Role of the TPR family protein VPA1365 in regulating type III secretion system 2 and virulence in Vibrio parahaemolyticus
Wenliang Yin, Mengyan Wan, Youkun Zhang, Hongmei Meng, Zhiming Pan, Xinan Jiao, Dan Gu

TL;DR
A new protein, VPA1365, was found to regulate a key system in Vibrio parahaemolyticus that helps it cause disease in the gut.
Contribution
VPA1365, a TPR family protein, is a novel regulator of T3SS2 and virulence in Vibrio parahaemolyticus.
Findings
VPA1365 positively regulates T3SS2 and its associated virulence genes.
Deletion of VPA1365 reduces cytotoxicity, adhesion, biofilm formation, and hemolytic activity.
VPA1365 binds to promoters of vtrA, scrG, pilA, and mshA to control virulence.
Abstract
Vibrio parahaemolyticus is a notable seafood-borne pathogen capable of colonizing the intestines of hosts and inducing acute gastroenteritis. The intestinal colonization and enterotoxicity of V. parahaemolyticus are highly reliant on the type III secretion system 2 (T3SS2), encoded within the pathogenicity island (Vp-PAI). The expression of Vp-PAI is strictly regulated by bile acid signals and transcriptional regulators VtrA/VtrB. In this study, we identified a tetratricopeptide repeat (TPR) family protein named VPA1365, which regulates the expression of T3SS2 and is indispensable for the intestinal colonization of V. parahaemolyticus. The expression and secretion of the T3SS2-dependent protein VopD2 were significantly reduced in Δvpa1365 compared to that of the wild type (WT), suggesting that VPA1365 positively regulates the function of T3SS2. Further research indicated that VPA1365…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
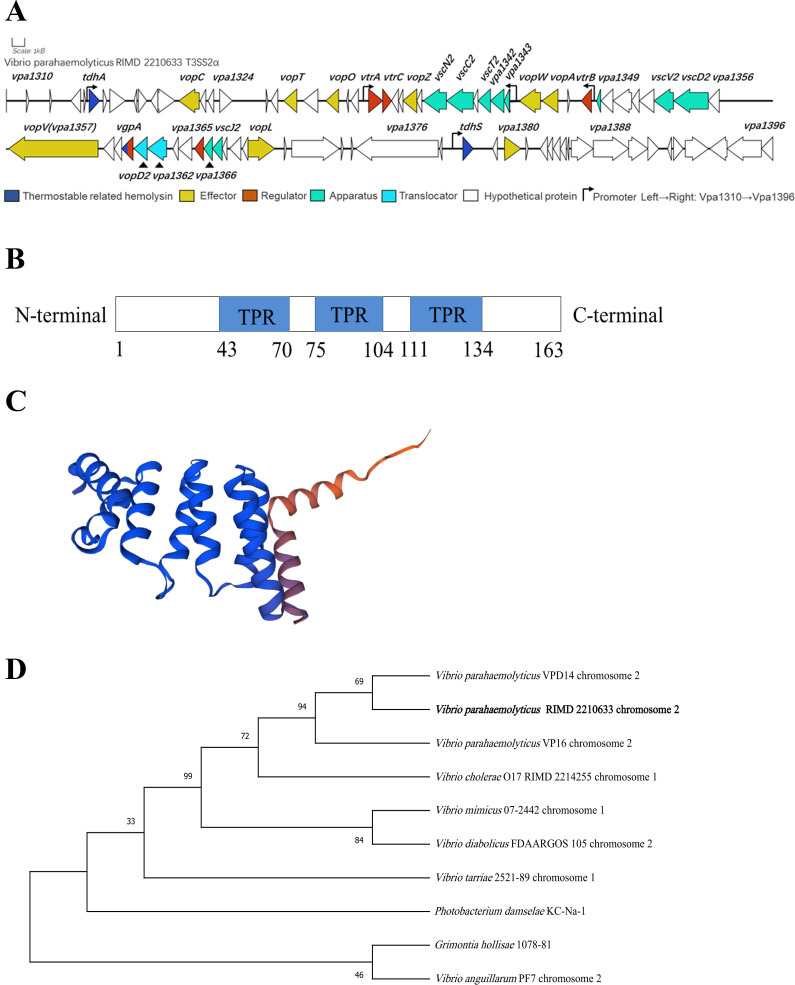 Fig 1
Fig 1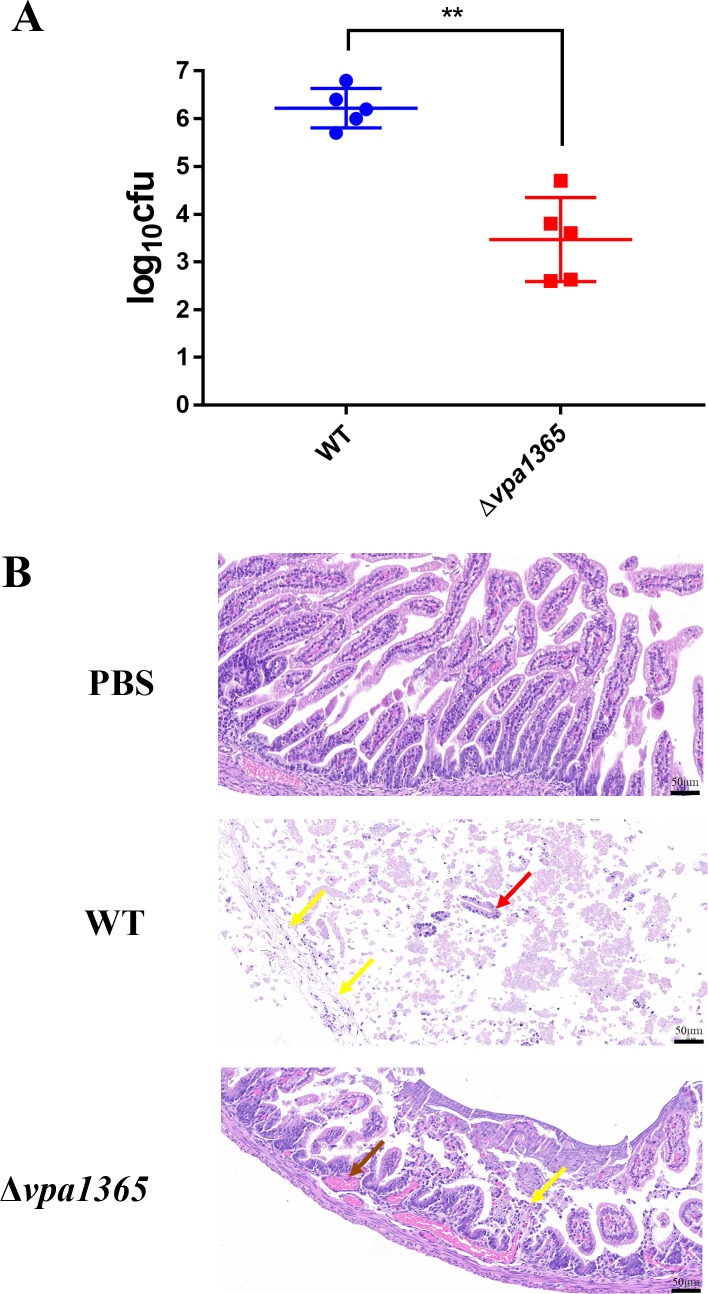 Fig 2
Fig 2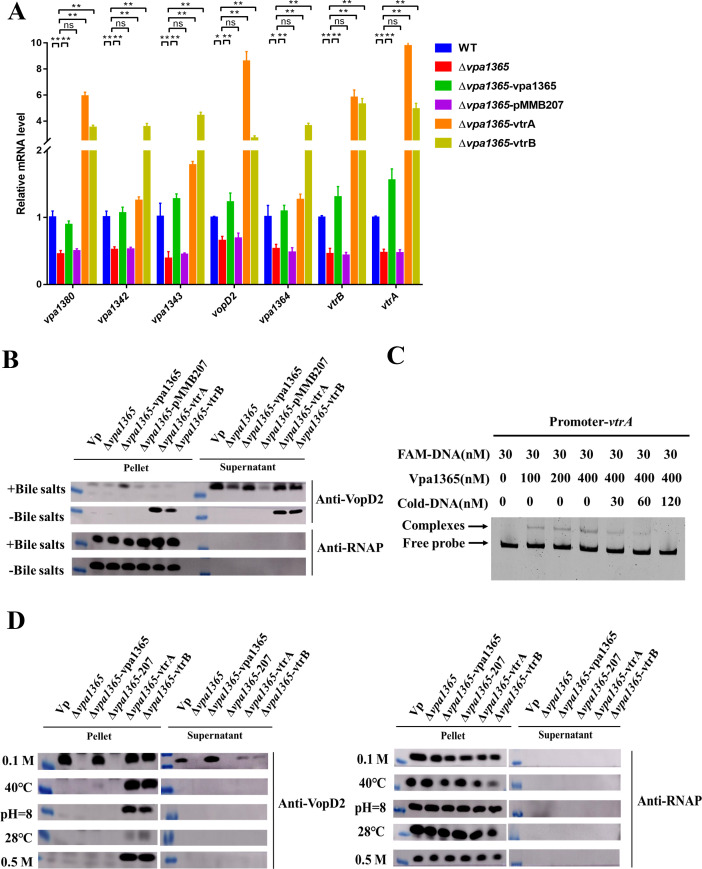 Fig 3
Fig 3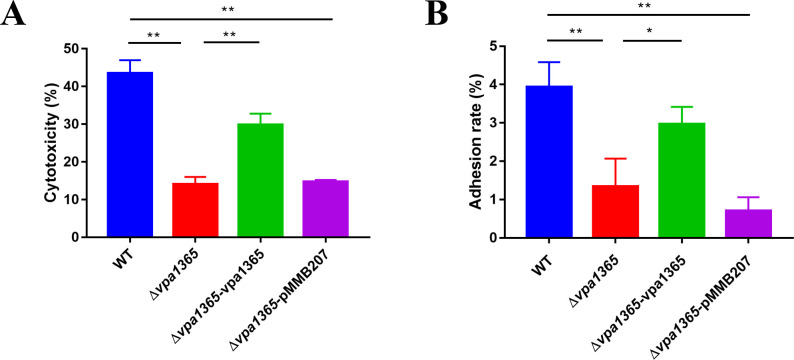 Fig 4
Fig 4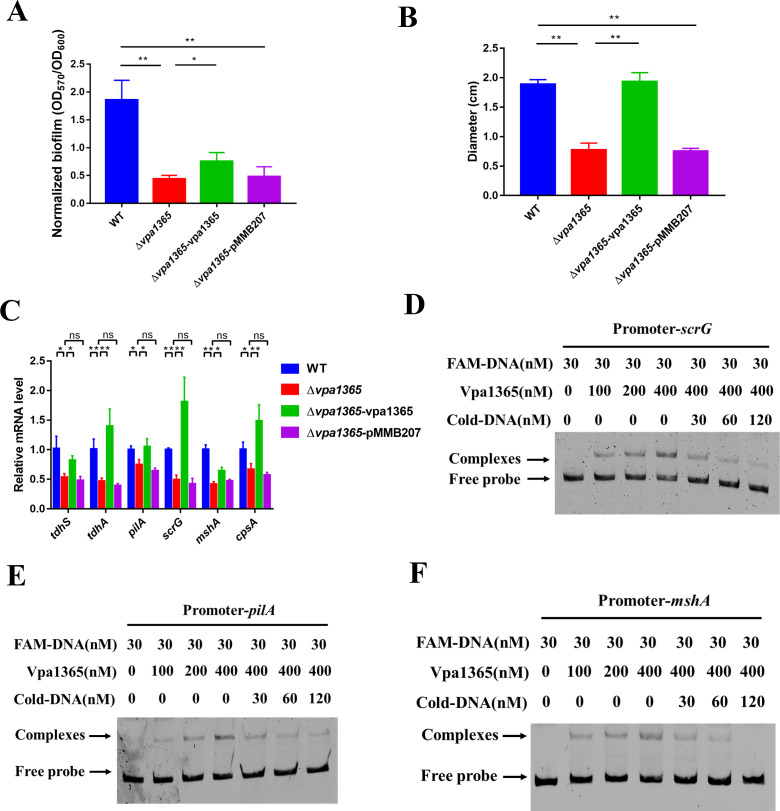 Fig 5
Fig 5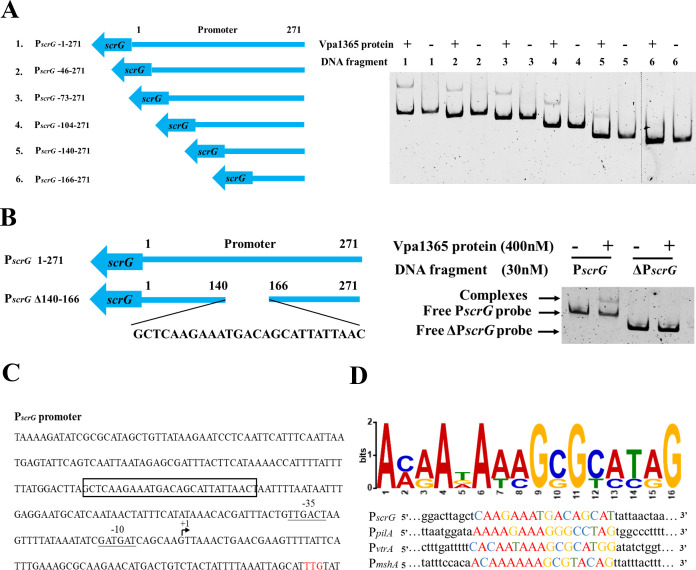 Fig 6
Fig 6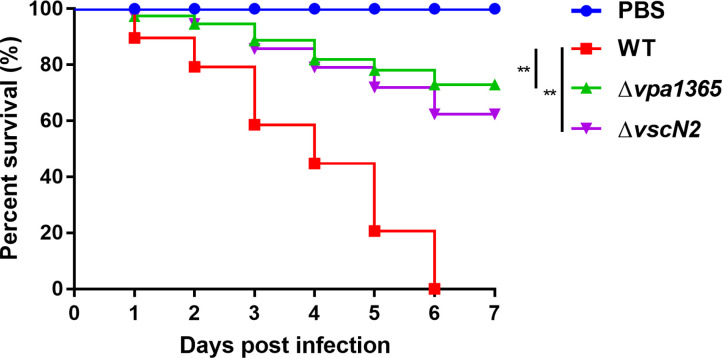 Fig 7
Fig 7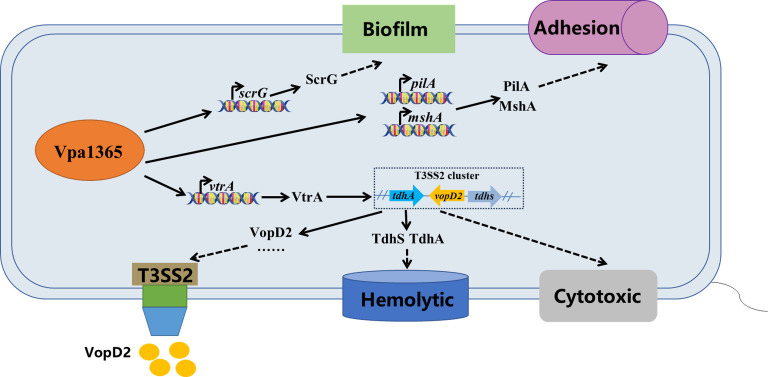 Fig 8
Fig 8| Strains/plasmids | Description | Reference/source |
|---|---|---|
| DH5α | Host for π requiring plasmids | ( |
| SM10 | Host for π requiring plasmids, conjugal donor | ( |
| BL21(DE3) | Host strain for protein expression | Lab collection |
| BL21(DE3)- | BL21, pCold1 carrying | This study |
| RIMD2210633 | WT, O3:K6 strain clinical isolate, CarbR | ( |
| Δ | RIMD2210633, in-frame deletion in | This study |
| Δ | RIMD2210633, in-frame deletion in | This study |
| Δ | Δ | This study |
| Δ | Δ | This study |
| Δ | Δ | This study |
| Δ | Δ | This study |
| Plasmids | ||
| pDM4 | CmR, | ( |
| pMMB207 | CmR, IPTG-induced expressing vector | ( |
| pCold1 | CmR, IPTG-induced expressing vector | Lab collection |
| pDM4- | CmR, pDM4 containing homologous arms of | This study |
| pMMB207- | CmR, | This study |
| pMMB207- | CmR, | This study |
| pMMB207- | CmR, | This study |
| pCold1- | CmR, | This study |
- —National Natural Science Foundation of Chinahttp://dx.doi.org/10.13039/501100001809
- —Postgraduate Research & Practice Innovation Program of Jiangsu Province
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsVibrio bacteria research studies · Child Nutrition and Water Access · Aquaculture disease management and microbiota
INTRODUCTION
