Distinct HLA Haplotypes Are Associated With an Altered Strength of SARS‐CoV‐2‐Specific T‐Cell Responses and Unfavorable Disease Courses
C. Dörnte, A. Datsi, V. Traska, J. Kostyra, M. Wagner, O. Brauns, C. Lamsfuß, H. Winkels, V. Balz, J. Enczmann, O. Adams, L. Mueller, H. Baurmann, B. Eiz‐Vesper, A. Bonifacius, R. V. Sorg, C. Dose, J. Schmitz, A. Richter, J. Fischer, M. Schuster

TL;DR
This study finds that certain HLA haplotypes are linked to weaker T-cell responses and worse outcomes in COVID-19, likely due to T-cell issues rather than fewer virus epitopes.
Contribution
The study identifies T-cell intrinsic factors, not epitope presentation, as key to disease severity and predicts stable T-cell immunity against future SARS-CoV-2 variants.
Findings
HLA haplotypes linked to severe disease do not show reduced epitope presentation potential.
T-cell intrinsic factors likely influence disease severity rather than epitope availability.
T-cell epitopes are not from highly mutated regions, suggesting stable T-cell immunity against new variants.
Abstract
Infection with SARS‐CoV‐2 results in mild to severe COVID‐19 disease courses. Several studies showed the association of impaired T‐cell responses and certain HLA haplotypes with disease severity. However, it remained unclear if T‐cell activation was compromised due to a general reduction of presented epitopes or other intrinsic factors within APCs or T cells. Furthermore, a potential reduction of presented epitopes would suggest if an upcoming SARS‐CoV‐2 variant could escape T‐cell immunity. Hence, knowledge about the T‐cell epitope landscape of SARS‐CoV‐2 would allow to better understand mechanisms leading to severe disease and to estimate the potential stability of the T‐cell response in light of virus evolution, which might provide insights for future vaccine designs. Hence, in the present study, the T‐cell epitope landscape of SARS‐CoV‐2 was determined via in vitro T‐cell…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
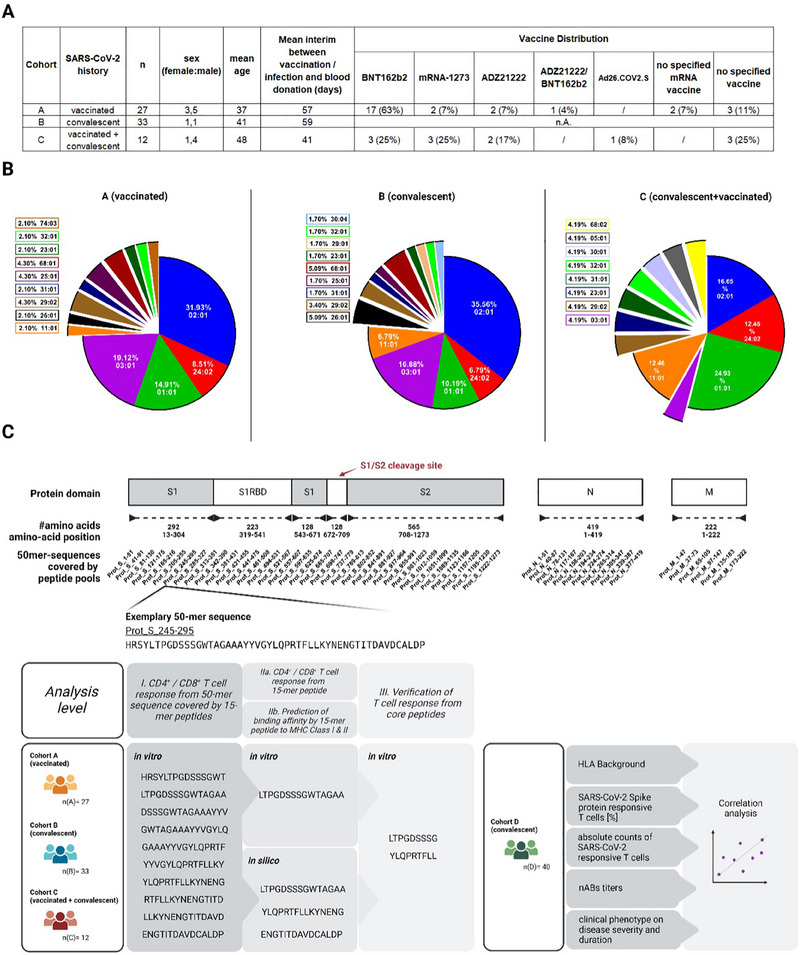 FIGURE 1
FIGURE 1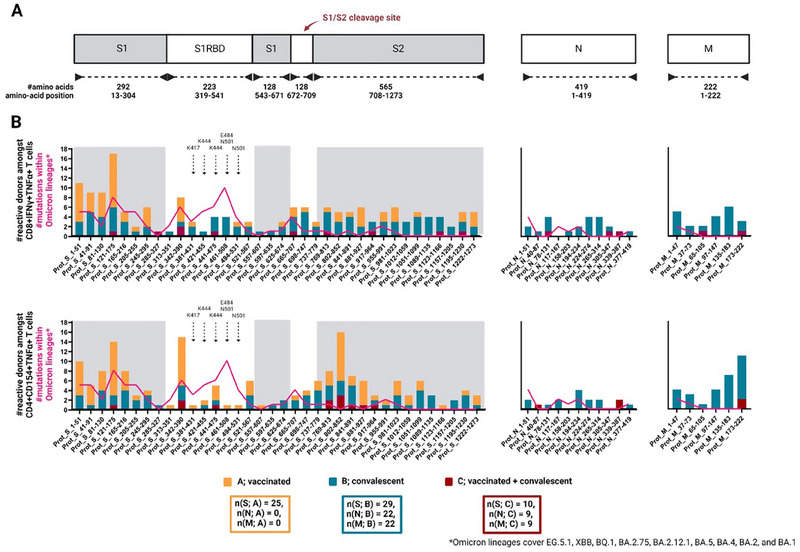 FIGURE 2
FIGURE 2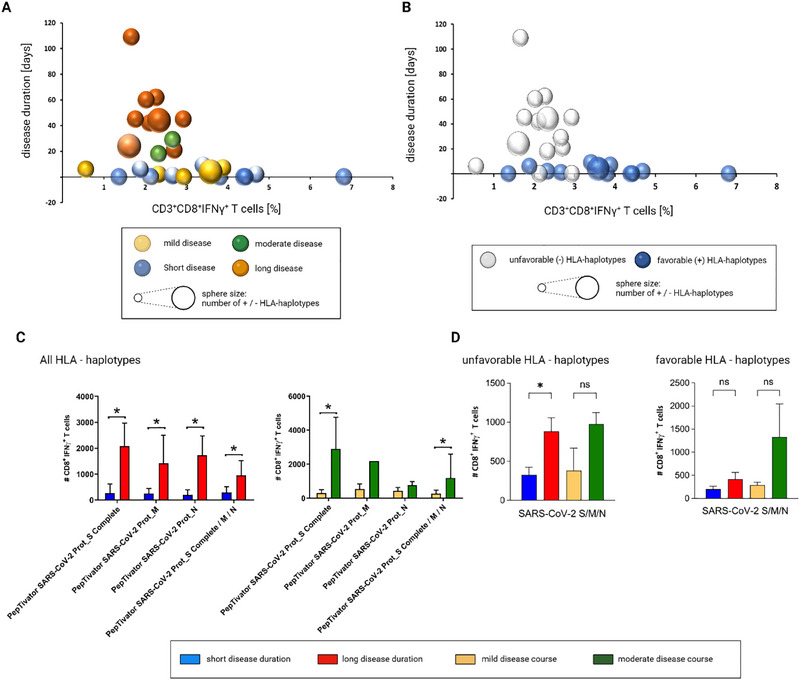 FIGURE 3
FIGURE 3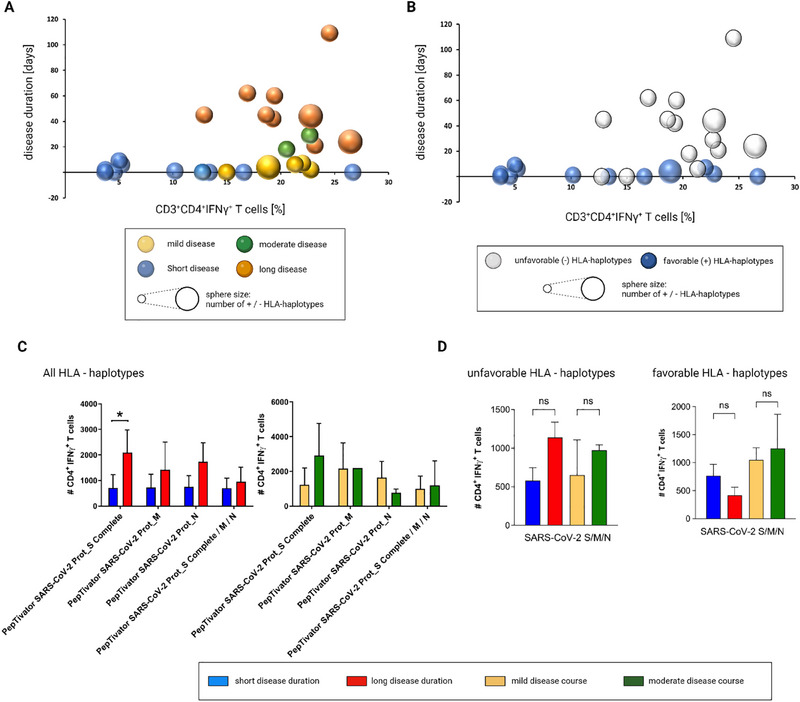 FIGURE 4
FIGURE 4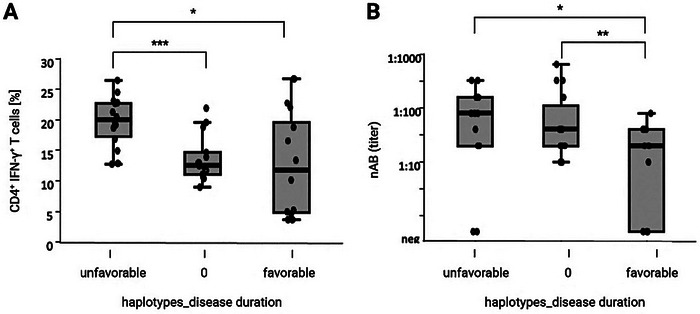 FIGURE 5
FIGURE 5| Protein | Peptide Pool | Sequence covered by 15‐mer peptide pools |
Number of reactive donors among CD8+TNFɑ+IFNɣ+ T cells (frequency of total) |
Number of reactive donors among CD4+CD154+TNFɑ+ T cells (frequency of total) | ||||
|---|---|---|---|---|---|---|---|---|
| Cohort A | Cohort B | Cohort C | Cohort A | Cohort B | Cohort C | |||
| Spike | Prot_S_1‐51 | MFVFLVLLPLVSSQCVNLTTRTQLPPAYTNSFTRGVYYPDKVFRSSVLHST | 8 (72.7) | 3 (27.3) | 0 | 3 (30) | 7 (70) | 0 |
| Prot_S_41‐91 | KVFRSSVLHSTQDLFLPFFSNVTWFHAIHVSGTNGTKRFDNPVLPFNDGVY | 4 (44.4) | 5 (55.6) | 0 | 1 (20) | 4 (80) | 0 | |
| Prot_S_81‐130 | NPVLPFNDGVYFASTEKSNIIRGWIFGTTLDSKTQSLLIVNNATNVVIKV | 5 (55.6) | 4 (44.4) | 0 | 4 (50) | 4 (50) | 0 | |
| Prot_S_121‐175 | NNATNVVIKVCEFQFCNDPFLGVYYHKNNKSWMESEFRVYSSANNCTFEYVSQPF | 11 (64.7) | 5 (29.4) | 1 (5.9) | 2 (15.4) | 10 (76.9) | 1 (7.7) | |
| Prot_S_165‐216 | NCTFEYVSQPFLMDLEGKQGNFKNLREFVFKNIDGYFKIYSKHTPINLVRDL | 1 (25) | 3 (75) | 0 | 4 (50) | 4 (50) | 0 | |
| Prot_S_205‐255 | SKHTPINLVRDLPQGFSALEPLVDLPIGINITRFQTLLALHRSYLTPGDSS | 1 (50) | 1 (50) | 0 | 2 (66.7) | 1 (33.3) | 0 | |
| Prot_S_245‐295 | HRSYLTPGDSSSGWTAGAAAYYVGYLQPRTFLLKYNENGTITDAVDCALDP | 4 (66.7) | 2 (33.3) | 0 | 3 (75) | 1 (25) | 0 | |
| Prot_S_285‐327 | ITDAVDCALDPLSETKCTLKSFTVEKGIYQTSNFRVQPTESIV | 0 | 0 | 1 (100) | 1 (100) | 0 | 0 | |
| Prot_S_313‐351 | YQTSNFRVQPTESIVRFPNITNLCPFGEVFNATRFASVY | 0 | 1 (100) | 0 | 0 | 1 (100) | 0 | |
| Prot_S_342‐390 | FNATRFASVYAWNRKRISNCVADYSVLYNSASFSTFKCYGVSPTKLNDL | 5 (62.5) | 1 (12.5) | 2 (25) | 3 (20) | 10 (66.7) | 2 (13.3) | |
| Prot_S_381‐431 | GVSPTKLNDLCFTNVYADSFVIRGDEVRQIAPGQTGKIADYNYKLPDDFTG | 1 (33.3) | 2 (66.7) | 0 | 0 | 1 (100) | 0 | |
| Prot_S_421‐455 | YNYKLPDDFTGCVIAWNSNNLDSKVGGNYNYLYRL | 0 | 1 (100) | 0 | 1 (50) | 1 (50) | 0 | |
| Prot_S_441‐475 | LDSKVGGNYNYLYRLFRKSNLKPFERDISTEIYQA | 0 | 3 (75) | 1 (25) | 1 (20) | 3 (60) | 1 (20) | |
| Prot_S_461‐508 | LKPFERDISTEIYQAGSTPCNGVEGFNCYFPLQSYGFQPTNGVGYQPY | 0 | 4 (100) | 0 | 0 | 1 (100) | 0 | |
| Prot_S_494‐531 | SYGFQPTNGVGYQPYRVVVLSFELLHAPATVCGPKKST | 1 (33.3) | 2 (66.7) | 0 | 0 | 1 (100) | 0 | |
| Prot_S_521‐567 | PATVCGPKKSTNLVKNKCVNFNFNGLTGTGVLTESNKKFLPFQQFGR | 1 (33.3) | 2 (66.7) | 0 | 3 (50) | 3 (50) | 0 | |
| Prot_S_557‐607 | KKFLPFQQFGRDIADTTDAVRDPQTLEILDITPCSFGGVSVITPGTNTSNQ | 0 | 1 (100) | 0 | 0 | 1 (100) | 0 | |
| Prot_S_597‐635 | VITPGTNTSNQVAVLYQDVNCTEVPVAIHADQLTPTWRV | 0 | 1 (100) | 0 | 1 (100) | 0 | 0 | |
| Prot_S_625‐674 | HADQLTPTWRVYSTGSNVFQTRAGCLIGAEHVNNSYECDIPIGAGICASY | 1 (33.3) | 2 (66.7) | 0 | 2 (100) | 0 | 0 | |
| Prot_S_665‐707 | PIGAGICASYQTQTNSPRRARSVASQSIIAYTMSLGAENSVAY | 2 (33.3) | 3 (50) | 1 (16.7) | 2 (66.7) | 1 (33.3) | 0 | |
| Prot_S_698‐747 | SLGAENSVAYSNNSIAIPTNFTISVTTEILPVSMTKTSVDCTMYICGDST | 1 (16.7) | 5 (83.3) | 0 | 3 (100) | 0 | 0 | |
| Prot_S_737‐779 | DCTMYICGDSTECSNLLLQYGSFCTQLNRALTGIAVEQDKNTQ | 1 (33.3) | 2 (66.7) | 0 | 4 (66.7) | 2 (33.3) | 0 | |
| Prot_S_769‐813 | GIAVEQDKNTQEVFAQVKQIYKTPPIKDFGGFNFSQILPDPSKPS | 1 (16.7) | 4 (66.7) | 1 (16.7) | 3 (37.5) | 3 (37.5) | 2 (25) | |
| Prot_S_802‐852 | FSQILPDPSKPSKRSFIEDLLFNKVTLADAGFIKQYGDCLGDIAARDLICA | 2 (40) | 3 (60) | 0 | 3 (18.8) | 10 (62.5) | 3 (18.8) | |
| Prot_S_841‐891 | LGDIAARDLICAQKFNGLTVLPPLLTDEMIAQYTSALLAGTITSGWTFGAG | 1 (20) | 4 (80) | 0 | 4 (66.7) | 1 (16.7) | 1 (16.7) | |
| Prot_S_881‐927 | TITSGWTFGAGAALQIPFAMQMAYRFNGIGVTQNVLYENQKLIANQF | 4 (66.7) | 2 (33.3) | 0 | 0 | 6 (85.7) | 1 (14.3) | |
| Prot_S_917‐964 | YENQKLIANQFNSAIGKIQDSLSSTASALGKLQDVVNQNAQALNTLVK | 0 | 2 (66.7) | 1 (33.3) | 1 (33.3) | 1 (33.3) | 1 (33.3) | |
| Prot_S_955‐991 | NAQALNTLVKQLSSNFGAISSVLNDILSRLDKVEAEV | 2 (40) | 3 (60) | 0 | 1 (50) | 1 (50) | 0 | |
| Prot_S_981‐1023 | LSRLDKVEAEVQIDRLITGRLQSLQTYVTQQLIRAAEIRASAN | 3 (50) | 3 (50) | 0 | 3 (50) | 3 (50) | 0 | |
| Prot_S_1012‐1059 | LIRAAEIRASANLAATKMSECVLGQSKRVDFCGKGYHLMSFPQSAPHG | 0 | 3 (100) | 0 | 1 (33.3) | 2 (66.7) | 0 | |
| Prot_S_1051‐1099 | SFPQSAPHGVVFLHVTYVPAQEKNFTTAPAICHDGKAHFPREGVFVSNG | 1 (20) | 4 (80) | 0 | 4 (66.7) | 2 (33.3) | 0 | |
| Prot_S_1089‐1135 | FPREGVFVSNGTHWFVTQRNFYEPQIITTDNTFVSGNCDVVIGIVNN | 0 | 4 (100) | 0 | 3 (100) | 0 | 0 | |
| Prot_S_1123‐1166 | SGNCDVVIGIVNNTVYDPLQPELDSFKEELDKYFKNHTSPDVDL | 0 | 3 (75) | 1 (25) | 0 | 1 (100) | 0 | |
| Prot_S_1157‐1205 | KNHTSPDVDLGDISGINASVVNIQKEIDRLNEVAKNLNESLIDLQELGK | 0 | 2 (100) | 0 | 2 (100) | 0 | 0 | |
| Prot_S_1195‐1230 | ESLIDLQELGKYEQYIKWPWYIWLGFIAGLIAIVMV | 2 (40) | 2 (40) | 1 (20) | 3 (75) | 1 (25) | 0 | |
| Prot_S_1222‐1273 | AGLIAIVMVTIMLCCMTSCCSCLKGCCSCGSCCKFDEDDSEPVLKGVKLHYT | 3 (60) | 2 (40) | 0 | 1 (33.3) | 2 (66.7) | 0 | |
| Nucleocapsid | Prot_N_1‐51 | MSDNGPQNQRNAPRITFGGPSDSTGSNQNGERSGARSKQRRPQGLPNNTAS | 0 | 2 (100) | 0 | 2 (100) | 0 | 0 |
