e 1710.1016/j.cell.2022.05.01935700730 PMC 9135795 · doi ↗ · pubmed ↗
- 8Cheng H.-Y. Jian S.-W. Liu D.-P. Contact Tracing Assessment of COVID-19 Transmission Dynamics in Taiwan and Risk at Different Exposure Periods Before and After Symptom Onset JAMA Internal Medicine 202018091156116310.1001/jamainternmed.2020.202032356867 PMC 7195694 · doi ↗ · pubmed ↗