Vibrio parahaemolyticus, widely distributed in the ocean, estuaries, and even freshwater (1), is the causal agent of severe vibriosis in aquatic economic animals (2). V. parahaemolyticus infections may lead to diverse life-threatening human diseases, including acute diarrhea and sepsis (3, 4). The pathogenicity of V. parahaemolyticus is related to various virulence factors, including the type III secretion system (T3SS1 and T3SS2), thermostable direct hemolysin (TDH), TDH-related hemolysin, type VI secretion system, motility, and biofilm formation (5, 6). During infections, bacteria usually adhere to the surface of the host cell through flagella or pili and then quickly adapt to the host environment by producing polysaccharides to form biofilms, secreting hemolysin, and T3SS effector proteins to evade the host immune system and acquire nutrients (7–9). Interestingly, the fecal samples from 1,900 patients in a Shanghai hospital indicated that the T3SS2, encoded on chromosome 2, was present in all V. parahaemolyticus clinical isolates (10). Most strains isolated from the environment only contain the T3SS1 and lack the T3SS2. This finding suggests that T3SS2 may be one of the critical virulence factors in foodborne diseases caused by V. parahaemolyticus (11). Thus, it is urgent to understand the pathogenesis mediated by the T3SS2, especially the regulatory mechanisms and biological functions of many hypothetical T3SS2 genes, which is essential for identifying anti-virulence targets and controlling the virulence of the opportunistic pathogen V. parahaemolyticus in the future.
The type III secretion system, extensively present in many Gram-negative pathogens, is the needle-like apparatus that injects many effector proteins into host cells to hijack host signaling pathways and quickly cause disease (12–14). The pandemic serotype O3:K6 V. parahaemolyticus strain RIMD2210633 generally carries two types of T3SSs (T3SS1 and T3SS2). T3SS1 is regarded as being tightly associated with cytotoxicity, whereas T3SS2 is responsible for bacterial colonization ability and enterotoxicity in vivo (15–17). Several functional analyses have shown that the T3SS2 encoded in an 80 kb Vp-PAI plays the major pathogenic factor in tissue invasion and gastroenteritis involved in many animal models (18–20). Various T3SS2 effectors in V. parahaemolyticus modifying normal cell functions have been identified, including VopA, VopC, VopG, VopL, VopO, VopT, VopV, VopY, VopZ, and VPA1380 (6, 21–23), which is constantly expanding our knowledge of the enteropathogenicity mechanism of T3SS2 in this bacterium.
The expression of T3SS1 and T3SS2 in V. parahaemolyticus is strictly regulated by various environmental signals and transcriptional factors. The AraC/XylR-type transcription factor ExsA regulates the expression of T3SS1 by directly binding to the conserved sequence (5′-TNAAANA-3′) in the promoter region of T3SS1 under the conditions of Dulbecco’s modified Eagle’s medium (DMEM) with low calcium levels (24). Bile salts act as an environmental signal by interacting with VtrA/C, which then VtrA/C activates the expression of the OmpR family transcription factor VtrB. VtrB directly binds to the promoter region of T3SS2 and induces its expression (25, 26). Additionally, the LysR-type transcription factor CalR and transmembrane regulatory protein ToxR could also bind to the promoter region of the gene cluster to directly control T3SS2 transcription (27). Recent studies have shown the essential function of the uncharacterized proteins located in the T3SS2 gene cluster and other environmental factors