| Prot_N_40‐87 | RRPQGLPNNTASWFTALTQHGKEDLKFPRGQGVPINTNSSPDDQIGYY | 0 | 1 (100) | 0 | 0 | 0 | 1 (100) | |
| Prot_N_78‐131 | SSPDDQIGYYRRATRRIRGGDGKMKDLSPRWYFYYLGTGPEAGLPYGANK | 0 | 3 (75) | 1 (25) | 1 (100) | 0 | 0 | |
| Prot_N_117‐167 | PEAGLPYGANKDGIIWVATEGALNTPKDHIGTRNPANNAAIVLQLPQGTTL | 0 | 0 | 0 | 2 (100) | 0 | 0 | |
| Prot_N_158‐203 | VLQLPQGTTLPKGFYAEGSRGGSQASSRSSSRSRNSSRNSTPGSSR | 0 | 1 (100) | 0 | 0 | 0 | 0 | |
| Prot_N_194‐234 | SRNSTPGSSRGTSPARMAGNGGDAALALLLLDRLNQLESKM | 0 | 2 (100) | 0 | 4 (100) | 0 | 0 | |
| Prot_N_224‐274 | LDRLNQLESKMSGKGQQQQGQTVTKKSAAEASKKPRQKRTATKAYNVTQAF | 0 | 4 (100) | 0 | 2 (100) | 0 | 0 | |
| Prot_N_265‐314 | TKAYNVTQAFGRRGPEQTQGNFGDQELIRQGTDYKHWPQIAQFAPSASAF | 0 | 4 (100) | 0 | 2 (100) | 0 | 0 | |
| Prot_N_305‐347 | AQFAPSASAFFGMSRIGMEVTPSGTWLTYTGAIKLDDKDPNFK | 0 | 1 (50) | 1 (50) | 0 | 0 | 0 | |
| Prot_N_339‐387 | LDDKDPNFKDQVILLNKHIDAYKTFPPTEPKKDKKKKADETQALPQRQK | 0 | 0 | 1 (100) | 0 | 0 | 2 (100) | |
| Prot_N_377‐419 | DETQALPQRQKKQQTVTLLPAADLDDFSKQLQQSMSSADSTQA | 0 | 1 (100) | 0 | 1 (100) | 0 | 0 | |
| Membrane | Prot_M_1‐47 | MADSNGTITVEELKKLLEQWNLVIGFLFLTWICLLQFAYANRNRFLY | 0 | 5 (100) | 0 | 4 (100) | 0 | 0 |
| Prot_M_37‐73 | FAYANRNRFLYIIKLIFLWLLWPVTLACFVLAAVYRI | 0 | 2 (100) | 0 | 3 (100) | 0 | 0 | |
| Prot_M_65‐105 | FVLAAVYRINWITGGIAIAMACLVGLMWLSYFIASFRLFAR | 0 | 1 (50) | 1 (50) | 1 (100) | 0 | 0 | |
| Prot_M_97‐147 | IASFRLFARTRSMWSFNPETNILLNVPLHGTILTRPLLESELVIGAVILRG | 0 | 4 (100) | 0 | 4 (100) | 0 | 0 | |
| Prot_M_135‐183 | ESELVIGAVILRGHLRIAGHHLGRCDIKDLPKEITVATSRTLSYYKLGA | 0 | 6 (100) | 0 | 7 (100) | 0 | 0 | |
| Prot_M_173‐222 | SRTLSYYKLGASQRVAGDSGFAAYSRYRIGNYKLNTDHSSSSDNIALLVQ | 0 | 2 (66.7) | 1 (33.3) | 9 (81.8) | 0 | 2 (18.2) | |
| 15‐mer peptide | Ancestor peptide pool | Peptide sequence | # Reactive donors | Predicted HLA restriction | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| CoV_Prot_S_1‐14 | CoV_Prot_S_1‐51 | MFVFLVLLPLVSSQ | 3 | HLA‐C03:04 | HLA‐C02:02 | |||||
| CoV_Prot_S_1‐14 | CoV_Prot_S_1‐51 | MFVFLVLLPLVSSQ | 1 | DRB1_1501 | ||||||
| CoV_Prot_S_5‐19 | CoV_Prot_S_1‐51 | LVLLPLVSSQCVNLT | 2 | HLA‐B51:01 | HLA‐C03:04 | |||||
| CoV_Prot_S_10‐24 | CoV_Prot_S_1‐51 | LVSSQCVNLTTRTQL | 1 | HLA‐C07:04 | ||||||
| CoV_Prot_S_13‐27 | CoV_Prot_S_1‐51 | SQCVNLTTRTQLPPA | 1 | HLA‐B37:01 | ||||||
| CoV_Prot_S_13‐27 | CoV_Prot_S_1‐51 | SQCVNLTTRTQLPPA | 1 | DRB1_1301 | ||||||
| CoV_Prot_S_18‐32 | CoV_Prot_S_1‐51 | LTTRTQLPPAYTNSF | 1 | DRB1_0101 | ||||||
| CoV_Prot_S_24‐38 | CoV_Prot_S_1‐51 | LPPAYTNSFTRGVYY | 2 | HLA‐C15:02 | ||||||
| CoV_Prot_S_29‐43 | CoV_Prot_S_1‐51 | TNSFTRGVYYPDKVF | 2 | HLA‐C02:02 | HLA‐C07:01 | |||||
| CoV_Prot_S_29‐43 | CoV_Prot_S_1‐51 | TNSFTRGVYYPDKVF | 1 | DRB1_0401 | ||||||
| CoV_Prot_S_33‐47 | CoV_Prot_S_1‐51 | TRGVYYPDKVFRSSV | 1 | HLA‐B15:01 | ||||||
| CoV_Prot_S_33‐47 | CoV_Prot_S_1‐51 | TRGVYYPDKVFRSSV | 2 | DRB1_0401 | ||||||
| CoV_Prot_S_37‐51 | CoV_Prot_S_1‐51 | YYPDKVFRSSVLHST | 1 | DRB1_0401 | ||||||
| CoV_Prot_S_41‐55 | CoV_Prot_S_1‐51 | KVFRSSVLHSTQDLF | 1 | HLA‐B14:01 | ||||||
| CoV_Prot_S_44‐58 | CoV_Prot_S_1‐51 | RSSVLHSTQDLFLPF | 2 | HLA‐A26:01 | ||||||
| CoV_Prot_S_44‐58 | CoV_Prot_S_41‐91 | RSSVLHSTQDLFLPF | 1 | DRB1_0101 | ||||||
| CoV_Prot_S_49‐63 | CoV_Prot_S_41‐91 | HSTQDLFLPFFSNVT | 1 | HLA‐A26:01 | ||||||
| CoV_Prot_S_57‐71 | CoV_Prot_S_41‐91 | PFFSNVTWFHAIHVS | 1 | HLA‐C15:02 | ||||||
| CoV_Prot_S_57‐71 | CoV_Prot_S_41‐91 | PFFSNVTWFHAIHVS | 1 | DRB1_0101 | ||||||
| CoV_Prot_S_62‐76 | CoV_Prot_S_41‐91 | VTWFHAIHVSGTNGT | 2 | HLA‐C15:02 | ||||||
| CoV_Prot_S_62‐76 | CoV_Prot_S_41‐91 | VTWFHAIHVSGTNGT | 1 | DRB1_0101 | ||||||
| CoV_Prot_S_72‐86 | CoV_Prot_S_41‐91 | GTNGTKRFDNPVLPF | 1 | HLA‐B27:05 | ||||||
| CoV_Prot_S_72‐86 | CoV_Prot_S_41‐91 | GTNGTKRFDNPVLPF | 1 | DRB1_1501 | ||||||
| CoV_Prot_S_77‐91 | CoV_Prot_S_41‐91 | KRFDNPVLPFNDGVY | 1 | HLA‐B27:05 | ||||||
| CoV_Prot_S_81‐95 | CoV_Prot_S_81‐130 | NPVLPFNDGVYFAST | 1 | HLA‐B35:01 | ||||||
| CoV_Prot_S_117‐130 | CoV_Prot_S_81‐130 | LLIVNNATNVVIKV | 1 | HLA‐B51:01 | ||||||
| CoV_Prot_S_117‐130 | CoV_Prot_S_81‐130 | LLIVNNATNVVIKV | 1 | DRB1_0401 | ||||||
| CoV_Prot_S_121‐135 | CoV_Prot_S_121‐175 | NNATNVVIKVCEFQF | 1 | HLA‐B51:01 | ||||||
| CoV_Prot_S_129‐143 | CoV_Prot_S_121‐175 | KVCEFQFCNDPFLGV | 1 | HLA‐C07:02 | ||||||
| CoV_Prot_S_129‐143 | CoV_Prot_S_121‐175 | KVCEFQFCNDPFLGV | 1 | DRB1_1101 | ||||||
| CoV_Prot_S_133‐147 | CoV_Prot_S_121‐175 | FQFCNDPFLGVYYHK | 3 | DRB1_1501 | DRB1_1501 | |||||
| CoV_Prot_S_138‐152 | CoV_Prot_S_121‐175 | DPFLGVYYHKNNKSW | 1 | DRB1_1501 | ||||||
| CoV_Prot_S_150‐164 | CoV_Prot_S_121‐175 | KSWMESEFRVYSSAN | 2 | HLA‐A29:02 | HLA‐A01:01 | |||||
| CoV_Prot_S_150‐164 | CoV_Prot_S_121‐175 | KSWMESEFRVYSSAN | 1 | DRB1_1101 | ||||||
| CoV_Prot_S_154‐168 | CoV_Prot_S_121‐175 | ESEFRVYSSANNCTF | 1 | HLA‐C16:01 | ||||||
| CoV_Prot_S_157‐171 | CoV_Prot_S_121‐175 | FRVYSSANNCTFEYV | 1 | HLA‐B35:01 | ||||||
| CoV_Prot_S_157‐171 | CoV_Prot_S_121‐175 | FRVYSSANNCTFEYV | 1 | DRB1_1501 | ||||||
| CoV_Prot_S_161‐175 | CoV_Prot_S_121‐175 | SSANNCTFEYVSQPF | 1 | HLA‐C04:01 | ||||||
| CoV_Prot_S_165‐179 | CoV_Prot_S_165‐216 | NCTFEYVSQPFLMDL | 3 | HLA‐B40:01 | HLA‐C04:01 | HLA‐B40:02 | ||||
| CoV_Prot_S_165‐179 | CoV_Prot_S_165‐216 | NCTFEYVSQPFLMDL | 2 | DRB1_1601 | DRB1_1501 | |||||
| CoV_Prot_S_170‐184 | CoV_Prot_S_165‐216 | YVSQPFLMDLEGKQG | 1 | HLA‐C02:02 | ||||||
| CoV_Prot_S_178‐192 | CoV_Prot_S_165‐216 | DLEGKQGNFKNLREF | 1 | HLA‐B15:01 | ||||||
| CoV_Prot_S_178‐192 | CoV_Prot_S_165‐216 | DLEGKQGNFKNLREF | 1 | DQA10102‐DQB10502 | ||||||
| CoV_Prot_S_189‐203 | CoV_Prot_S_165‐216 | LREFVFKNIDGYFKI | 2 | DRB1_1501 | ||||||
| CoV_Prot_S_194‐208 | CoV_Prot_S_165‐216 | FKNIDGYFKIYSKHT | 2 | DRB1_1501 | ||||||
| CoV_Prot_S_198‐212 | CoV_Prot_S_165‐216 | DGYFKIYSKHTPINL | 2 | DRB1_1501 | ||||||
| CoV_Prot_S_202‐216 | CoV_Prot_S_165‐216 | KIYSKHTPINLVRDL | 2 | DRB1_1501 | ||||||
| CoV_Prot_S_229‐243 | CoV_Prot_S_205‐255 | LPIGINITRFQTLLA | 1 | HLA‐B08:01 | ||||||
| CoV_Prot_S_233‐247 | CoV_Prot_S_205‐255 | INITRFQTLLALHRS | 1 | DQA10501‐DQB10501 | ||||||
| CoV_Prot_S_236‐250 | CoV_Prot_S_205‐255 | TRFQTLLALHRSYLT | 1 | DRB1_0101 | ||||||
| CoV_Prot_S_241‐255 | CoV_Prot_S_205‐255 | LLALHRSYLTPGDSS | 1 | DRB1_1501 | ||||||
| CoV_Prot_S_245‐259 | CoV_Prot_S_245‐295 | HRSYLTPGDSSSGWT | 1 | DQA10501‐DQB10301 | ||||||
| CoV_Prot_S_249‐263 | CoV_Prot_S_245‐295 | LTPGDSSSGWTAGAA | 1 | HLA‐A01:01 | ||||||
| CoV_Prot_S_253‐267 | CoV_Prot_S_245‐295 | DSSSGWTAGAAAYYV | 1 | HLA‐A01:01 | ||||||
| CoV_Prot_S_257‐271 | CoV_Prot_S_245‐295 | GWTAGAAAYYVGYLQ | 1 | HLA‐A01:01 | ||||||
| CoV_Prot_S_261‐275 | CoV_Prot_S_245‐295 | GAAAYYVGYLQPRTF | 1 | HLA‐C03:03 | ||||||
| CoV_Prot_S_265‐279 | CoV_Prot_S_245‐295 | YYVGYLQPRTFLLKY | 3 | HLA‐A02:01 | ||||||
| CoV_Prot_S_265‐279 | CoV_Prot_S_245‐295 | YYVGYLQPRTFLLKY | 1 | DRB1_1101 | ||||||
| CoV_Prot_S_269‐283 | CoV_Prot_S_245‐295 | YLQPRTFLLKYNENG | 3 | HLA‐A02:01 | ||||||
| CoV_Prot_S_269‐283 | CoV_Prot_S_245‐295 | YLQPRTFLLKYNENG | 1 | DRB1_1101 | ||||||
| CoV_Prot_S_285‐299 | CoV_Prot_S_285‐327 | ITDAVDCALDPLSET | 1 | HLA‐C05:01 | ||||||
| CoV_Prot_S_289‐303 | CoV_Prot_S_285‐327 | VDCALDPLSETKCTL | 1 | HLA‐C05:01 | ||||||
| CoV_Prot_S_304‐319 | CoV_Prot_S_285‐327 | KSFTVEKGIYQTSNFR | 1 | HLA‐C02:02 | ||||||
| CoV_Prot_S_342‐356 | CoV_Prot_S_342‐390 | FNATRFASVYAWNRK | 3 | DQA10501‐DQB10301 | DRB1_1501 | |||||
| CoV_Prot_S_345‐359 | CoV_Prot_S_342‐390 | TRFASVYAWNRKRIS | 2 | HLA‐A03:01 | HLA‐B27:05 | |||||
| CoV_Prot_S_345‐359 | CoV_Prot_S_342‐390 | TRFASVYAWNRKRIS | 7 | DRB1_1103 | DRB1_1302 | DRB1_1101 | DRB1_1301 | DRB1_1501 | DRB1_1501 | DRB1_1302 |
| CoV_Prot_S_349‐363 | CoV_Prot_S_342‐390 | SVYAWNRKRISNCVA | 5 | DRB1_1101 | DRB1_1301 | DRB1_0401 | DRB1_1302 | |||
| CoV_Prot_S_353‐367 | CoV_Prot_S_342‐390 | WNRKRISNCVADYSV | 4 | DRB1_0401 | DRB1_1501 | DRB1_1302 | ||||
| CoV_Prot_S_369‐383 | CoV_Prot_S_342‐390 | YNSASFSTFKCYGVS | 1 | HLA‐C02:02 | ||||||
| CoV_Prot_S_369‐383 | CoV_Prot_S_342‐390 | YNSASFSTFKCYGVS | 3 | DRB1_1501 | DRB1_0701 | |||||
| CoV_Prot_S_373‐387 | CoV_Prot_S_342‐390 | SFSTFKCYGVSPTKL | 5 | HLA‐A24:02 | HLA‐A03:01 | |||||
| CoV_Prot_S_373‐387 | CoV_Prot_S_342‐390 | SFSTFKCYGVSPTKL | 2 | DRB1_1501 | ||||||
| CoV_Prot_S_376‐390 | CoV_Prot_S_342‐390 | TFKCYGVSPTKLNDL | 4 | HLA‐A24:02 | HLA‐A03:01 | |||||
| CoV_Prot_S_376‐390 | CoV_Prot_S_342‐390 | TFKCYGVSPTKLNDL | 2 | DRB1_1501 | ||||||
| CoV_Prot_S_385‐399 | CoV_Prot_S_381‐431 | TKLNDLCFTNVYADS | 1 | DRB1_0701 | ||||||
| CoV_Prot_S_445‐459 | CoV_Prot_S_441‐475 | VGGNYNYLYRLFRKS | 1 | HLA‐A23:01 | ||||||
| CoV_Prot_S_445‐459 | CoV_Prot_S_441‐475 | VGGNYNYLYRLFRKS | 1 | DRB1_0301 | ||||||
| CoV_Prot_S_457‐471 | CoV_Prot_S_441‐475 | RKSNLKPFERDISTE | 1 | DRB1_0301 | ||||||
| CoV_Prot_S_457‐471 | CoV_Prot_S_441‐475 | RKSNLKPFERDISTE | 1 | DRB1_0301 | ||||||
| CoV_Prot_S_461‐475 | CoV_Prot_S_441‐475 | LKPFERDISTEIYQA | 1 | DRB1_0401 | ||||||
| CoV_Prot_S_509‐523 | CoV_Prot_S_494‐531 | RVVVLSFELLHAPAT | 1 | DRB1_0101 | ||||||
| CoV_Prot_S_553‐567 | CoV_Prot_S_521‐567 | TESNKKFLPFQQFGR | 1 | HLA‐C07:01 | ||||||
| CoV_Prot_S_553‐567 | CoV_Prot_S_521‐567 | TESNKKFLPFQQFGR | 1 | DRB1_1302 | ||||||
| CoV_Prot_S_669‐683 | CoV_Prot_S_665‐707 | GICASYQTQTNSPRR | 2 | HLA‐A11:01 | ||||||
| CoV_Prot_S_685‐699 | CoV_Prot_S_665‐707 | RSVASQSIIAYTMSL | 1 | HLA‐A02:01 | ||||||
| CoV_Prot_S_698‐712 | CoV_Prot_S_698‐747 | SLGAENSVAYSNNSI | 1 | HLA‐C14:02 | ||||||
| CoV_Prot_S_702‐716 | CoV_Prot_S_698‐747 | ENSVAYSNNSIAIPT | 1 | HLA‐B55:01 | ||||||
| CoV_Prot_S_713‐727 | CoV_Prot_S_698‐747 | AIPTNFTISVTTEIL | 1 | HLA‐B51:01 | ||||||
| CoV_Prot_S_719‐733 | CoV_Prot_S_698‐747 | TISVTTEILPVSMTK | 1 | HLA‐A11:01 | ||||||
| CoV_Prot_S_722‐736 | CoV_Prot_S_698‐747 | VTTEILPVSMTKTSV | 1 | HLA‐A11:01 | ||||||
| CoV_Prot_S_725‐739 | CoV_Prot_S_698‐747 | EILPVSMTKTSVDCT | 1 | HLA‐A11:01 | ||||||
| CoV_Prot_S_753‐765 | CoV_Prot_S_737‐779 | LLQYGSFCTQLNR | 1 | DRB1_1501 | ||||||
| CoV_Prot_S_765‐779 | CoV_Prot_S_737‐779 | RALTGIAVEQDKNTQ | 1 | DRB1_1301 | ||||||
| CoV_Prot_S_777‐791 | CoV_Prot_S_769‐813 | NTQEVFAQVKQIYKT | 2 | DRB1_1301 | DRB1_1501 | |||||
| CoV_Prot_S_781‐795 | CoV_Prot_S_769‐813 | VFAQVKQIYKTPPIK | 1 | DRB1_1501 | ||||||
| CoV_Prot_S_788‐802 | CoV_Prot_S_769‐813 | IYKTPPIKDFGGFNF | 1 | DRB1_1301 | ||||||
| CoV_Prot_S_799‐813 | CoV_Prot_S_769‐813 | GFNFSQILPDPSKPS | 1 | HLA‐B15:01 | ||||||
| CoV_Prot_S_799‐813 | CoV_Prot_S_769‐813 | GFNFSQILPDPSKPS | 1 | DRB1_1301 | ||||||
| CoV_Prot_S_802‐816 | CoV_Prot_S_802‐852 | FSQILPDPSKPSKRS | 8 | DRB1_1501 | DRB1_0401 | DRB1_1501 | DRB1_0301 | |||
| CoV_Prot_S_809‐823 | CoV_Prot_S_802‐852 | PSKPSKRSFIEDLLF | 2 | DRB1_1501 | ||||||
| CoV_Prot_S_813‐827 | CoV_Prot_S_802‐852 | SKRSFIEDLLFNKVT | 1 | HLA‐C07:01 | ||||||
| CoV_Prot_S_813‐827 | CoV_Prot_S_802‐852 | SKRSFIEDLLFNKVT | 8 | DQA10102‐DQB10501 | DRB1_0401 | DRB1_0101 | DRB1_0301 | |||
| CoV_Prot_S_817‐831 | CoV_Prot_S_802‐852 | FIEDLLFNKVTLADA | 2 | DRB1_0801 | DRB1_0401 | |||||
| CoV_Prot_S_821‐835 | CoV_Prot_S_802‐852 | LLFNKVTLADAGFIK | 1 | HLA‐B08:01 | ||||||
| CoV_Prot_S_821‐835 | CoV_Prot_S_802‐852 | LLFNKVTLADAGFIK | 2 | DRB1_0101 | DRB1_0301 | |||||
| CoV_Prot_S_825‐839 | CoV_Prot_S_802‐852 | KVTLADAGFIKQYGD | 1 | HLA‐B18:01 | ||||||
| CoV_Prot_S_825‐839 | CoV_Prot_S_802‐852 | KVTLADAGFIKQYGD | 1 | DRB1_0801 | ||||||
| CoV_Prot_S_829‐843 | CoV_Prot_S_802‐852 | ADAGFIKQYGDCLGD | 1 | DRB1_0101 | ||||||
| CoV_Prot_S_838‐852 | CoV_Prot_S_802‐852 | GDCLGDIAARDLICA | 1 | HLA‐C12:03 | ||||||
| CoV_Prot_S_838‐852 | CoV_Prot_S_802‐852 | GDCLGDIAARDLICA | 1 | DRB1_0301 | ||||||
| CoV_Prot_S_844‐858 | CoV_Prot_S_841‐891 | IAARDLICAQKFNGL | 1 | HLA‐C08:02 | ||||||
| CoV_Prot_S_865‐879 | CoV_Prot_S_841‐891 | LTDEMIAQYTSALLA | 2 | HLA‐C08:02 | HLA‐A01:01 | |||||
| CoV_Prot_S_865‐879 | CoV_Prot_S_841‐891 | LTDEMIAQYTSALLA | 1 | DRB1_1501 | ||||||
| CoV_Prot_S_869‐883 | CoV_Prot_S_841‐891 | MIAQYTSALLAGTIT | 1 | DRB1_1501 | ||||||
| CoV_Prot_S_873‐887 | CoV_Prot_S_841‐891 | YTSALLAGTITSGWT | 1 | DRB1_1302 | ||||||
| CoV_Prot_S_885‐899 | CoV_Prot_S_881‐927 | GWTFGAGAALQIPFA | 1 | HLA‐C03:03 | ||||||
| CoV_Prot_S_885‐899 | CoV_Prot_S_881‐927 | GWTFGAGAALQIPFA | 1 | DRB1_0401 | ||||||
| CoV_Prot_S_889‐902 | CoV_Prot_S_881‐927 | GAGAALQIPFAMQM | 1 | HLA‐B15:01 | ||||||
| CoV_Prot_S_892‐906 | CoV_Prot_S_881‐927 | AALQIPFAMQMAYRF | 3 | HLA‐B56:01 | HLA‐B15:01 | HLA‐C03:03 | ||||
| CoV_Prot_S_892‐906 | CoV_Prot_S_881‐927 | AALQIPFAMQMAYRF | 1 | DRB1_0101 | ||||||
| CoV_Prot_S_896‐910 | CoV_Prot_S_881‐927 | IPFAMQMAYRFNGIG | 2 | HLA‐B56:01 | HLA‐C03:03 | |||||
| CoV_Prot_S_896‐910 | CoV_Prot_S_881‐927 | IPFAMQMAYRFNGIG | 1 | DRB1_0101 | ||||||
| CoV_Prot_S_917‐931 | CoV_Prot_S_917‐964 | YENQKLIANQFNSAI | 1 | DRB1_1302 | ||||||
| CoV_Prot_S_921‐935 | CoV_Prot_S_917‐964 | KLIANQFNSAIGKIQ | 1 | DRB1_1302 | ||||||
| CoV_Prot_S_977‐991 | CoV_Prot_S_955‐991 | LNDILSRLDKVEAEV | 1 | HLA‐B51:01 | ||||||
| CoV_Prot_S_986‐1000 | CoV_Prot_S_981‐1023 | KVEAEVQIDRLITGR | 1 | HLA‐B40:01 | ||||||
| CoV_Prot_S_990‐1004 | CoV_Prot_S_981‐1023 | EVQIDRLITGRLQSL | 1 | HLA‐C03:04 | ||||||
| CoV_Prot_S_990‐1004 | CoV_Prot_S_981‐1023 | EVQIDRLITGRLQSL | 1 | DRB1_1501 | ||||||
| CoV_Prot_S_994‐1008 | CoV_Prot_S_981‐1023 | DRLITGRLQSLQTYV | 2 | HLA‐C03:04 | HLA‐B27:05 | |||||
| CoV_Prot_S_997‐1011 | CoV_Prot_S_981‐1023 | ITGRLQSLQTYVTQQ | 2 | HLA‐C15:05 | HLA‐B27:05 | |||||
| CoV_Prot_S_997‐1011 | CoV_Prot_S_981‐1023 | ITGRLQSLQTYVTQQ | 1 | DRB1_1501 | ||||||
| CoV_Prot_S_1000‐1014 | CoV_Prot_S_981‐1023 | RLQSLQTYVTQQLIR | 1 | DRB1_1501 | ||||||
| CoV_Prot_S_1055‐1068 | CoV_Prot_S_1051‐1099 | SAPHGVVFLHVTYV | 1 | DRB1_1201 | ||||||
| CoV_Prot_S_1056‐1070 | CoV_Prot_S_1051‐1099 | APHGVVFLHVTYVPA | 2 | DRB1_1201 | DRB1_1501 | |||||
| CoV_Prot_S_1097‐1111 | CoV_Prot_S_1089‐1135 | SNGTHWFVTQRNFYE | 1 | DRB1_0405 | ||||||
| CoV_Prot_S_1101‐1115 | CoV_Prot_S_1089‐1135 | HWFVTQRNFYEPQII | 1 | DRB1_0405 | ||||||
| CoV_Prot_S_1117‐1131 | CoV_Prot_S_1089‐1135 | TDNTFVSGNCDVVIG | 1 | HLA‐C05:01 | ||||||
| CoV_Prot_S_1141‐1155 | CoV_Prot_S_1123‐1166 | LQPELDSFKEELDKY | 1 | DRB1_1601 | ||||||
| CoV_Prot_S_1149‐1163 | CoV_Prot_S_1123‐1166 | KEELDKYFKNHTSPD | 1 | HLA‐B40:01 | ||||||