affecting the function of T3SS2. For example, the gatekeeper protein VgpA (VPA1360) was found to promote the secretion of T3SS2 substrates by interacting with the ATPase VscN2 (28), and a new T3SS2 effector VopY (VPA1324) was proved to suppress host STING-dependent signaling by hydrolyzing c-di-GMP (29). Our previous study discovered that low salinity (0.5% NaCl) induces the secretion of the T3SS2 protein VPA1361, which indicates that low salinity may be a neglected environmental signal regulating virulence of V. parahaemolyticus (30). Unfortunately, the regulation patterns of T3SS2 have not been fully elucidated, and the biological functions of several hypothetical proteins in the T3SS2 gene cluster (vpa1310 to vpa1396) require further intensive exploration.
Twenty-two regulators of 230 genes were identified to be involved in the small intestine colonization of V. parahaemolyticus by transposon insertion sequencing (Tn-seq) technology (27), including the uncharacterized gene vpa1365 located on the T3SS2 gene cluster. In this study, we first confirmed that the vpa1365 indeed contributed to the intestinal colonization of V. parahaemolyticus. Moreover, we further investigated the roles of VPA1365 in regulating T3SS2 and the virulence of V. parahaemolyticus. Our results indicated VPA1365 positively regulates the expression of numerous genes by binding to the promoters of T3SS2-related and virulence genes, thereby influencing the expression and secretion of T3SS2, hemolytic activity, biofilm formation, adhesion, and cytotoxicity. These findings deepen our understanding of the essential role of VPA1365 in regulating T3SS2 and virulence in V. parahaemolyticus.
MATERIALS AND METHODS
Bacterial strains, plasmids, and growth conditions
All strains and plasmids utilized in this study were listed and described in Table 1. Escherichia coli, V. parahaemolyticus RIMD2210633, and its derivative mutants were all grown at 37°C in the Luria-Bertani (LB) medium. The following concentrations of antibiotics were used when necessary: 100 µg/mL carbenicillin (Carb) and 24 µg/mL chloramphenicol (Cm). Furthermore, 1 mM isopropyl β-D-1-thiogalactopyranoside (IPTG) was used to induce the expression and secretion of VPA1365 in the Δ*vpa1365-vpa1365 and Δvpa1365-*pMMB207 strains. The bile salts (0.04%, [wt/vol]) were used to stimulate the expression and secretion of VopD2 in the wild type (WT), ∆vpa1365, and complemented strains.
Construction of deletion mutants and complemented strains
The vpa1365 and vscN2 (vpa1338) in-frame deletion mutants of V. parahaemolyticus were generated according to a previous method (36), and the primers were tabulated in Table S1. Briefly, the upstream and downstream fragments of vpa1365, amplified from the RIMD2210633 genome DNA with primers vpa1365 up-F/R and down-F/R, were cloned into the SacI/SalI sites of the suicide vector pDM4 using a pEASY-Basic seamless cloning kit (Transgen, Beijing, China). The positive recombinant plasmid pDM4-vpa1365 was transformed into SM10 λpir by heat shock, and then the recombinant plasmid was transferred into the RIMD2210633 strain through conjugation. After that, the double-crossover recombination was selected to produce the in-frame deletion mutant Δvpa1365 in the LB agar plate with 10% sucrose. All mutants were confirmed by PCR with the primers vpa1365 out-F/R and sequenced for reaffirmation. The deletion of the vscN2 gene in V. parahaemolyticus is similar to the above method.