| CoV_Prot_S_1203‐1215 | CoV_Prot_S_1195‐1230 | LGKYEQYIKWPWY | 1 | HLA‐B44:02 | ||||||
| CoV_Prot_S_1205‐1219 | CoV_Prot_S_1195‐1230 | KYEQYIKWPWYIWLG | 2 | HLA‐B44:02 | HLA‐A23:01 | |||||
| CoV_Prot_S_1209‐1223 | CoV_Prot_S_1195‐1230 | YIKWPWYIWLGFIAG | 2 | HLA‐A02:01 | HLA‐B35:01 | |||||
| CoV_Prot_S_1214‐1228 | CoV_Prot_S_1195‐1230 | WYIWLGFIAGLIAIV | 1 | HLA‐A02:01 | ||||||
| CoV_Prot_S_1215‐1225 | CoV_Prot_S_1195‐1230 | YIWLGFIAGLI | 1 | HLA‐A02:01 | ||||||
| CoV_Prot_S_1217‐1231 | CoV_Prot_S_1195‐1230 | WLGFIAGLIAIVMVT | 1 | HLA‐A02:01 | ||||||
| CoV_Prot_S_1218‐1228 | CoV_Prot_S_1195‐1230 | LGFIAGLIAIV | 1 | HLA‐A02:01 | ||||||
| CoV_Prot_S_1220‐1230 | CoV_Prot_S_1195‐1230 | FIAGLIAIVMV | 1 | HLA‐A02:01 | ||||||
| CoV_Prot_Nuc_37‐51 | CoV_Prot_Nuc_1‐51 | SKQRRPQGLPNNTAS | 1 | DRB1_1501 | ||||||
| CoV_Prot_Nuc_49‐61 | CoV_Prot_Nuc_40‐87 | TASWFTALTQHGK | 1 | DQA10301‐DQB10201 | ||||||
| CoV_Prot_Nuc_73‐87 | CoV_Prot_Nuc_40‐87 | PINTNSSPDDQIGYY | 1 | DQA10501‐DQB10201 | ||||||
| CoV_Prot_N_102‐116 | CoV_Prot_Nuc_78‐127 | KDLSPRWYFYYLGTG | 1 | HLA‐B07:02 | ||||||
| CoV_Prot_N_105‐119 | CoV_Prot_Nuc_78‐127 | SPRWYFYYLGTGPEA | 1 | HLA‐B07:02 | ||||||
| CoV_Prot_Nuc_339‐353 | CoV_Prot_Nuc_339‐387 | LDDKDPNFKDQVILL | 1 | DQA10201‐DQB10201 | ||||||
| CoV_Prot_Nuc_341‐355 | CoV_Prot_Nuc_339‐387 | DKDPNFKDQVILLNK | 2 | DRB1_0408 | DQA10201‐DQB10201 | |||||
| CoV_Prot_N_360‐374 | CoV_Prot_Nuc_339‐387 | IDAYKTFPPTEPKKD | 1 | HLA‐A30:01 | ||||||
| CoV_Prot_N_405‐419 | CoV_Prot_Nuc_377‐419 | KQLQQSMSSADSTQA | 1 | HLA‐A01:01 | ||||||
| CoV_Prot_M_9‐23 | CoV_Prot_M_1‐47 | TVEELKKLLEQWNLV | 1 | HLA‐C01:02 | ||||||
| CoV_Prot_M_9‐23 | CoV_Prot_M_1‐47 | TVEELKKLLEQWNLV | 1 | DRB1_1101 | ||||||
| CoV_Prot_M_12‐26 | CoV_Prot_M_1‐47 | ELKKLLEQWNLVIGF | 2 | HLA‐A02:01 | HLA‐A25:01 | |||||
| CoV_Prot_M_21‐31 | CoV_Prot_M_1‐47 | NLVIGFLFLTW | 1 | HLA‐C01:02 | ||||||
| CoV_Prot_M_21‐31 | CoV_Prot_M_1‐47 | NLVIGFLFLTW | 1 | DQA10101‐DQB10501 | ||||||
| CoV_Prot_M_27‐37 | CoV_Prot_M_1‐47 | LFLTWICLLQF | 1 | HLA‐C02:02 | ||||||
| CoV_Prot_M_27‐37 | CoV_Prot_M_1‐47 | LFLTWICLLQF | 1 | DQA10101‐DQB10501 | ||||||
| CoV_Prot_M_42‐56 | CoV_Prot_M_37‐73 | RNRFLYIIKLIFLWL | 1 | HLA‐A23:01 | ||||||
| CoV_Prot_M_47‐57 | CoV_Prot_M_37‐73 | YIIKLIFLWLL | 1 | DRB1_1101 | ||||||
| CoV_Prot_M_60‐70 | CoV_Prot_M_37‐73 | VTLACFVLAAV | 1 | HLA‐A02:01 | ||||||
| CoV_Prot_M_137‐151 | CoV_Prot_M_135‐183 | ELVIGAVILRGHLRI | 2 | HLA‐A68:01 | HLA‐A03:01 | |||||
| CoV_Prot_M_144‐158 | CoV_Prot_M_135‐183 | ILRGHLRIAGHHLGR | 1 | HLA‐B08:01 | ||||||
| CoV_Prot_M_149‐163 | CoV_Prot_M_135‐183 | LRIAGHHLGRCDIKD | 1 | HLA‐A03:01 | ||||||
| CoV_Prot_M_149‐163 | CoV_Prot_M_135‐183 | LRIAGHHLGRCDIKD | 3 | DRB1_1301 | DQA10201‐DQB10303 | |||||
| CoV_Prot_M_157‐171 | CoV_Prot_M_135‐183 | GRCDIKDLPKEITVA | 2 | DRB1_0405 | ||||||
| CoV_Prot_M_165‐179 | CoV_Prot_M_135‐183 | PKEITVATSRTLSYY | 1 | HLA‐C02:02 | ||||||
| CoV_Prot_M_169‐183 | CoV_Prot_M_173‐222 | TVATSRTLSYYKLGA | 1 | DQA10201‐DQB10302 | ||||||
| CoV_Prot_M_173‐187 | CoV_Prot_M_173‐222 | SRTLSYYKLGASQRV | 5 | DRB1_1501 | DRB1_0701 | DQA10301‐DQB10301 | ||||
| CoV_Prot_M_177‐191 | CoV_Prot_M_173‐222 | SYYKLGASQRVAGDS | 4 | DRB1_0701 | DQA10301‐DQB10301 | |||||
| CoV_Prot_M_193‐207 | CoV_Prot_M_173‐222 | FAAYSRYRIGNYKLN | 3 | HLA‐C02:02 | HLA‐C14:02 | HLA‐A30:01 | ||||
| CoV_Prot_M_197‐211 | CoV_Prot_M_173‐222 | SRYRIGNYKLNTDHS | 1 | HLA‐A30:01 | ||||||
| CoV_Prot_M_197‐211 | CoV_Prot_M_173‐222 | SRYRIGNYKLNTDHS | 1 | DRB1_1501 | ||||||
| CoV_Prot_M_201‐215 | CoV_Prot_M_173‐222 | IGNYKLNTDHSSSSD | 3 | DRB1_0405 | DRB1_0408 | DRB1_0401 | ||||
| CoV_Prot_M_208‐222 | CoV_Prot_M_173‐222 | TDHSSSSDNIALLVQ | 1 | HLA‐A01:01 | ||||||
| CoV_Prot_M_208‐222 | CoV_Prot_M_173‐222 | TDHSSSSDNIALLVQ | 1 | DRB1_0401 | ||||||
| 15‐mer peptide | Ancestor peptide pool | Peptide sequence | Predicted HLA restriction | ||||||
|---|---|---|---|---|---|---|---|---|---|
| CoV_Prot_S_13‐27 | CoV_Prot_S_1‐51 | SQCVNLTTRTQLPPA | DRB1_1401 | ||||||
| CoV_Prot_S_18‐32 | CoV_Prot_S_1‐51 | LTTRTQLPPAYTNSF | HLA‐C07:02 | HLA‐B35:03 | HLA‐C07:01 | ||||
| CoV_Prot_S_21‐35 | CoV_Prot_S_1‐51 | RTQLPPAYTNSFTRG | HLA‐B35:03 | ||||||
| CoV_Prot_S_24‐38 | CoV_Prot_S_1‐51 | LPPAYTNSFTRGVYY | HLA‐B35:03 | HLA‐C12:03 | HLA‐A29:02 | HLA‐C15:02 | |||
| CoV_Prot_S_29‐43 | CoV_Prot_S_1‐51 | TNSFTRGVYYPDKVF | HLA‐C12:03 | HLA‐A29:02 | |||||
| CoV_Prot_S_33‐47 | CoV_Prot_S_1‐51 | TRGVYYPDKVFRSSV | HLA‐A31:01 | DRB1_0101 | DRB1_1302 | DRB1_1101 | DRB1_0301 | ||
| CoV_Prot_S_37‐51 | CoV_Prot_S_1‐51 | YYPDKVFRSSVLHST | HLA‐B14:01 | DRB1_1501 | |||||
| CoV_Prot_S_41‐55 | CoV_Prot_S_41‐91 | KVFRSSVLHSTQDLF | HLA‐A03:01 | ||||||
| CoV_Prot_S_44‐58 | CoV_Prot_S_41‐91 | PFFSNVTWFHAIHVS | DRB1_1501 | ||||||
| CoV_Prot_S_65‐79 | CoV_Prot_S_41‐91 | FHAIHVSGTNGTKRF | DRB1_1302 | ||||||
| CoV_Prot_S_72‐86 | CoV_Prot_S_41‐91 | GTNGTKRFDNPVLPF | HLA‐C04:01 | HLA‐C07:02 | |||||
| CoV_Prot_S_77‐91 | CoV_Prot_S_41‐91 | KRFDNPVLPFNDGVY | HLA‐C04:01 | HLA‐C07:02 | |||||
| CoV_Prot_S_81‐95 | CoV_Prot_S_81‐130 | NPVLPFNDGVYFAST | HLA‐B35:03 | DRB1_1501 | |||||
| CoV_Prot_S_85‐99 | CoV_Prot_S_81‐130 | PFNDGVYFASTEKSN | HLA‐A03:01 | HLA‐DQA10401‐DQB10601 | HLA‐DQA10401‐DQB10402 | HLA‐DQA10103‐DQB10603 | DRB1_0404 | ||
| CoV_Prot_S_89‐103 | CoV_Prot_S_81‐130 | GVYFASTEKSNIIRG | HLA‐A03:01 | ||||||
| CoV_Prot_S_93‐107 | CoV_Prot_S_81‐130 | ASTEKSNIIRGWIFG | HLA‐C15:02 | ||||||
| CoV_Prot_S_105‐119 | CoV_Prot_S_81‐130 | IFGTTLDSKTQSLLI | HLA‐A02:01 | HLA‐C07:04 | HLA‐C04:01 | HLA‐B08:01 | DRB1_0301 | ||
| CoV_Prot_S_109‐123 | CoV_Prot_S_81‐130 | TLDSKTQSLLIVNNA | HLA‐A02:01 | HLA‐C07:04 | HLA‐C04:01 | HLA‐B08:01 | |||
| CoV_Prot_S_121‐135 | CoV_Prot_S_121‐175 | NNATNVVIKVCEFQF | HLA‐C12:03 | HLA‐C12:03 | HLA‐B51:01 | ||||
| CoV_Prot_S_138‐152 | CoV_Prot_S_121‐175 | DPFLGVYYHKNNKSW | HLA‐A03:01 | HLA‐A24:02 | HLA‐DQA10103‐DQB10402 | DRB1_1301 | |||
| CoV_Prot_S_150‐164 | CoV_Prot_S_121‐175 | KSWMESEFRVYSSAN | HLA‐B50:01 | DRB1_0301 | DRB1_1101 | ||||
| CoV_Prot_S_154‐168 | CoV_Prot_S_121‐175 | ESEFRVYSSANNCTF | HLA‐B50:01 | DRB1_1501 | DRB1_0404 | ||||
| CoV_Prot_S_157‐171 | CoV_Prot_S_121‐175 | FRVYSSANNCTFEYV | HLA‐C16:01 | HLA‐A01:01 | |||||
| CoV_Prot_S_161‐175 | CoV_Prot_S_121‐175 | SSANNCTFEYVSQPF | HLA‐A01:01 | ||||||
| CoV_Prot_S_165‐179 | CoV_Prot_S_165‐216 | NCTFEYVSQPFLMDL | HLA‐C07:02 | ||||||
| CoV_Prot_S_181‐195 | CoV_Prot_S_165‐216 | GKQGNFKNLREFVFK | DRB1_1201 | ||||||
| CoV_Prot_S_198‐212 | CoV_Prot_S_165‐216 | DGYFKIYSKHTPINL | HLA‐C03:03 | HLA‐C07:04 | DRB1_0801 | HLA‐DQA10301‐DQB10503 | DRB1_1101 | ||
| CoV_Prot_S_202‐216 | CoV_Prot_S_165‐216 | KIYSKHTPINLVRDL | HLA‐B07:02 | HLA‐C03:03 | HLA‐C07:04 | ||||
| CoV_Prot_S_205‐219 | CoV_Prot_S_205‐255 | SKHTPINLVRDLPQG | HLA‐DQA10103‐DQB10603 | HLA‐DQA10101‐DQB10603 | HLA‐DQA10101‐DQB10301 | HLA‐DQA10101‐DQB10501 | |||
| CoV_Prot_S_217‐231 | CoV_Prot_S_205‐255 | PQGFSALEPLVDLPI | HLA‐C03:03 | ||||||
| CoV_Prot_S_221‐235 | CoV_Prot_S_205‐255 | SALEPLVDLPIGINI | HLA‐C03:03 | ||||||
| CoV_Prot_S_233‐247 | CoV_Prot_S_205‐255 | INITRFQTLLALHRS | HLA‐C07:01 | ||||||
| CoV_Prot_S_236‐250 | CoV_Prot_S_205‐255 | TRFQTLLALHRSYLT | HLA‐C07:01 | ||||||
| CoV_Prot_S_245‐259 | CoV_Prot_S_245‐295 | HRSYLTPGDSSSGWT | HLA‐A02:01 | ||||||
| CoV_Prot_S_249‐263 | CoV_Prot_S_245‐295 | LTPGDSSSGWTAGAA | HLA‐C12:03 | ||||||
| CoV_Prot_S_253‐267 | CoV_Prot_S_245‐295 | DSSSGWTAGAAAYYV | HLA‐DQA10501‐DQB10301 | ||||||
| CoV_Prot_S_257‐271 | CoV_Prot_S_245‐295 | GWTAGAAAYYVGYLQ | HLA‐DQA10501‐DQB10301 | ||||||
| CoV_Prot_S_265‐279 | CoV_Prot_S_245‐295 | YYVGYLQPRTFLLKY | HLA‐A02:01 | ||||||
| CoV_Prot_S_269‐283 | CoV_Prot_S_245‐295 | YLQPRTFLLKYNENG | HLA‐A02:01 | ||||||
| CoV_Prot_S_273‐287 | CoV_Prot_S_245‐295 | RTFLLKYNENGTITD | DRB1_0301 | ||||||
| CoV_Prot_S_302‐316 | CoV_Prot_S_285‐327 | TLKSFTVEKGIYQTS | DRB1_1101 | ||||||
| CoV_Prot_S_322‐335 | CoV_Prot_S_313‐351 | PTESIVRFPNITNL | HLA‐C07:02 | ||||||
| CoV_Prot_S_323‐335 | CoV_Prot_S_313‐351 | TESIVRFPNITNL | HLA‐C07:02 | ||||||
| CoV_Prot_S_324‐338 | CoV_Prot_S_313‐351 | ESIVRFPNITNLCPF | HLA‐C07:02 | ||||||
| CoV_Prot_S_326‐338 | CoV_Prot_S_313‐351 | IVRFPNITNLCPF | HLA‐C07:02 | ||||||
| CoV_Prot_S_342‐356 | CoV_Prot_S_342‐390 | FNATRFASVYAWNRK | HLA‐C07:01 | HLA‐B27:05 | HLA‐DQA10401‐DQB10402 | HLA‐DQA10103‐DQB10402 | |||
| CoV_Prot_S_345‐359 | CoV_Prot_S_342‐390 | TRFASVYAWNRKRIS | HLA‐A03:01 | HLA‐A74:03 | DRB1_1301 | ||||
| CoV_Prot_S_349‐363 | CoV_Prot_S_342‐390 | SVYAWNRKRISNCVA | HLA‐A03:01 | HLA‐A74:03 | |||||
| CoV_Prot_S_357‐371 | CoV_Prot_S_342‐390 | RISNCVADYSVLYNS | HLA‐A01:01 | ||||||
| CoV_Prot_S_362‐376 | CoV_Prot_S_313‐351 | VADYSVLYNSASFST | DRB1_0401 | DRB1_0101 | DRB1_1101 | ||||
| CoV_Prot_S_364‐378 | CoV_Prot_S_342‐390 | DYSVLYNSASFSTFK | HLA‐A11:01 | DRB1_1302 | |||||
| CoV_Prot_S_369‐383 | CoV_Prot_S_342‐390 | YNSASFSTFKCYGVS | HLA‐A11:01 | DRB1_1501 | |||||
| CoV_Prot_S_373‐387 | CoV_Prot_S_342‐390 | SFSTFKCYGVSPTKL | HLA‐A03:01 | ||||||
| CoV_Prot_S_376‐390 | CoV_Prot_S_342‐390 | TFKCYGVSPTKLNDL | HLA‐A03:01 | ||||||
| CoV_Prot_S_397‐411 | CoV_Prot_S_381‐431 | ADSFVIRGDEVRQIA | HLA‐C06:02 | ||||||
| CoV_Prot_S_401‐415 | CoV_Prot_S_381‐431 | VIRGDEVRQIAPGQT | HLA‐C06:02 | ||||||
| CoV_Prot_S_405‐419 | CoV_Prot_S_381‐431 | DEVRQIAPGQTGKIA | HLA‐A03:01 | ||||||
| CoV_Prot_S_413‐427 | CoV_Prot_S_381‐431 | GQTGKIADYNYKLPD | HLA‐A02:01 | ||||||
| CoV_Prot_S_417‐431 | CoV_Prot_S_381‐431 | KIADYNYKLPDDFTG | HLA‐A02:01 | ||||||
| CoV_Prot_S_430‐444 | CoV_Prot_S_421‐455 | TGCVIAWNSNNLDSK | HLA‐DQA10103‐DQB10501 | HLA‐DQA10101‐DQB10501 | DRB1_1501 | ||||
| CoV_Prot_S_438‐452 | CoV_Prot_S_421‐455 | SNNLDSKVGGNYNYL | HLA‐C07:02 | ||||||
| CoV_Prot_S_441‐455 | CoV_Prot_S_421‐455 | LDSKVGGNYNYLYRL | HLA‐C07:02 | ||||||
| CoV_Prot_S_445‐459 | CoV_Prot_S_441‐475 | VGGNYNYLYRLFRKS | DRB1_1101 | ||||||
| CoV_Prot_S_453‐467 | CoV_Prot_S_441‐475 | YRLFRKSNLKPFERD | HLA‐A31:01 | ||||||
| CoV_Prot_S_461‐475 | CoV_Prot_S_441‐475 | LKPFERDISTEIYQA | HLA‐B52:01 | HLA‐B50:01 | DRB1_0101 | HLA‐B52:01 | HLA‐B56:01 | ||
| CoV_Prot_S_464‐478 | CoV_Prot_S_461‐508 | FERDISTEIYQAGST | HLA‐B52:01 | ||||||
| CoV_Prot_S_486‐500 | CoV_Prot_S_461‐508 | FNCYFPLQSYGFQPT | HLA‐C07:02 | ||||||
| CoV_Prot_S_489‐503 | CoV_Prot_S_461‐508 | YFPLQSYGFQPTNGV | HLA‐C07:02 | ||||||
| CoV_Prot_S_494‐508 | CoV_Prot_S_461‐508 | SYGFQPTNGVGYQPY | HLA‐B15:01 | DRB1_1302 | |||||
| CoV_Prot_S_500‐513 | CoV_Prot_S_494‐531 | TNGVGYQPYRVVVL | HLA‐C01:02 | ||||||
| CoV_Prot_S_501‐515 | CoV_Prot_S_494‐531 | NGVGYQPYRVVVLSF | HLA‐C01:02 | ||||||
| CoV_Prot_S_502‐515 | CoV_Prot_S_494‐531 | GVGYQPYRVVVLSF | HLA‐C01:02 | ||||||
| CoV_Prot_S_505‐519 | CoV_Prot_S_494‐531 | YQPYRVVVLSFELLH | HLA‐B52:01 | HLA‐C01:02 | HLA‐C01:02 | HLA‐B08:01 | |||
| CoV_Prot_S_521‐535 | CoV_Prot_S_521‐567 | PATVCGPKKSTNLVK | HLA‐C01:02 | ||||||
| CoV_Prot_S_526‐539 | CoV_Prot_S_521‐567 | GPKKSTNLVKNKCV | HLA‐A03:01 | ||||||
| CoV_Prot_S_529‐543 | CoV_Prot_S_521‐567 | KSTNLVKNKCVNFNF | HLA‐A03:01 | DRB1_1302 | |||||
| CoV_Prot_S_541‐555 | CoV_Prot_S_521‐567 | FNFNGLTGTGVLTES | DRB1_0101 | ||||||
| CoV_Prot_S_543‐557 | CoV_Prot_S_521‐567 | FNGLTGTGVLTESNK | HLA‐DQA10103‐DQB10601 | ||||||
| CoV_Prot_S_553‐567 | CoV_Prot_S_521‐567 | TESNKKFLPFQQFGR | DRB1_1501 | ||||||
| CoV_Prot_S_557‐571 | CoV_Prot_S_557‐607 | KKFLPFQQFGRDIAD | DRB1_1501 | ||||||
| CoV_Prot_S_573‐587 | CoV_Prot_S_557‐607 | TDAVRDPQTLEILDI | HLA‐C07:02 | ||||||
| CoV_Prot_S_581‐595 | CoV_Prot_S_557‐607 | TLEILDITPCSFGGV | HLA‐C05:01 | ||||||
| CoV_Prot_S_621‐635 | CoV_Prot_S_597‐635 | PVAIHADQLTPTWRV | DRB1_0301 | ||||||
| CoV_Prot_S_656‐670 | CoV_Prot_S_597‐635 | VNNSYECDIPIGAGI | HLA‐B50:01 | DRB1_0301 | DRB1_0401 | ||||
| CoV_Prot_S_660‐674 | CoV_Prot_S_597‐635 | YECDIPIGAGICASY | HLA‐B50:01 | HLA‐B56:01 | |||||
| CoV_Prot_S_673‐687 | CoV_Prot_S_665‐707 | SYQTQTNSPRRARSV | HLA‐A68:01 | ||||||
| CoV_Prot_S_678‐692 | CoV_Prot_S_625‐674 | TNSPRRARSVASQSI | HLA‐B07:02 | HLA‐B07:02 | HLA‐B56:01 | ||||
| CoV_Prot_S_683‐696 | CoV_Prot_S_625‐674 | RARSVASQSIIAYT | HLA‐C12:02 | HLA‐C12:03 | DRB1_0301 | ||||
| CoV_Prot_S_685‐699 | CoV_Prot_S_625‐674 | RSVASQSIIAYTMSL | HLA‐C12:02 | HLA‐C12:03 | |||||
| CoV_Prot_S_689‐703 | CoV_Prot_S_665‐707 | SQSIIAYTMSLGAEN | HLA‐DQA10102‐DQB10503 | ||||||
| CoV_Prot_S_702‐716 | CoV_Prot_S_698‐747 | ENSVAYSNNSIAIPT | HLA‐DQA10103‐DQB10402 | HLA‐DQA10103‐DQB10601 | HLA‐DQA10102‐DQB10503 | DRB1_1501 | |||
| CoV_Prot_S_706‐720 | CoV_Prot_S_698‐747 | AYSNNSIAIPTNFTI | HLA‐B52:01 | ||||||
| CoV_Prot_S_709‐723 | CoV_Prot_S_698‐747 | NNSIAIPTNFTISVT | HLA‐B52:01 | HLA‐B55:01 | HLA‐B51:01 | ||||
| CoV_Prot_S_713‐727 | CoV_Prot_S_698‐747 | AIPTNFTISVTTEIL | HLA‐C05:01 | HLA‐B51:01 | |||||
| CoV_Prot_S_719‐733 | CoV_Prot_S_698‐747 | TISVTTEILPVSMTK | HLA‐A03:01 | HLA‐DQA10501‐DQB10201 | |||||
| CoV_Prot_S_722‐736 | CoV_Prot_S_698‐747 | VTTEILPVSMTKTSV | HLA‐A03:01 | ||||||
| CoV_Prot_S_725‐739 | CoV_Prot_S_698‐747 | EILPVSMTKTSVDCT | HLA‐A03:01 | ||||||
| CoV_Prot_S_745‐759 | CoV_Prot_S_737‐779 | DSTECSNLLLQYGSF | HLA‐C05:01 | ||||||
| CoV_Prot_S_749‐763 | CoV_Prot_S_737‐779 | CSNLLLQYGSFCTQL | HLA‐C14:02 | DRB1_1501 | |||||
| CoV_Prot_S_753‐765 | CoV_Prot_S_737‐779 | LLQYGSFCTQLNR | HLA‐A11:01 | HLA‐C14:02 | |||||
| CoV_Prot_S_756‐767 | CoV_Prot_S_737‐779 | YGSFCTQLNRAL | HLA‐A11:01 | ||||||
| CoV_Prot_S_757‐770 | CoV_Prot_S_737‐779 | GSFCTQLNRALTGI | HLA‐A11:01 | ||||||
| CoV_Prot_S_765‐779 | CoV_Prot_S_737‐779 | RALTGIAVEQDKNTQ | HLA‐DQA10401‐DQB10402 | HLA‐DQA10501‐DQB10201 | HLA‐DQA10102‐DQB10602 | HLA‐DQA10501‐DQB10602 | |||
| CoV_Prot_S_769‐783 | CoV_Prot_S_769‐813 | GIAVEQDKNTQEVFA | HLA‐B38:01 | DRB1_0301 | |||||
| CoV_Prot_S_773‐787 | CoV_Prot_S_769‐813 | EQDKNTQEVFAQVKQ | HLA‐B38:01 | ||||||
| CoV_Prot_S_777‐791 | CoV_Prot_S_769‐813 | NTQEVFAQVKQIYKT | HLA‐A29:01 | HLA‐C14:02 | DRB1_1401 | ||||
| CoV_Prot_S_781‐795 | CoV_Prot_S_769‐813 | VFAQVKQIYKTPPIK | HLA‐A11:01 | HLA‐A03:01 | HLA‐C14:02 | ||||
| CoV_Prot_S_785‐799 | CoV_Prot_S_769‐813 | VKQIYKTPPIKDFGG | HLA‐A11:01 | HLA‐A03:01 | HLA‐C07:01 | ||||
| CoV_Prot_S_788‐802 | CoV_Prot_S_769‐813 | IYKTPPIKDFGGFNF | HLA‐C07:01 | ||||||