The open reading frame (ORF) of vpa1365 was amplified with primers vpa1365-207-F/R and cloned into the XbaI/HindIII sites of the pMMB207 plasmid (with the chloramphenicol resistance gene). The recombinant plasmids pMMB207::vpa1365, pMMB207::vtrA, and pMMB207::vtrB were transformed into the deletion strain Δvpa1365 to generate the complemented strains named Δvpa1365-vpa1365, Δvpa1365-vtrA, and Δvpa1365-vtrB, respectively. The pMMB207 plasmid was transformed into the Δvpa1365 to generate the vector control strain named Δ*vpa1365-*pMMB207. All complemented strains were confirmed by PCR with the primers pMMB207-F/R and sequenced.
Sequence analysis
The National Center for Biotechnology Information accession numbers (http://www.ncbi.nlm.nih.gov/genbank) for VPA1365 and homologous fragments are V. parahaemolyticus VPD14 (CP031782.1), V. parahaemolyticus VP16 (CP127847.1), Vibrio cholerae O17 (AP023371.1), Vibrio tarriae 2,521–89 (CP022353.1), Vibrio mimicus 07–2,442 (CP046825.1), Vibrio diabolicus (CP014037.1), Vibrio anguillarum (CP011465.1), Photobacterium damselae KC-Na-1 (CP021153.1), and Grimontia hollisae 1,078–81 (CP046810.1), respectively. The InterPro tool was used to identify the conserved domains of VPA1365 (https://www.ebi.ac.uk/interpro/), and the 3D protein structure prediction was conducted by SWISS-MODEL (https://swissmodel.expasy.org/). A bootstrap consensus tree was constructed using the neighbor-joining method from the MEGA-X program (37).
Cytotoxicity and adhesion assay
The lactate dehydrogenase (LDH) released from infected HeLa cells was measured as described previously (38). Overnight cultured V. parahaemolyticus strains were adjusted to an OD_600_ of 1, diluted 1:100 into LB medium, and cultured at 37°C with shaking for 4 h. Subsequently, the culture was centrifuged, resuspended, and diluted to a concentration of 10^8^ CFU/mL in DMEM containing 10% fetal bovine serum. HeLa cells were infected by WT, Δvpa1365, Δvpa1365-vpa1365, and Δvpa1365-pMMB207 at a multiplicity of infection of 1:100. After infection at 37°C for 2 h, the LDH cytotoxicity assay was examined and assessed according to the manufacturer’s protocol (Invitrogen, USA). Meanwhile, the infected HeLa cells were lysed with 200 µL of 0.01% Triton-X 100 after washing once with Dulbecco’s phosphate-buffered saline (PBS). The 10-fold serial dilution lysates (100 µL) were respectively plated on thiosulfate citrate bile salt sucrose agar plates culturing at 37°C for 18 h. The adhesion rate was calculated as the number of bacterial cells adhered per the input total number of bacterial cells.
Hemolytic activity assay
Overnight cultures of WT, Δvpa1365, Δvpa1365-vpa1365, and Δvpa1365-pMMB207 were centrifuged, washed twice with PBS, resuspended in PBS at a final concentration of 1 × 10^9^ CFU/mL, and then spotted 10 µL on the Wagatsuma blood agar base for 37 h. The measured circle diameter of the clear zone was considered as the hemolytic activity (39, 40). Three separate experiments were performed in biological triplicates each.
Biofilm formation assay
Overnight cultures of WT, Δvpa1365, Δvpa1365-vpa1365, and Δvpa1365-pMMB207 were adjusted to an OD_600_ of 1, diluted 1:100 into 200 µL of fresh LB medium in the 96-well polystyrene microplate, and cultured statically at 37°C for 31 h. The bacterial cells were removed and washed twice with PBS, and 220 µL of 0.1% crystal violet was added to stain biofilm for 30 min. Afterward, 200 µL of 95% ethanol was added to dissolve the crystal violet for 1 h after removing residual stains, and the amount of biofilm was determined at 570 nm using a microplate reader (Epoch 2, BioTeK). The biofilm formation was normalized as the total biofilm per cell density of bacteria. Three separate experiments were performed with biological triplicates each.
Immunoblotting analysis
For VopD2 immunoblotting analysis was carried out as previously reported (30), the overnight cultures of WT, Δvpa1365, Δvpa1365-vpa1365, Δvpa1365-pMMB207, Δvpa1365-vtrA, and Δvpa1365-vtrB were diluted 1:100 into 5 mL of fresh LB medium and cultured under different environmental factors (at 28°C, 40°C, pH = 8, 0.1 M NaCl, 0.5 M NaCl, 37°C with or without 0.04% bile salts) for 7 h, respectively. Next, the bacterial cultures were adjusted to an OD_600_ of 1, the supernatant protein was collected and concentrated using protein filter columns Amicon Ultracel and Regenerated Cellulose (Millipore, Billerica, MA, USA), and cell pellets were resuspended in 100 µL PBS. The 5× SDS-PAGE sample loading buffer (Beyotime, Shanghai, China) was added to the mixture, and then each 10 µL normalized sample incubated at 100°C for 10 min was separated on a 10% polyacrylamide gel and transferred to the nitrocellulose filter membranes (Solarbio, Beijing, China). After blocking in 1% bovine serum albumin for 1.5 h, the nitrocellulose filter membranes were incubated with the VopD2 antibody or RNA polymerase (RNAP) antibody at a 1:1,000 dilution and then incubated with goat anti-rabbit IgG (Sangon, Shanghai, China) at a 1:4,000 dilution. Finally, the SuperFemto ECL chemiluminescence reagent (Vazyme, Beijing, China) was used to visualize each blot.
EMSA assay
The ORF region of vpa1365 was amplified and inserted into the pColD1 plasmid with the EcoRI and BglII sites, and then the recombinant plasmid pColD1-vpa1365 was transformed into E. coli BL21 (DE3). The primers are listed in Table S1. IPTG was added to induce the expression and secretion of VPA1365 growing in 500 mL of LB medium at 16°C with 120 rpm for 16 h; subsequently, the cultures were collected, washed, lysed, and subjected to the purification of VPA1365-His protein using the His Bind Purification Kit (Sangon, Shanghai, China). All purified proteins were examined for concentration using a bicinchoninic acid assay kit (Beyotime, Shanghai, China). The DNA probes with the 5′-FAM fluorescent label were amplified and purified using the Gel Extraction Kit (Omega, USA). Each 20 µL of a different electrophoretic mobility shift (EMSA) mixture containing 30 nM of DNA-purified probes, 4 µL of 5× EMSA binding buffer (10 mM NaCl, 0.1 mM dithiothreitol, 0.1 mM EDTA, 10 mM Tris, pH 7.4), and the VPA1365-His protein was individually incubated at 25°C for 0.5 h and separated on a 6% precast PAGE gel (Beyotime, Shanghai, China). When necessary, unlabeled cold DNA was added to the mixture. Finally, Typhoon FLA 9500 was used to visualize the imprint (GE Healthcare, Uppsala, Sweden).
Quantitative real-time reverse transcription PCR analysis
Quantitative real-time reverse transcription PCR (qRT-PCR) was performed as described previously (41). In brief, the overnight cultures of WT, Δvpa1365, Δ*vpa1365-vpa1365, and Δvpa1365-*pMMB207 were grown in LB medium with 0.04% bile salts to the stationary phase, the total RNA was extracted with the bacterial RNA extraction kit (Tiangen, Beijing, China), and the RNA quality was measured by Nanodrop (Nanophotometer NP80, IMPLEN, München, Germany). All normalized samples were treated with the HiScript III All-in-One RT SuperMix kit (Vazyme, Nanjing, China) at 50°C for 15 min and then at 85°C for 15 s to reverse transcription into cDNA. The cDNA samples were determined and analyzed on the ABI PRISM 7500 Real-Time PCR System (Applied Biosystems, USA) with the ChamQ Universal SYBR qPCR Master Mix (Vazyme, Nanjing, China). Expression levels of the genes encoding T3SS2 apparatus protein VPA1342 and VPA1343, regulator protein VPA1348 (VtrB), translocator protein VPA1361 (VopD2), hypothetical protein VPA1364, effector VPA1380, type IV pilin MshA and PilA, thermostable direct hemolysin TdhA and TdhS, biofilm regulatory protein ScrG, and capsular polysaccharide-protein CpsA were determined. Primers in this study are listed in Table S1. The relative mRNA expression values of each gene normalized to gyrB gene expression were calculated by using the 2^−ΔΔCt^ method (42). Three separate experiments were performed in biological triplicates each.