| CoV_Prot_S_792‐806 | CoV_Prot_S_769‐813 | PPIKDFGGFNFSQIL | DRB1_1501 | ||||||
| CoV_Prot_S_796‐810 | CoV_Prot_S_769‐813 | DFGGFNFSQILPDPS | HLA‐DQA10401‐DQB10402 | ||||||
| CoV_Prot_S_802‐816 | CoV_Prot_S_802‐852 | FSQILPDPSKPSKRS | HLA‐DQA10301‐DQB10503 | DRB1_1302 | DRB1_0401 | DRB1_1301 | DRB1_1601 | ||
| CoV_Prot_S_817‐831 | CoV_Prot_S_802‐852 | FIEDLLFNKVTLADA | HLA‐A02:01 | HLA‐B08:01 | |||||
| CoV_Prot_S_821‐835 | CoV_Prot_S_802‐852 | LLFNKVTLADAGFIK | HLA‐A02:01 | ||||||
| CoV_Prot_S_825‐839 | CoV_Prot_S_802‐852 | KVTLADAGFIKQYGD | HLA‐B44:02 | ||||||
| CoV_Prot_S_829‐843 | CoV_Prot_S_802‐852 | ADAGFIKQYGDCLGD | HLA‐B44:02 | HLA‐DQA10103‐DQB10501 | DRB1_1501 | ||||
| CoV_Prot_S_841‐855 | CoV_Prot_S_841‐891 | LGDIAARDLICAQKF | HLA‐C05:01 | ||||||
| CoV_Prot_S_847‐861 | CoV_Prot_S_841‐891 | RDLICAQKFNGLTVL | HLA‐B14:02 | ||||||
| CoV_Prot_S_852‐866 | CoV_Prot_S_841‐891 | AQKFNGLTVLPPLLT | HLA‐B14:02 | ||||||
| CoV_Prot_S_860‐874 | CoV_Prot_S_841‐891 | VLPPLLTDEMIAQYT | DRB1_0301 | ||||||
| CoV_Prot_S_865‐879 | CoV_Prot_S_841‐891 | LTDEMIAQYTSALLA | HLA‐B50:01 | HLA‐C03:04 | HLA‐DQA10101‐DQB10501 | DRB1_1501 | |||
| CoV_Prot_S_869‐883 | CoV_Prot_S_841‐891 | MIAQYTSALLAGTIT | HLA‐B08:01 | HLA‐B50:01 | HLA‐C03:04 | HLA‐DQA10401‐DQB10601 | HLA‐DQA10102‐DQB10602 | ||
| CoV_Prot_S_881‐895 | CoV_Prot_S_881‐927 | TITSGWTFGAGAALQ | HLA‐C03:04 | HLA‐C03:03 | |||||
| CoV_Prot_S_885‐899 | CoV_Prot_S_881‐927 | GWTFGAGAALQIPFA | HLA‐C03:04 | HLA‐C03:03 | HLA‐DQA10501‐DQB10301 | DRB1_0101 | |||
| CoV_Prot_S_889‐902 | CoV_Prot_S_881‐927 | GAGAALQIPFAMQM | HLA‐B52:01 | ||||||
| CoV_Prot_S_892‐906 | CoV_Prot_S_881‐927 | AALQIPFAMQMAYRF | HLA‐B52:01 | ||||||
| CoV_Prot_S_896‐910 | CoV_Prot_S_881‐927 | IPFAMQMAYRFNGIG | DRB1_1101 | ||||||
| CoV_Prot_S_902‐915 | CoV_Prot_S_881‐927 | MAYRFNGIGVTQNV | HLA‐B39:06 | ||||||
| CoV_Prot_S_904‐918 | CoV_Prot_S_881‐927 | YRFNGIGVTQNVLYE | HLA‐B39:06 | ||||||
| CoV_Prot_S_912‐923 | CoV_Prot_S_881‐927 | TQNVLYENQKLI | HLA‐A02:01 | DRB1_1501 | |||||
| CoV_Prot_S_915‐927 | CoV_Prot_S_881‐927 | VLYENQKLIANQF | HLA‐A02:01 | HLA‐B15:01 | |||||
| CoV_Prot_S_933‐947 | CoV_Prot_S_917‐964 | KIQDSLSSTASALGK | HLA‐A11:01 | ||||||
| CoV_Prot_S_937‐951 | CoV_Prot_S_917‐964 | SLSSTASALGKLQDV | HLA‐A11:01 | ||||||
| CoV_Prot_S_945‐959 | CoV_Prot_S_917‐964 | LGKLQDVVNQNAQAL | HLA‐C08:02 | HLA‐C03:04 | |||||
| CoV_Prot_S_948‐962 | CoV_Prot_S_917‐964 | LQDVVNQNAQALNTL | HLA‐C08:02 | HLA‐C03:04 | DRB1_0301 | DRB1_1302 | |||
| CoV_Prot_S_959‐973 | CoV_Prot_S_955‐991 | LNTLVKQLSSNFGAI | DRB1_1501 | ||||||
| CoV_Prot_S_963‐976 | CoV_Prot_S_955‐991 | VKQLSSNFGAISSV | HLA‐C15:02 | ||||||
| CoV_Prot_S_967‐981 | CoV_Prot_S_955‐991 | SSNFGAISSVLNDIL | HLA‐DQA10501‐DQB10201 | ||||||
| CoV_Prot_S_972‐984 | CoV_Prot_S_955‐991 | AISSVLNDILSRL | HLA‐A11:01 | HLA‐A02:01 | |||||
| CoV_Prot_S_975‐987 | CoV_Prot_S_955‐991 | SVLNDILSRLDKV | HLA‐A11:01 | HLA‐A02:01 | |||||
| CoV_Prot_S_981‐995 | CoV_Prot_S_981‐1023 | LSRLDKVEAEVQIDR | HLA‐A02:01 | ||||||
| CoV_Prot_S_986‐1000 | CoV_Prot_S_981‐1023 | KVEAEVQIDRLITGR | DRB1_0301 | ||||||
| CoV_Prot_S_994‐1008 | CoV_Prot_S_981‐1023 | DRLITGRLQSLQTYV | HLA‐C07:02 | ||||||
| CoV_Prot_S_997‐1011 | CoV_Prot_S_981‐1023 | ITGRLQSLQTYVTQQ | HLA‐C07:02 | ||||||
| CoV_Prot_S_1000‐1014 | CoV_Prot_S_981‐1023 | RLQSLQTYVTQQLIR | HLA‐B52:01 | ||||||
| CoV_Prot_S_1004‐1018 | CoV_Prot_S_981‐1023 | LQTYVTQQLIRAAEI | HLA‐B52:01 | ||||||
| CoV_Prot_S_1009‐1023 | CoV_Prot_S_981‐1023 | TQQLIRAAEIRASAN | HLA‐DQA10103‐DQB10603 | HLA‐DQA10101‐DQB10603 | HLA‐DQA10102‐DQB10602 | HLA‐DQA10301‐DQB10602 | HLA‐DQA10102‐DQB10503 | HLA‐DQA10301‐DQB10503 | |
| CoV_Prot_S_1012‐1026 | CoV_Prot_S_1012‐1059 | LIRAAEIRASANLAA | HLA‐B39:06 | HLA‐DQA10102‐DQB10602 | |||||
| CoV_Prot_S_1014‐1028 | CoV_Prot_S_1012‐1059 | RAAEIRASANLAATK | HLA‐B39:06 | HLA‐DQA10301‐DQB10602 | HLA‐DQA10102‐DQB10503 | HLA‐DQA10301‐DQB10503 | DRB1_1501 | IRASANLAA | DRB1_0404 |
| CoV_Prot_S_1045‐1059 | CoV_Prot_S_1012‐1059 | KGYHLMSFPQSAPHG | HLA‐A02:01 | ||||||
| CoV_Prot_S_1061‐1075 | CoV_Prot_S_1051‐1099 | VFLHVTYVPAQEKNF | HLA‐A11:01 | HLA‐A03:01 | |||||
| CoV_Prot_S_1066‐1080 | CoV_Prot_S_1051‐1099 | TYVPAQEKNFTTAPA | HLA‐B45:01 | ||||||
| CoV_Prot_S_1069‐1083 | CoV_Prot_S_1051‐1099 | PAQEKNFTTAPAICH | HLA‐B45:01 | ||||||
| CoV_Prot_S_1073‐1087 | CoV_Prot_S_1051‐1099 | KNFTTAPAICHDGKA | HLA‐DQA10103‐DQB10601 | ||||||
| CoV_Prot_S_1077‐1091 | CoV_Prot_S_1051‐1099 | TAPAICHDGKAHFPR | DRB1_0301 | DRB1_1401 | |||||
| CoV_Prot_S_1093‐1107 | CoV_Prot_S_1089‐1135 | GVFVSNGTHWFVTQR | HLA‐A11:01 | HLA‐A03:01 | |||||
| CoV_Prot_S_1097‐1111 | CoV_Prot_S_1089‐1135 | SNGTHWFVTQRNFYE | HLA‐A11:01 | HLA‐A03:01 | DRB1_0301 | HLA‐DQA10102‐DQB10503 | |||
| CoV_Prot_S_1131‐1145 | CoV_Prot_S_1123‐1166 | GIVNNTVYDPLQPEL | HLA‐C04:01 | HLA‐C05:01 | HLA‐C06:02 | HLA‐C01:02 | |||
| CoV_Prot_S_1136‐1149 | CoV_Prot_S_1123‐1166 | TVYDPLQPELDSFK | HLA‐C04:01 | HLA‐C05:01 | HLA‐C06:02 | HLA‐C01:02 | |||
| CoV_Prot_S_1167‐1181 | CoV_Prot_S_1157‐1205 | GDISGINASVVNIQK | HLA‐DQA10401‐DQB10601 | HLA‐DQA10103‐DQB10601 | HLA‐DQA10501‐DQB10301 | ||||
| CoV_Prot_S_1171‐1185 | CoV_Prot_S_1157‐1205 | GINASVVNIQKEIDR | DRB1_0801 | ||||||
| CoV_Prot_S_1175‐1189 | CoV_Prot_S_1157‐1205 | SVVNIQKEIDRLNEV | HLA‐B40:01 | ||||||
| CoV_Prot_S_1179‐1193 | CoV_Prot_S_1157‐1205 | IQKEIDRLNEVAKNL | HLA‐B40:01 | ||||||
| CoV_Prot_S_1181‐1195 | CoV_Prot_S_1157‐1205 | KEIDRLNEVAKNLNE | HLA‐B40:01 | ||||||
| CoV_Prot_S_1186‐1200 | CoV_Prot_S_1157‐1205 | LNEVAKNLNESLIDL | HLA‐C12:02 | ||||||
| CoV_Prot_S_1189‐1203 | CoV_Prot_S_1157‐1205 | VAKNLNESLIDLQEL | HLA‐C12:02 | ||||||
| CoV_Prot_S_1197‐1211 | CoV_Prot_S_1195‐1230 | LIDLQELGKYEQYIK | HLA‐B44:02 | HLA‐B18:01 | DRB1_0801 | DRB1_0401 | |||
| CoV_Prot_S_1217‐1231 | CoV_Prot_S_1195‐1230 | WLGFIAGLIAIVMVT | HLA‐DQA10501‐DQB10301 | ||||||
| CoV_Prot_S_1255‐1268 | CoV_Prot_S_1222‐1273 | KFDEDDSEPVLKGV | HLA‐B38:01 | DRB1_0401 | |||||
| CoV_Prot_S_1257‐1271 | CoV_Prot_S_1222‐1273 | DEDDSEPVLKGVKLH | HLA‐B35:03 | ||||||
| CoV_Prot_S_1259‐1273 | CoV_Prot_S_1222‐1273 | DDSEPVLKGVKLHYT | HLA‐C02:02 | HLA‐B15:01 | HLA‐B35:03 | HLA‐DQA10103‐DQB10601 | |||
| CoV_Prot_Nuc_5‐19 | CoV_Prot_N_1‐51 | GPQNQRNAPRITFGG | HLA‐DQA10102‐DQB10602 | HLA‐B38:01 | |||||
| CoV_Prot_Nuc_10‐24 | CoV_Prot_N_1‐51 | RNAPRITFGGPSDST | HLA‐DQA10401‐DQB10601 | ||||||
| CoV_Prot_Nuc_13‐27 | CoV_Prot_N_1‐51 | PRITFGGPSDSTGSN | HLA‐DQA10401‐DQB10601 | ||||||
| CoV_Prot_Nuc_37‐51 | CoV_Prot_N_1‐51 | SKQRRPQGLPNNTAS | HLA‐B07:02 | ||||||
| CoV_Prot_Nuc_54‐68 | CoV_Prot_N_40‐87 | TALTQHGKEDLKFPR | HLA‐B38:01 | ||||||
| CoV_Prot_Nuc_57‐71 | CoV_Prot_N_40‐87 | TQHGKEDLKFPRGQG | HLA‐B38:01 | ||||||
| CoV_Prot_Nuc_78‐92 | CoV_Prot_N_78‐127 | SSPDDQIGYYRRATR | HLA‐C04:01 | ||||||
| CoV_Prot_Nuc_81‐95 | CoV_Prot_N_78‐127 | DDQIGYYRRATRRIR | HLA‐A24:02 | ||||||
| CoV_Prot_Nuc_85‐99 | CoV_Prot_N_78‐127 | GYYRRATRRIRGGDG | HLA‐A24:02 | ||||||
| CoV_Prot_Nuc_113‐127 | CoV_Prot_N_78‐127 | LGTGPEAGLPYGANK | HLA‐DQA10401‐DQB10601 | ||||||