Animal infection
First, the infant rabbit infection model was performed to determine the bacterial T3SS2 function and intestinal colonization assay (43). Overnight cultures of WT and Δvpa1365 were diluted 1:100 into 50 mL of fresh LB medium and cultured at 30°C for 16 h. Infant rabbits were infected with WT and Δvpa1365 at a dose of 10^9^ CFU orally, and colonized bacteria were counted by enumerating the CFU in 1 g of the small intestine tissues with gradient dilution. Three infected tissues in each group were submitted to Wuhan Servicebio Technology Co. Ltd. for pathological section analysis. The representative image was respectively presented. Besides, the zebrafish infection model was used to evaluate the effect of VPA1365 on bacterial virulence (44). Healthy Danio rerio (zebrafish, 2–2.5 cm in length) was raised in freshwater at the constant 26°C ± 2°C for 1 week before infection. Each group of 30 zebrafish cultured at constant temperature was infected with intramuscular injection at a dose of 10^6^ CFU per 5 µL of the WT, Δvpa1365, and ΔvscN2, respectively, afterward observed and recorded the number of living zebrafish every day.
Statistical analysis
All statistics were analyzed with GraphPad Prism version 7.01, and the results were summarized as mean ± SD (n = 3). The one-way analysis of variance with Dunnett’s post-test or t-test was used to determine the differences by calculating P value. The P values ≤0.05 and ≤0.01 were considered significant (*) and extremely significant (**), respectively. The P value > 0.05 was regarded as non-significant.
RESULTS
Identification and sequence analysis of vpa1365
Nucleotide sequence analysis showed that the ORF of vpa1365 contains 489 base pairs, encoding 163 amino acids within the T3SS2 gene cluster located in the small chromosome of V. parahaemolyticus RIMD2210633 (Fig. 1A). The INTERPRO and UnitProtKB analyses indicated that the VPA1365 protein contains three conserved tetratricopeptide repeat (TPR) domains, which mediate protein-protein and protein-DNA interactions (45–48), suggesting a potential role as a regulator (Fig. 1B). The structural prediction of the VPA1365 protein using SWISS-MODEL revealed a 90.68% similarity to the structure of a regulatory factor called A0A7Z7VL62_VIBCL found in V. cholerae (Fig. 1C). Additionally, phylogenetic tree analysis indicated that the sequence of vpa1365 in V. parahaemolyticus shares a relatively high degree of similarity with other Vibrio species (60%–85% amino acid identity; Fig. 1D). These findings collectively suggested that the uncharacterized VPA1365 might be a highly conserved TPR family protein commonly present in Vibrio species.
Conserved domain structure and phylogenetic tree analysis of VPA1365 in V. parahaemolyticus. (A) Genetic location and organization of the T3SS2 in V. parahaemolyticus RIMD 2210633. (B) Three blue boxes represent the conserved TPR domain predicted by the INTERPRO. (C) The VPA1365 structure prediction was accomplished using the SWISS-MODEL (the blue and orange regions indicate the amino terminal and carboxy terminal, respectively). (D) The phylogenetic tree inferred from 1,000 replicates was constructed by the neighbor-joining method using MEGA-X.
Deletion of vpa1365 reduces the intestinal colonization of V. parahaemolyticus
To further elucidate the function of VPA1365 in infant rabbit intestinal colonization, we generated the in-frame gene deletion mutant strain Δvpa1365, as well as complement strains Δvpa1365-vpa1365, Δvpa1365-pMMB207, Δvpa1365-vtrA, and Δvpa1365-vtrB. The growth curve results showed that no significant differences were observed among the WT, Δvpa1365, and complement strains when cultured at 37°C in the LB liquid medium (Fig. S1). However, after infection for 38 h, the bacterial burden in the Δvpa1365-infected small intestine of infant rabbits was significantly lower than that of the WT-infected infant rabbits (Fig. 2A; P ≤ 0.01). Moreover, the pathological analysis indicated that the unchallenged intestinal tissue exhibited long intestinal villi and an intact villous epithelium composed of columnar epithelial cells. Intestinal tissue infected with Δvpa1365 displayed short villi, increased shedding of villous epithelial cells, and an exposed lamina propria, indicating a slight injury to the tissue (Fig. 2B). However, widespread and obvious shedding of intestinal villi was observed in the intestinal tissue challenged with WT. The intestinal glands in the lamina propria, the structures of the lamina propria, and the muscle layers were not obvious, suggesting that the intestinal tissue suffered severe injury challenged with WT (Fig. 2B). These findings showed that vpa1365 did play an important part in the enterotoxicity and intestinal colonization of V. parahaemolyticus.
*Analysis of the intestinal pathological section and bacterial colonization quantity in infant rabbits. (A) The amount of WT and Δvpa1365 colonized in the small intestine. *P ≤ 0.01. (B) Observations on the small intestine were challenged with PBS, WT, and Δvpa1365, respectively. The yellow arrow, red arrow, and brown arrow represented exfoliated intestinal villous epithelial cells, damaged lamina propria, and vascular congestion, respectively. Three infected tissues from each group were analyzed, and a representative image was presented for each case. Scale bars: 50 µm.
VPA1365 regulates the expression and secretion of T3SS2 by directly binding to the promoters of vtrA
It is generally considered that V. parahaemolyticus T3SS2 is induced by bile salts and responsible for enterotoxicity and intestinal colonization (11, 26). Therefore, we evaluated the expression levels of several important T3SS2 genes in WT, Δvpa1365, Δvpa1365-vpa1365, and Δvpa1365-pMMB207 in the presence of bile salts. Transcription levels of multiple T3SS2-related genes (vpa1342, vpa1343, vtrB, vopD2, vpa1364, and vpa1380) were significantly decreased in Δvpa1365 and Δvpa1365-pMMB207 compared to that of WT, and the expression levels of these genes were restored in the Δvpa1365-vpa1365 (Fig. 3A; P ≤ 0.05). VopD2 has been described as a translocator protein secreted by the T3SS2, which serves as an indicator of T3SS2 functionality (49). Furthermore, we subsequently assessed the secretion of T3SS2 protein VopD2 in WT, Δvpa1365, Δvpa1365-vpa1365, and Δvpa1365-pMMB207 cultured in LB medium with 0.04% bile salts. An RNAP antibody was used to demonstrate that the samples were evenly loaded by assessing RNAP levels. As expected, the secretion of VopD2 in the WT was significantly higher than that in both Δvpa1365 and Δvpa1365-pMMB207 (Fig. 3B), and the complementary strain Δvpa1365-vpa1365 recovered the VopD2 secretion to WT levels. These findings indicated that VPA1365 positively regulates the expression and secretion of the T3SS2 proteins in V. parahaemolyticus.
*The effects of vpa1365 on the expression and secretion of T3SS2. (A) Relative transcript levels of the T3SS2-associated genes in WT, ∆vpa1365, ∆vpa1365-vpa1365, ∆vpa1365-vtrA, and ∆vpa1365-vtrB. Results are represented as mean ± SD (n = 3, biologically independent experiments). *P ≤ 0.05 and *P ≤ 0.01. ns, no significance. (B) The T3SS2 translocon protein VopD2 secretion and RNAP of WT, ∆vpa1365, and complemented strains in LB medium with or without 0.04% bile salts. (C) The analysis of binding of VPA1365 to the promoter of the regulator vtrA. (D) The T3SS2 translocon protein VopD2 secretion and RNAP of WT, ∆vpa1365, and complemented strains in LB medium at 28°C, 40°C, pH = 8, 0.1 M NaCl, and 0.5 M NaCl, respectively.
To further investigate the potential regulatory role of VPA1365 on the T3SS2, EMSA was performed to observe the binding of VPA1365 to the promoter of the T3SS2 effector gene vpa1380, hypothetical protein gene vpa1364, and regulator genes vtrA and vtrB in vitro. The results showed that the VPA1365 protein binds to the promoters of vtrA in a concentration-dependent manner, and the addition of unlabeled cold DNA resulted in a reduced shifted band (Fig. 3C). While no binding is observed to the promoter of the vtrB, vpa1380, and vpa1364 (Fig. S2B through D). The gyrB probe was used as a negative control, showing no binding with the VPA1365 protein (Fig. S2A). These results suggested that the VPA1365 protein binds directly to the promoter regions of vtrA to activate the expression of T3SS2-related genes, thereby positively affecting the virulence of V. parahaemolyticus.
Environmental factors have been found as a component to induce the expression and secretion of the T3SS2. To further explore this, we analyzed the expression and secretion of VopD2 under various environmental conditions (28°C, 40°C, pH = 8, 0.1 M NaCl, 0.5 M NaCl). As expected, the expression and secretion of VopD2 were observed in WT under the condition of 0.1 M NaCl (Fig. 3D), which is consistent with our previous findings (30). Meanwhile, the deletion of vpa1365 abolished the expression and secretion of VopD2 under low NaCl conditions. The complementary strain Δvpa1365-vpa1365 was restored to the level of WT (Fig. 3D), further confirming the regulation of VPA1365 to T3SS2.
Our study demonstrated that VPA1365 inhibits the expression of vtrA, a key activator of T3SS2 in V. parahaemolyticus (50). This led us to investigate whether VPA1365 regulates T3SS2 expression via VtrA. To explore this, we overexpressed VtrA or VtrB in Δvpa136 and assessed the expression levels of T3SS2-related genes using qRT-PCR, while VopD2 secretion was evaluated by western blotting. As expected, overexpression of either VtrA or VtrB in Δvpa1365 significantly increased the expression levels of T3SS2-related genes (Fig. 3A; P ≤ 0.01), which indicates that overexpression of VtrA or VtrB suppresses the reduced expression of T3SS2 and secretion of VopD2 in Δvpa1365. Additionally, overexpressing VtrA or VtrB in Δvpa1365 restored the expression of VopD2 in the whole cell under all tested environmental conditions. However, the secretion of VopD2 in Δvpa1365-vtrA or Δvpa1365-vtrB strains was observed under low NaCl conditions, and the secretion levels were lower than those in the WT and complementary strains (Fig. 3D). These results revealed the function of VPA1365 in sensing the environmental cue (low NaCl) and extensive regulatory connections between the T3SS2 positive regulator VPA1365 and the VtrA/VtrB.
Deletion of vpa1365 reduces the cytotoxicity and adhesion rate to HeLa cells
The main virulence factor T3SS2 is crucial for V. parahaemolyticus to colonize the intestine and induce massive cell death in the host by interfering with intracellular signal pathways (18, 22, 29). Therefore, we determined the cytotoxicity and adhesion rate of WT, Δvpa1365, and complement strains to HeLa cells. As shown in Fig. 4, both the cytotoxicity and adhesion rate of the Δvpa1365 were significantly lower than those of the WT (P ≤ 0.01), while complementation of this gene partially restored its cytotoxicity and adhesion rate. These results revealed that vpa1365 plays a key role in promoting adhesion ability and cytotoxicity toward HeLa cells.
*The effects of vpa1365 on cytotoxicity and cell adhesion ability. (A) Cytotoxicity and (B) adhesion rate of HeLa cell monolayers infected with WT, ∆vpa1365, ∆vpa1365-vpa1365, and ∆vpa1365-pMMB207 at 2 h, respectively. Results are presented as mean ± SD (n = 3). Three separate experiments were performed in biological triplicates each. *P ≤ 0.05 and *P ≤ 0.01.
VPA1365 promotes the hemolytic activity and biofilm formation by directly binding to the promoters of scrG, pilA, and mshA
Recent studies have revealed the significant role of biofilm and hemolytic activity in the pathogenicity of V. parahaemolyticus (50–52). Therefore, we also investigated the regulatory role of VPA1365 in these virulence factors. Our results showed that the deletion of vpa1365 resulted in significantly lower biofilm formation than that of the Δvpa1365-vpa1365 and WT (Fig. 5A; P ≤ 0.01). Furthermore, the diameters of hemolytic rings in Δvpa1365 and Δvpa1365-pMMB207 were significantly smaller than those in other tested strains (Fig. 5B; P ≤ 0.01), indicating a marked reduction in the hemolytic activity of Δvpa1365. These results revealed an important role of VPA1365 in biofilm formation and hemolytic activity.
*The effects of vpa1365 on biofilm formation and hemolytic activity. (A) The normalized biofilm formation (total amount of biofilm/growth) of WT, Δvpa1365, ∆vpa1365-vpa1365, and ∆vpa1365-pMMB207 cultured at 37°C for 31 h was stained with 0.1% crystal violet, respectively. (B) The hemolytic circle diameter of WT, Δvpa1365, ∆vpa1365-vpa1365, and ∆vpa1365-pMMB207 growing on the Wagatsuma blood agar base. (C) Relative transcript levels of virulence-associated genes in WT, ∆vpa1365, and ∆vpa1365-vpa1365. (A–C) Results are presented as mean ± SD (n = 3). Three separate experiments were performed in biological triplicates each. *P ≤ 0.05 and *P ≤ 0.01. ns, no significance. (D–F) The analysis of binding of VPA1365 to the promoter of biofilm regulator scrG, type IV pilus pilA, and mshA.
V. parahaemolyticus often relies on the pili for attachment and then forms biofilm by producing polysaccharides, which is regulated by the concentration of the second messengers. The mannose-sensitive hemagglutinin type IV pili, polysaccharides, and the second messenger bis-(3′−5′)-cyclic di-GMP (c-di-GMP) have been demonstrated to play positive roles in biofilm formation (53–55). To further explore how VPA1365 affects biofilm formation and hemolytic activity, the expression levels of type IV pilin pilA and mshA, c-di-GMP metabolism enzymatic gene scrG, capsular polysaccharide-protein cpsA, thermostable direct hemolysin tdhA, and tdhS were determined in WT and Δvpa1365. The results showed that VPA1365 positively regulates the expression of these virulence-associated genes (Fig. 5C). The EMSA was then used to determine whether VPA1365 directly binds to the selected promoters of pilA, mshA, scrG, tdhA, and tdhS genes. As expected, the VPA1365 protein directly binds to the promoters of pilA, mshA, and scrG in a concentration-dependent manner (Fig. 5D through 5F). The addition of cold DNA changed the shifted bands to an unshifted position. However, no binding was observed with the promoters of tdhA and tdhS, suggesting an indirect regulatory effect of VPA1365 on hemolytic activity (Fig. S3). In conclusion, our findings indicated that VPA1365 is directly bound to the promoter regions of pilA, mshA, and scrG, thereby regulating biofilm formation and hemolytic activity.
Identification of VPA1365-binding motifs
To determine the binding sites of VPA1365, we chose the promoter of scrG as a representative for EMSA analysis due to its more obvious binding band. Six truncated fragments of the scrG promoter were tested for their interaction with VPA1365. EMSA results indicated that VPA1365 binds to DNA fragments 1, 2, 3, 4, and 5 but not to DNA fragment 6 (Fig. 6A). Subsequently, we amplified a DNA fragment with position 140–166 truncation (ΔPscrG) and found that VPA1365 did not bind to this fragment (Fig. 6B and C). Furthermore, using the MEME-Suite tool, we identified a putative VPA1365-binding motif within the promoters of vtrA, pilA, mshA, and scrG, designated as 5′-AAAATAAAGCGCATAG-3′. The promoter region including the transcriptional initial site of the scrG gene was identified by others (56). The predicted VPA1365-binding site is highly similar to the scrG promoter region with nucleotide positions 140–166. The transcriptional initial sites in the promoter regions of the vtrA, pilA, and mshA genes were identified by previous transcriptome sequencing (Fig. S4) (9, 55). Finally, we predicted and analyzed the consensus binding motif in the promoter of scrG and other promoters of vtrA, pilA, and mshA (Fig. 6D). These results further revealed that VPA1365 binds directly to the promoter and regulates the expression levels of these genes in V. parahaemolyticus.