| CoV_Prot_Nuc_149‐163 | CoV_Prot_N_117‐167 | RNPANNAAIVLQLPQ | HLA‐DQA10103‐DQB10603 | ||||||
| CoV_Prot_Nuc_158‐172 | CoV_Prot_N_158‐203 | VLQLPQGTTLPKGFY | HLA‐C07:02 | ||||||
| CoV_Prot_Nuc_197‐211 | CoV_Prot_N_194‐234 | STPGSSRGTSPARMA | HLA‐DQA10101‐DQB10603 | HLA‐DQA10101‐DQB10301 | |||||
| CoV_Prot_Nuc_217‐230 | CoV_Prot_N_194‐234 | AALALLLLDRLNQL | HLA‐A02:01 | DRB1_1101 | |||||
| CoV_Prot_Nuc_220‐234 | CoV_Prot_N_194‐234 | ALLLLDRLNQLESKM | HLA‐A02:01 | ||||||
| CoV_Prot_Nuc_243‐256 | CoV_Prot_N_224‐274 | GQTVTKKSAAEASK | HLA‐DQA10103‐DQB10402 | HLA‐DQA10501‐DQB10301 | |||||
| CoV_Prot_Nuc_253‐267 | CoV_Prot_N_224‐274 | EASKKPRQKRTATKA | HLA‐B07:02 | ||||||
| CoV_Prot_Nuc_257‐271 | CoV_Prot_N_224‐274 | KPRQKRTATKAYNVT | HLA‐B07:02 | ||||||
| CoV_Prot_Nuc_261‐274 | CoV_Prot_N_224‐274 | KRTATKAYNVTQAF | HLA‐C03:03 | HLA‐C06:02 | |||||
| CoV_Prot_Nuc_265‐278 | CoV_Prot_N_265‐314 | TKAYNVTQAFGRRG | HLA‐C12:02 | ||||||
| CoV_Prot_Nuc_293‐307 | CoV_Prot_N_265‐314 | RQGTDYKHWPQIAQF | HLA‐B38:01 | HLA‐C07:02 | |||||
| CoV_Prot_Nuc_297‐311 | CoV_Prot_N_265‐314 | DYKHWPQIAQFAPSA | HLA‐B38:01 | HLA‐C07:02 | |||||
| CoV_Prot_Nuc_300‐314 | CoV_Prot_N_265‐314 | HWPQIAQFAPSASAF | HLA‐DQA10103‐DQB10402 | ||||||
| CoV_Prot_Nuc_318‐331 | CoV_Prot_N_305‐347 | SRIGMEVTPSGTWL | HLA‐B44:03 | ||||||
| CoV_Prot_Nuc_321‐335 | CoV_Prot_N_305‐347 | GMEVTPSGTWLTYTG | HLA‐B44:03 | ||||||
| CoV_Prot_Nuc_333‐347 | CoV_Prot_N_305‐347 | YTGAIKLDDKDPNFK | HLA‐C04:01 | ||||||
| CoV_Prot_Nuc_357‐371 | CoV_Prot_N_339‐387 | YKTFPPTEPKKDKKK | HLA‐A30:01 | ||||||
| CoV_Prot_Nuc_357‐371 | CoV_Prot_N_339‐387 | IDAYKTFPPTEPKKD | DRB1_0408 | DRB1_0701 | |||||
| CoV_Prot_M_1‐15 | CoV_Prot_M_1‐47 | MADSNGTITVEELKK | HLA‐DQA10103‐DQB10603 | ||||||
| CoV_Prot_M_9‐23 | CoV_Prot_M_1‐47 | TVEELKKLLEQWNLV | HLA‐A02:01 | ||||||
| CoV_Prot_M_12‐26 | CoV_Prot_M_1‐47 | ELKKLLEQWNLVIGF | HLA‐B52:01 | HLA‐A02:01 | |||||
| CoV_Prot_M_14‐29 | CoV_Prot_M_1‐47 | KKLLEQWNLVIGFLFL | HLA‐B52:01 | HLA‐A02:01 | |||||
| CoV_Prot_M_34‐47 | CoV_Prot_M_1‐47 | LLQFAYANRNRFLY | HLA‐C03:03 | HLA‐C12:03 | HLA‐C02:02 | DRB1_1101 | |||
| CoV_Prot_M_37‐51 | CoV_Prot_M_37‐73 | FAYANRNRFLYIIKL | HLA‐C12:02 | HLA‐C03:03 | |||||
| CoV_Prot_M_39‐53 | CoV_Prot_M_37‐73 | YANRNRFLYIIKLIF | HLA‐A29:01 | HLA‐DQA10101‐DQB10501 | |||||
| CoV_Prot_M_42‐56 | CoV_Prot_M_37‐73 | RNRFLYIIKLIFLWL | DRB1_1101 | DRB1_0301 | |||||
| CoV_Prot_M_65‐76 | CoV_Prot_M_65‐105 | FVLAAVYRINWI | HLA‐A02:01 | ||||||
| CoV_Prot_M_72‐85 | CoV_Prot_M_65‐105 | RINWITGGIAIAMA | HLA‐DQA10501‐DQB10301 | HLA‐DQA10501‐DQB10201 | |||||
| CoV_Prot_M_89‐103 | CoV_Prot_M_65‐105 | GLMWLSYFIASFRLF | HLA‐A23:01 | ||||||
| CoV_Prot_M_93‐105 | CoV_Prot_M_65‐105 | LSYFIASFRLFAR | HLA‐A23:01 | ||||||
| CoV_Prot_M_97‐111 | CoV_Prot_M_97‐147 | IASFRLFARTRSMWS | DRB1_0801 | ||||||
| CoV_Prot_M_102‐116 | CoV_Prot_M_97‐147 | LFARTRSMWSFNPET | HLA‐C07:01 | ||||||
| CoV_Prot_M_105‐119 | CoV_Prot_M_97‐147 | RTRSMWSFNPETNIL | HLA‐C04:01 | ||||||
| CoV_Prot_M_110‐124 | CoV_Prot_M_97‐147 | WSFNPETNILLNVPL | HLA‐C04:01 | DRB1_0101 | |||||
| CoV_Prot_M_117‐131 | CoV_Prot_M_97‐147 | NILLNVPLHGTILTR | HLA‐C01:02 | ||||||
| CoV_Prot_M_120‐134 | CoV_Prot_M_97‐147 | LNVPLHGTILTRPLL | HLA‐C01:02 | ||||||
| CoV_Prot_M_129‐143 | CoV_Prot_M_97‐147 | LTRPLLESELVIGAV | HLA‐B45:01 | ||||||
| CoV_Prot_M_133‐147 | CoV_Prot_M_97‐147 | LLESELVIGAVILRG | HLA‐B45:01 | DRB1_0301 | |||||
| CoV_Prot_M_135‐148 | CoV_Prot_M_135‐183 | ESELVIGAVILRGH | HLA‐DQA10401‐DQB10601 | HLA‐DQA10102‐DQB10602 | |||||
| CoV_Prot_M_165‐179 | CoV_Prot_M_135‐183 | PKEITVATSRTLSYY | HLA‐C03:03 | HLA‐A01:01 | HLA‐DQA10103‐DQB10402 | DRB1_0101 | DRB1_0701 | ||
| CoV_Prot_M_169‐183 | CoV_Prot_M_135‐183 | TVATSRTLSYYKLGA | HLA‐C07:02 | HLA‐A01:01 | HLA‐C02:02 | ||||
| CoV_Prot_M_173‐187 | CoV_Prot_M_173‐222 | SRTLSYYKLGASQRV | HLA‐C07:02 | HLA‐C06:02 | |||||
| CoV_Prot_M_177‐191 | CoV_Prot_M_173‐222 | SYYKLGASQRVAGDS | HLA‐DQA10501‐DQB10301 | DRB1_0101 | |||||
| CoV_Prot_M_201‐215 | CoV_Prot_M_173‐222 | IGNYKLNTDHSSSSD | HLA‐DQA10103‐DQB10402 | HLA‐DQA10101‐DQB10603 | HLA‐DQA10103‐DQB10603 | HLA‐DQA10102‐DQB10201 | |||
| CoV_Prot_M_205‐219 | CoV_Prot_M_173‐222 | KLNTDHSSSSDNIAL | HLA‐DQA10101‐DQB10603 |
| PBMC | Expanded cells | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Core peptide ID | Ancestor 15mer peptide | Ancestor peptide pool | HLA‐restric‐tion | Core peptide sequence | #Tests total | #CD4+/ CD8+ res‐ponse | #CD4+ res‐ponse | #CD8+ res‐ponse | #Total postive hits | Total postive hits [%] | #Tests total | #CD4+/ CD8+ res‐ponse | #CD4+ res‐ponse | #CD8+ res‐ponse | #Total postive hits | Total postive hits [%] |
| DRB1_0101_P1 | CoV_Prot_S_821‐835 | CoV_Prot_S_802‐852 | DRB1_0101 | VTLADAGFI | 1 | 0 | 0 | 1 | 1 | 1/1 (100) | 1 | 0 | 0 | 0 | 0 | 0/1 |
| DRB1_0101_P2 | CoV_Prot_S_829‐843 | CoV_Prot_S_802‐852 | DRB1_0101 | FIKQYGDCL | 1 | 0 | 0 | 1 | 1 | 1/1 (100) | 1 | 0 | 0 | 0 | 0 | 0/1 |
| DRB1_0101_P3 | CoV_Prot_S_896‐910 | CoV_Prot_S_881‐927 | DRB1_0101 | FAMQMAYRF | 1 | 0 | 0 | 1 | 1 | 1/1 (100) | 1 | 0 | 0 | 0 | 0 | 0/1 |
| DRB1_0101_P4 | CoV_Prot_S_813‐827 | CoV_Prot_S_802‐852 | DRB1_0101 | IEDLLFNKV | 1 | 0 | 0 | 1 | 1 | 1/1 (100) | 1 | 0 | 1 | 0 | 1 | 1/1 (100) |
| DRB1_0301_0401_P1 | CoV_Prot_S_445‐459 | CoV_Prot_S_441‐475 | DRB1_0301 | YNYLYRLFR | 1 | 0 | 1 | 0 | 1 | 1/1 (100) | 2 | 0 | 0 | 0 | 0 | 0/2 |
| DRB1_0301_0401_P1 | CoV_Prot_S_445‐459 | CoV_Prot_S_441‐475 | DRB1_0401 | YNYLYRLFR | 1 | 0 | 0 | 0 | 0 | 0/1 | 4 | 0 | 0 | 0 | 0 | 0/4 |
| DRB1_0301_P1 | CoV_Prot_S_457‐471 | CoV_Prot_S_441‐475 | DRB1_0301 | LKPFERDIS | 1 | 0 | 0 | 0 | 0 | 0/1 | 2 | 0 | 0 | 0 | 0 | 0/2 |
| DRB1_0301_P2 | CoV_Prot_S_765‐779 | CoV_Prot_S_737‐779 | DRB1_0301 | LTGIAVEQD | 2 | 0 | 0 | 0 | 0 | 0/2 | 3 | 0 | 1 | 0 | 1 | 1/3 (33,33) |
| DRB1_0401_0701_P1 | CoV_Prot_M_201‐215 | CoV_Prot_M_173‐222 | DRB1_0401 | YKLNTDHSS | 1 | 0 | 0 | 0 | 0 | 0/1 | 3 | 0 | 0 | 1 | 1 | 1/3 (33) |
| DRB1_0401_0701_P1 | CoV_Prot_M_201‐215 | CoV_Prot_M_173‐222 | DRB1_0701 | YKLNTDHSS | 1 | 0 | 0 | 0 | 0 | 0/1 | 3 | 0 | 0 | 1 | 1 | 1/3 (33) |
| DRB1_0401_1501_P1 | CoV_Prot_S_799‐813 | CoV_Prot_S_769‐813 | DRB1_0401 | ILPDPSKPS | 1 | 0 | 0 | 0 | 0 | 0/1 | 3 | 0 | 0 | 2 | 2 | 2/3 (66,67) |
| DRB1_0401_1501_P1 | CoV_Prot_S_799‐813 | CoV_Prot_S_769‐813 | DRB1_1501 | ILPDPSKPS | 5 | 0 | 0 | 1 | 1 | 1/5 (20) | 6 | 0 | 0 | 0 | 0 | 0/6 |
| DRB1_0401_HLA‐A02:01_P1 | CoV_Prot_S_817‐831 | CoV_Prot_S_802‐852 | DRB1_0401 | LLFNKVTLA | 1 | 0 | 0 | 0 | 0 | 0/1 | 3 | 0 | 0 | 1 | 1 | 1/3 (33) |
| DRB1_0401_HLA‐A02:01_P1 | CoV_Prot_S_817‐831 | CoV_Prot_S_802‐852 | HLA‐A02:01 | LLFNKVTLA | 9 | 0 | 0 | 0 | 0 | 0/9 | 8 | 0 | 1 | 1 | 2 | 2/8 (25) |
| DRB1_0401_P1 | CoV_Prot_S_813‐827 | CoV_Prot_S_802‐852 | DRB1_0401 | FIEDLLFNK | 1 | 0 | 0 | 0 | 0 | 0/1 | 3 | 0 | 0 | 1 | 1 | 1/3 (33) |
| DRB1_0701_P2 | CoV_Prot_M_173‐187 | CoV_Prot_M_173‐222 | DRB1_0701 | YKLGASQRV | 1 | 0 | 0 | 0 | 0 | 0/1 | 2 | 0 | 0 | 1 | 1 | 1/2 (50) |
| DRB1_1101_HLA‐A02:01_P1 | CoV_Prot_S_265‐279 | CoV_Prot_S_245‐295 | DRB1_1101 | YLQPRTFLL | 3 | 0 | 0 | 0 | 0 | 0/3 | 3 | 0 | 0 | 2 | 2 | 2/3 (67) |