Identification of the VPA1365-binding motifs. (A) EMSA analysis of VPA1365 binding to the scrG promoter with different truncations. Six truncated fragments for the scrG promoter (left panel) were analyzed for their interaction with VPA1365 (right panel). (B) Nucleotide sequences (271 bp) of the scrG promoter (PscrG). The VPA1365-binding site is boxed in black, while −35 box and −10 box sequences are underlined. Start codon (ATG) and +1 of transcription are shown in red and with an arrow, respectively. (C) EMSA analysis of VPA1365 binding to the truncated ∆scrG promoter (∆PscrG) with the deletion of the VPA1365-binding site (140–166 bp). The fragment of ∆PscrG (left panel) was analyzed for its binding with VPA1365 (right panel). (D) The predicted consensus VPA1365-binding motif was generated from MEME-Suite analysis of the selected promoters.
Lacking of vpa1365 attenuates virulence to D. rerio
Given the significant impairment of virulence factors in Δvpa1365, we sought to determine the dependency of V. parahaemolyticus pathogenicity on the VPA1365. Healthy D. rerio (zebrafish) was infected with the WT, Δvpa1365, and ΔvscN2 strains, each at a concentration of 1 × 10^6^ CFU in the muscle. In the PBS-negative control group, no zebrafish died, whereas, in the WT-infected group, zebrafish died daily, resulting in complete mortality by day 6 (Fig. 7; P ≤ 0.01). The relative survival rates of zebrafish infected with Δvpa1365 and ΔvscN2 were approximately 73% and 63%, respectively. These results provided additional evidence that VPA1365 is essential for the virulence of V. parahaemolyticus.
*The survival rate of zebrafish challenged with WT, Δvpa1365, and ∆vscN2, respectively. The number of dead zebrafish was recorded, and the percent survival rate in every group was calculated. The comparison of survival distribution between WT and mutant strains was performed using the log-rank (Mantel-Cox) test in GraphPad Prism version 7.0. *P ≤ 0.01.
DISCUSSION
Since the early discovery of the structure of the T3SS in pathogenic Yersinia spp. (57), two T3SSs, namely T3SS1 and T3SS2, respectively, have been identified in V. parahaemolyticus. The T3SS1 has been found in nearly all sequenced clinical or environmental isolates. In contrast, the occurrence of T3SS2 in pathogenic islands remains limited to pathogenic strains (11, 58), thereby suggesting an essential role for T3SS2 in the pathogenesis of V. parahaemolyticus. An increasing amount of evidence has indicated that the virulence genes of V. parahaemolyticus are stringently controlled by numerous regulators (6, 59). Previous studies have demonstrated that VtrA/B/C activates the expression of the T3SS2 gene cluster in the presence of bile salts (25). In this study, we identified and characterized a protein belonging to the TPR family, namely VPA1365, which positively regulates the function of the T3SS2. This discovery broadens the regulatory network of virulence genes and deepens our understanding of the pathogenic molecular mechanisms of V. parahaemolyticus.
When pathogens invade the host intestine, bile acids bind to the hydrophobic interior of the β barrel within the periplasmic spatial domains of VtrA and VtrC, thereby promoting the expression of the downstream gene vtrB (60). Subsequently, the activated VtrB induces the expression of T3SS2-related genes. Notably, both VtrA and VtrB belong to the OmpR family, which encompasses a DNA-binding domain featuring a winged-helix-turn-helix structure (24, 61). The transcriptional factor ToxR has also been discovered to be indispensable for the induction of vtrB and T3SS2 transcription (62, 63). These findings imply that the expression of T3SS2 was tightly regulated by many regulators. Moreover, Troy et al. screened 230 genes that play a vital role in the colonization of V. parahaemolyticus in the small intestine using Tn-seq (27). Intriguingly, among them are multiple potential transcriptional regulators, some of which have been affirmed to directly regulate the expression of flagellar-related genes and contribute to the motility of V. parahaemolyticus (9, 64). Therefore, we were eager to know whether these regulators have an impact on T3SS2, which has been identified as essential for the colonization in the small intestine of V. parahaemolyticus (27). Among them, we discovered that the protein VPA1365, located within the T3SS2 gene cluster, possesses three TPR domains and is predicted to have the potential to interact with other nucleotides. Our study subsequently confirmed that the deletion of vpa1365 significantly reduced the virulence and colonization ability of V. parahaemolyticus in infant rabbits (P ≤ 0.01). Thus, we further hypothesized that VPA1365 might affect the colonization of V. parahaemolyticus in the rabbit intestine by regulating the function of T3SS2.
The TPR domain contains two antiparallel α-helix subunits that display a right-handed supercoil structure, enhancing the ability of the channel to effectively interact with the target protein (65, 66). More recently, it was found that TPR domains are present in numerous DNA- and RNA-binding proteins (45–48). In a previous study, the T3SS2 protein PthA containing a TPR-like domain in Xanthomonas axonopodis pv. citri was shown to undergo conformational changes upon DNA interaction, similar to pentatricopeptide repeat domain, a nucleic acid-binding motif, further revealing its nucleic acid binding ability (65). Until now, the TPR family is widely associated with diverse biological processes, including metabolic regulation, gene expression, and protein export (66). The biological properties of many TPR family genes have been extensively studied in plants like Solanum lycopersicum and Arabidopsis (67, 68); unfortunately, their roles in microorganisms remain relatively limited. In avian pathogenic E. coli, the T3SS chaperone protein YgeG possessing TPR domains has an impact on virulence and pathogenicity (69). In Borrelia burgdorferi, a putative TPR domain is of great significance for the function of the hypothetical protein BB0238, and mutation of key residues in the α-helix significantly reduces tissue infection rates in the C3H/HeN mice infection model (70). Furthermore, two diguanylate cyclase genes, scrJ (VPA1115) and scrL (VPA1069), were identified to contain conserved TPR domains and coupled GGDEF proteins, contributing to the surface colonization of V. parahaemolyticus (71). Current studies have demonstrated a close connection between the TPR domain and virulence in pathogens, warranting further exploration of its molecular pathogenic mechanism. Our study found that VPA1365, containing three conserved TPR domains, directly binds to the promoter regions of the T3SS2 master regulatory factor vtrA (Fig. 3). Both qRT-PCR and immunoblotting results further confirmed that the absence of vpa1365 decreased the expression levels of T3SS2-related genes and the secretion of VopD2 (Fig. 3). Therefore, we proposed that VPA1365 directly binds to the vtrA promoter and then regulates the expression of vtrB and T3SS2-related genes.
Interestingly, overexpression of the T3SS2 positive regulator VtrA or VtrB in Δvpa1365 not only upregulates the expression level of T3SS2-related genes but also increases the secretion of VopD2 into the supernatant under bile salts conditions. These results suggested that VPA1365 may regulate the expression of T3SS2 through VtrA/VtrB, warranting further exploration of the relationship between VPA1365 and VtrA/VtrB. Furthermore, environmental factors also influence the expression of T3SS2, and we found that the absence of VPA1365 completely abolishes the expression and secretion of VopD2 under low NaCl conditions (Fig. 3). This finding aligns with our previous discovery that low NaCl affects the T3SS2 function in V. parahaemolyticus (30), indicating that low NaCl might serve as a signal influencing VPA1365-mediated regulation of T3SS2. However, under low NaCl conditions, overexpression of either VtrA or VtrB in the Δvpa1365 did not restore VopD2 secretion to levels comparable to those in WT and Δ*vpa1365-*vpa1365, further highlighting the importance of VPA1365 in positively regulating the function of T3SS2 in response to low NaCl. Additionally, the high conservation of VPA1365 in the different Vibrio species suggests that this regulatory mechanism may be widespread among other Vibrio species.
It is generally considered that V. parahaemolyticus T3SS1 is mainly associated with cytotoxicity, while T3SS2 is primarily connected to enterotoxicity (16). However, previous studies have revealed that VtrA/VtrB is also of vital importance for T3SS2-dependent cytotoxicity (72, 73). The comparative genome analysis indicated that the Vp-PAI encompasses not only the T3SS2 gene cluster but also two tdh genes (tdhA and tdhS) (15, 74). Deletion of vtrA or vtrB decreased the production of TDH, the major virulent determinant influencing cytotoxicity and enterotoxicity, in V. parahaemolyticus (20, 75). Additionally, motility, biofilm formation, and adhesion ability play important roles in the infection process of V. parahaemolyticus (76). Therefore, we sought to determine whether VPA1365 has an impact on these crucial virulence factors. In comparison to the WT, deletion of vpa1365 significantly reduced cytotoxicity, adhesion rate, biofilm formation, and hemolytic activity ([Fig. 4, Fig. 5A and B](#F4 F5)), while it had no effect in affecting the swimming and swarming motility (Fig. S5). The biofilm formation in Δ*vpa1365-*vpa1365 did not fully restore to the level of WT, possibly because overexpression of VPA1365 might also regulate the other regulators of biofilm formation. The EMSA results revealed an interesting finding: the VPA1365 directly binds to the promoter regions of type IV pilin mshA, pilA, and biofilm regulatory protein scrG. However, it is unable to directly bind to the promoter regions of the thermostable direct hemolysin tdhA and tdhS. These results suggested that VPA1365 might act as a global regulatory factor to influence the expression level of various virulence genes. Moreover, it may indirectly regulate hemolysin activity by promoting the expression level of T3SS2 regulator vtrA or other transcriptional factors. Based on the truncated EMSA analysis and the MEME-Suite tool, we predicted a putative VPA1365-binding motif within the promoters of vtrA, pilA, mshA, and scrG designated as 5′-AAAATAAAGCGCATAG-3′. This motif bears some resemblance to the previously reported DNA sequence bound with a TPR-like protein PthA, which is a TA-rich region (65). Interestingly, the nuc2+ protein containing the TPR domain in Schizosaccharomyces pombe is also capable of binding AT-rich DNA in vitro (46). These findings suggested that the fragments containing AT-rich regions are likely to be the fragments of proteins bound with the TPR domain. Finally, considering the relatively weak interactions of VPA1365 with these promoters and the involvement of the TPR domain in protein-protein interactions, VPA1365 may exert its effect on transcription in an alternative manner, such as through interactions with other nucleoid-binding proteins like H-NS. Regardless, the positive regulatory effect of VPA1365 on the function of T3SS2 and virulence-related genes is unquestionable. However, whether VPA1365 interacts with other DNA-binding proteins to co-regulate the expression of T3SS2 remains to be explored in the future.
Zebrafish commonly serve as an ideal model to investigate the virulence mechanisms employed by both Gram-negative and Gram-positive pathogens that affect fish and humans (77). Currently, zebrafish have been employed as an animal model to assess the virulence of V. anguillarum, V. cholerae, and V. parahaemolyticus (44, 78, 79). Therefore, we used the zebrafish as an infection model to evaluate the influence of VPA1365 on the virulence of zoonotic V. parahaemolyticus in fish and humans. The survival rates of zebrafish infected with Δvpa1365 and ΔvscN2 (ΔvscN2 serving as a T3SS2 functional reference) were approximately 73% and 63%, respectively, whereas all zebrafish died by day 6 in the WT-infected group. Notably, the survival rate of Δvpa1365 was significantly higher than that of ΔvscN2, suggesting that the impact of the regulatory factor VPA1365 on virulence includes but is not limited to the T3SS2. Nevertheless, the deletion of vpa1365 significantly reduced the lethality and colonization ability of V. parahaemolyticus in infant rabbits and zebrafish.
Conclusion
In the present study, we identified a protein belonging to the TPR family, namely VPA1365, which is located within the T3SS2 gene cluster. This particular protein directly binds to the promoter regions to regulate the expression of T3SS2, thereby exerting a significant influence on the intestinal colonization of V. parahaemolyticus. Additionally, VPA1365 functions as a global regulator, modulating the hemolytic activity, biofilm formation, adhesion ability, and cytotoxicity of V. parahaemolyticus. Its regulation of biofilm formation and adhesion likely occurs by directly binding to the promoter regions of pilA, mshA, and scrG (Fig. 8). Overall, this study established the regulatory networks of VPA1365 and its contribution to diverse virulence factors of V. parahaemolyticus.
The putative model of VPA1365 regulation in V. parahaemolyticus. VPA1365 indirectly contributes to the function of T3SS2 by binding to the promoter of vtrA and additionally positively regulates hemolytic activity, biofilm formation, cytotoxicity, and adhesion ability. The black solid arrows indicate direct binding or regulation, and the black dashed arrows indicate indirect or unclear regulation.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Li M, Xu H, Tian Y, Zhang Y, Jiao X, Gu D. 2023. Comparative genomic analysis reveals the potential transmission of Vibrio parahaemolyticus from freshwater food to humans. Food Microbiol 113:104277. doi:10.1016/j.fm.2023.10427737098434 · doi ↗ · pubmed ↗
- 2Lei T, Jiang F, He M, Zhang J, Zeng H, Chen M, Pang R, Wu S, Wei L, Wang J, Ding Y, Wu Q. 2020. Prevalence, virulence, antimicrobial resistance, and molecular characterization of fluoroquinolone resistance of Vibrio parahaemolyticus from different types of food samples in China. Int J Food Microbiol 317:108461. doi:10.1016/j.ijfoodmicro.2019.10846131794931 · doi ↗ · pubmed ↗
- 3Tan CW, Rukayadi Y, Hasan H, Thung TY, Lee E, Rollon WD, Hara H, Kayali AY, Nishibuchi M, Radu S. 2020. Prevalence and antibiotic resistance patterns of Vibrio parahaemolyticus isolated from different types of seafood in Selangor, Malaysia. Saudi J Biol Sci 27:1602–1608. doi:10.1016/j.sjbs.2020.01.00232489301 PMC 7253911 · doi ↗ · pubmed ↗
- 4Ceccarelli D, Hasan NA, Huq A, Colwell RR. 2013. Distribution and dynamics of epidemic and pandemic Vibrio parahaemolyticus virulence factors. Front Cell Infect Microbiol 3:97. doi:10.3389/fcimb.2013.0009724377090 PMC 3858888 · doi ↗ · pubmed ↗
- 5Lovell CR. 2017. Ecological fitness and virulence features of Vibrio parahaemolyticus in estuarine environments. Appl Microbiol Biotechnol 101:1781–1794. doi:10.1007/s 00253-017-8096-928144705 · doi ↗ · pubmed ↗
- 6Li L, Meng H, Gu D, Li Y, Jia M. 2019. Molecular mechanisms of Vibrio parahaemolyticus pathogenesis. Microbiol Res 222:43–51. doi:10.1016/j.micres.2019.03.00330928029 · doi ↗ · pubmed ↗
- 7Austin B. 2010. Vibrios as causal agents of zoonoses. Vet Microbiol 140:310–317. doi:10.1016/j.vetmic.2009.03.01519342185 · doi ↗ · pubmed ↗
- 8Gu D, Zhang Y, Wang K, Li M, Jiao X. 2022. Characterization of the Rpo N regulon reveals the regulation of motility, T 6SS 2 and metabolism in Vibrio parahaemolyticus. Front Microbiol 13:1025960. doi:10.3389/fmicb.2022.102596036620062 PMC 9817140 · doi ↗ · pubmed ↗