| DRB1_1101_HLA‐A02:01_P1 | CoV_Prot_S_265‐279 | CoV_Prot_S_245‐295 | HLA‐A02:01 | YLQPRTFLL | 10 | 0 | 0 | 1 | 1 | 1/10 (10) | 10 | 0 | 0 | 8 | 8 | 8/10 (80) |
| DRB1_1101_P1 | CoV_Prot_M_009‐023 | CoV_Prot_M_1‐47 | DRB1_1101 | VEELKKLLE | 2 | 0 | 1 | 0 | 1 | 1/2 (50) | 2 | 0 | 0 | 0 | 0 | 0/2 |
| DRB1_1101_P2 | CoV_Prot_S_269‐283 | CoV_Prot_S_245‐295 | DRB1_1101 | FLLKYNENG | 3 | 0 | 0 | 0 | 0 | 0/3 | 3 | 0 | 0 | 0 | 0 | 0/3 |
| DRB1_1101_P3 | CoV_Prot_S_150‐164 | CoV_Prot_S_121‐175 | DRB1_1101 | MESEFRVYS | 2 | 0 | 0 | 0 | 0 | 0/2 | 2 | 0 | 0 | 1 | 1 | 1/2 (50) |
| DRB1_1101_P4 | CoV_Prot_S_349‐363 | CoV_Prot_S_342‐390 | DRB1_1101 | YAWNRKRIS | 2 | 0 | 0 | 0 | 0 | 0/2 | 1 | 1 | 0 | 0 | 1 | 1/1 (100) |
| DRB1_1101_P6 | CoV_Prot_S_253‐267 | CoV_Prot_S_245‐295 | DRB1_1101 | WTAGAAAYY | 3 | 0 | 0 | 0 | 0 | 0/3 | 3 | 0 | 0 | 0 | 0 | 0/3 |
| DRB1_1301_HLA‐C14:02_P1 | CoV_Prot_S_781‐795 | CoV_Prot_S_769‐813 | DRB1_1301 | VFAQVKQIY | 1 | 1 | 0 | 0 | 1 | 1/1 (100) | 1 | 0 | 0 | 0 | 0 | 0/1 |
| DRB1_1301_HLA‐C14:02_P1 | CoV_Prot_S_781‐795 | CoV_Prot_S_769‐813 | HLA‐C14:02 | VFAQVKQIY | 2 | 1 | 0 | 0 | 1 | 1/2 (50) | 2 | 0 | 0 | 0 | 0 | 0/2 |
| DRB1_1501_HLA‐C03:03_P1 | CoV_Prot_S_202‐216 | CoV_Prot_S_165‐216 | DRB1_1501 | YSKHTPINL | 1 | 0 | 0 | 1 | 1 | 1/1 (100) | 1 | 0 | 0 | 0 | 0 | 0/1 |
| DRB1_1501_HLA‐C03:03_P1 | CoV_Prot_S_198‐212 | CoV_Prot_S_165‐216 | HLA‐C03:03 | YSKHTPINL | 2 | 0 | 0 | 0 | 0 | 0/2 | 1 | 0 | 0 | 0 | 0 | 0/1 |
| DRB1_1501_P1 | CoV_Prot_M_173‐187 | CoV_Prot_M_173‐222 | DRB1_1501 | LSYYKLGAS | 5 | 2 | 0 | 0 | 2 | 2/5 (40) | 6 | 1 | 1 | 0 | 2 | 2/6 (33) |
| DRB1_1501_P10 | CoV_Prot_S_194‐208 | CoV_Prot_S_165‐216 | DRB1_1501 | IDGYFKIYS | 5 | 0 | 0 | 1 | 1 | 1/5 (20) | 6 | 0 | 0 | 0 | 0 | 0/6 |
| DRB1_1501_P11 | CoV_Prot_S_353‐367 | CoV_Prot_S_342‐390 | DRB1_1501 | ISNCVADYS | 5 | 0 | 1 | 1 | 2 | 2/5 (40) | 5 | 0 | 0 | 0 | 0 | 0/5 |
| DRB1_1501_P12 | CoV_Prot_S_198‐212 | CoV_Prot_S_165‐216 | DRB1_1501 | FKIYSKHTP | 5 | 0 | 0 | 1 | 1 | 1/5 (20) | 6 | 0 | 0 | 0 | 0 | 0/6 |
| DRB1_1501_P13 | CoV_Prot_S_345‐359 | CoV_Prot_S_342‐390 | DRB1_1501 | VYAWNRKRI | 5 | 0 | 1 | 1 | 2 | 2/5 (40) | 6 | 0 | 1 | 0 | 1 | 1/6 (17) |
| DRB1_1501_P14 | CoV_Prot_S_753‐765 | CoV_Prot_S_737‐779 | DRB1_1501 | YGSFCTQLN | 5 | 0 | 0 | 1 | 1 | 1/5 (20) | 6 | 1 | 0 | 0 | 1 | 1/6 (17) |
| DRB1_1501_P15 | CoV_Prot_S_990‐1004 | CoV_Prot_S_981‐1023 | DRB1_1501 | IDRLITGRL | 5 | 0 | 0 | 0 | 0 | 0/5 | 6 | 1 | 0 | 0 | 1 | 1/6 (17) |
| DRB1_1501_P16 | CoV_Prot_S_997‐1011 | CoV_Prot_S_981‐1023 | DRB1_1501 | LQSLQTYVT | 5 | 1 | 0 | 1 | 2 | 2/5 (40) | 6 | 0 | 0 | 1 | 1 | 1/6 (17) |
| DRB1_1501_P17 | CoV_Prot_S_1056‐1070 | CoV_Prot_S_1051‐1099 | DRB1_1501 | VVFLHVTYV | 5 | 0 | 0 | 0 | 0 | 0/5 | 6 | 0 | 0 | 1 | 1 | 1/6 (17) |
| DRB1_1501_P18 | CoV_Prot_S_342‐356 | CoV_Prot_S_342‐390 | DRB1_1501 | FASVYAWNR | 5 | 0 | 0 | 0 | 0 | 0/5 | 5 | 0 | 1 | 0 | 1 | 2/6 (33) |
| DRB1_1501_P19 | CoV_Prot_N_49‐61 | CoV_Prot_N_40‐87 | DRB1_1501 | WFTALTQHG | 5 | 0 | 0 | 0 | 0 | 0/5 | 6 | 0 | 1 | 0 | 1 | 1/6 (16,67) |
| DRB1_1501_P2 | CoV_Prot_M_197‐211 | CoV_Prot_M_173‐222 | DRB1_1501 | IGNYKLNTD | 5 | 1 | 3 | 0 | 4 | 4/5 (80) | 5 | 1 | 0 | 1 | 2 | 2/5 (40) |
| DRB1_1501_P3 | CoV_Prot_N_037‐051 | CoV_Prot_N_1‐51 | DRB1_1501 | PQGLPNNTA | 5 | 0 | 0 | 0 | 0 | 0/5 | 6 | 0 | 1 | 1 | 2 | 2/6 (33) |
| DRB1_1501_P4 | CoV_Prot_S_001‐014 | CoV_Prot_S_1‐51 | DRB1_1501 | LVLLPLVSS | 5 | 0 | 1 | 1 | 2 | 2/5 (40) | 6 | 0 | 0 | 0 | 0 | 0/6 |
| DRB1_1501_P5 | CoV_Prot_S_369‐383 | CoV_Prot_S_342‐390 | DRB1_1501 | FSTFKCYGV | 5 | 0 | 0 | 0 | 0 | 0/5 | 6 | 0 | 0 | 0 | 0 | 0/6 |
| DRB1_1501_P6 | CoV_Prot_S_376‐390 | CoV_Prot_S_342‐390 | DRB1_1501 | FKCYGVSPT | 5 | 0 | 0 | 0 | 0 | 0/5 | 5 | 0 | 0 | 0 | 0 | 0/5 |
| DRB1_1501_P7 | CoV_Prot_S_072‐086 | CoV_Prot_S_41‐91 | DRB1_1501 | TKRFDNPVL | 5 | 0 | 0 | 0 | 0 | 0/5 | 6 | 0 | 0 | 0 | 0 | 0/6 |
| DRB1_1501_P8 | CoV_Prot_S_138‐152 | CoV_Prot_S_121‐175 | DRB1_1501 | VYYHKNNKS | 5 | 0 | 0 | 0 | 0 | 0/5 | 6 | 0 | 0 | 0 | 0 | 0/6 |
| DRB1_1501_P9 | CoV_Prot_S_189‐203 | CoV_Prot_S_165‐216 | DRB1_1501 | FVFKNIDGY | 5 | 0 | 0 | 0 | 0 | 0/5 | 6 | 0 | 0 | 0 | 0 | 0/6 |
| HLA‐A01:01_P1 | CoV_Prot_M_208‐222 | CoV_Prot_M_173‐222 | HLA‐A01:01 | SSDNIALLV | 7 | 0 | 0 | 0 | 0 | 0/7 | 7 | 0 | 1 | 1 | 2 | 2/7 (29) |
| HLA‐A01:01_P2 | CoV_Prot_N_405‐419 | CoV_Prot_N_377‐419 | HLA‐A01:01 | MSSADSTQA | 12 | 7 | 2 | 0 | 9 | 9/12 (75) | 7 | 0 | 1 | 2 | 3 | 3/7 (43) |
| HLA‐A01:01_P3 | CoV_Prot_S_157‐171 | CoV_Prot_S_121‐175 | HLA‐A01:01 | SANNCTFEY | 4 | 1 | 0 | 1 | 2 | 2/4 (50) | 5 | 0 | 0 | 1 | 1 | 1/5 (20) |
| HLA‐A01:01_P4 | CoV_Prot_M_165‐179 | CoV_Prot_M_135‐183 | HLA‐A01:01 | ATSRTLSYY | 6 | 0 | 1 | 0 | 1 | 1/6 (16,67) | 6 | 0 | 2 | 0 | 2 | 2/6 (33,33) |
| HLA‐A02:01_HLAC04:01_P1 | CoV_Prot_S_105‐119 | CoV_Prot_S_81‐130 | HLA‐A02:01 | TLDSKTQSL | 10 | 0 | 0 | 0 | 0 | 0/10 | 10 | 1 | 0 | 3 | 4 | 4/10 (40) |
| HLA‐A02:01_HLAC04:01_P1 | CoV_Prot_S_105‐119 | CoV_Prot_S_81‐130 | HLA‐C04:01 | TLDSKTQSL | 1 | 0 | 0 | 1 | 1 | 1/1 (100) | 1 | 0 | 0 | 0 | 0 | 0/1 |
| HLA‐A02:01_P1 | CoV_Prot_M_012‐026 | CoV_Prot_M_1‐47 | HLA‐A02:01 | KLLEQWNLV | 6 | 0 | 0 | 0 | 0 | 0/6 | 6 | 0 | 0 | 1 | 1 | 1/6 (17) |
| HLA‐A02:01_P10 | CoV_Prot_S_972‐984 | CoV_Prot_S_955‐991 | HLA‐A02:01 | VLNDILSRL | 10 | 0 | 0 | 0 | 0 | 0/10 | 10 | 0 | 0 | 3 | 3 | 3/10 (30) |
| HLA‐A02:01_P11 | CoV_Prot_S_981‐995 | CoV_Prot_S_981‐1023 | HLA‐A02:01 | RLDKVEAEV | 5 | 0 | 0 | 0 | 0 | 0/5 | 4 | 1 | 0 | 1 | 2 | 2/4 (50) |
| HLA‐A02:01_P12 | CoV_Prot_S_1045‐1059 | CoV_Prot_S_1012‐1059 | HLA‐A02:01 | HLMSFPQSA | 10 | 0 | 0 | 1 | 1 | 1/10 (10) | 10 | 0 | 0 | 1 | 1 | 1/10 (10) |
| HLA‐A02:01_P13 | CoV_Prot_S_1209‐1223 | CoV_Prot_S_1195‐1230 | HLA‐A02:01 | IKWPWYIWL | 9 | 0 | 0 | 0 | 0 | 0/9 | 9 | 0 | 0 | 1 | 1 | 1/9 (11,11) |
| HLA‐A02:01_P14 | CoV_Prot_S_245‐259 | CoV_Prot_S_245‐295 | HLA‐A02:01 | YLTPGDSSS | 9 | 0 | 0 | 0 | 0 | 0/9 | 9 | 0 | 1 | 1 | 2 | 2/9 (22,22) |
| HLA‐A02:01_P2 | CoV_Prot_M_060‐070 | CoV_Prot_M_37‐73 | HLA‐A02:01 | TLACFVLAA | 5 | 0 | 1 | 0 | 1 | 1/5 (20) | 6 | 0 | 0 | 2 | 2 | 2/6 (33) |
| HLA‐A02:01_P3 | CoV_Prot_S_685‐699 | CoV_Prot_S_665‐707 | HLA‐A02:01 | SIIAYTMSL | 5 | 0 | 1 | 0 | 1 | 1/5 (20) | 5 | 0 | 0 | 2 | 2 | 2/5 (40) |
| HLA‐A02:01_P6 | CoV_Prot_M_65‐76 | CoV_Prot_M_65‐105 | HLA‐A02:01 | FVLAAVYRI | 6 | 0 | 0 | 0 | 0 | 0/6 | 6 | 0 | 0 | 0 | 0 | 0/6 |
| HLA‐A02:01_P7 | CoV_Prot_N_217‐230 | CoV_Prot_N_194‐234 | HLA‐A02:01 | LLLDRLNQL | 6 | 0 | 0 | 0 | 0 | 0/6 | 7 | 0 | 1 | 0 | 1 | 1/7 (14,29) |
| HLA‐A02:01_P8 | CoV_Prot_S_417‐431 | CoV_Prot_S_381‐431 | HLA‐A02:01 | KIADYNYKL | 10 | 0 | 0 | 0 | 0 | 0/10 | 9 | 0 | 0 | 0 | 0 | 0/9 |
| HLA‐A02:01_P9 | CoV_Prot_S_915‐927 | CoV_Prot_S_881‐927 | HLA‐A02:01 | VLYENQKLI | 5 | 1 | 1 | 0 | 2 | 2/5 (40) | 5 | 0 | 0 | 2 | 2 | 2/5 (40) |
| HLA‐A03:01_HLA‐A11:01_P1 | CoV_Prot_S_1061‐1075 | CoV_Prot_S_1051‐1099 | HLA‐A03:01 | VTYVPAQEK | 6 | 0 | 0 | 0 | 0 | 0/6 | 8 | 0 | 0 | 5 | 5 | 5/8 (62,5) |
| HLA‐A03:01_HLA‐A11:01_P1 | CoV_Prot_S_1061‐1075 | CoV_Prot_S_1051‐1099 | HLA‐A11:01 | VTYVPAQEK | 1 | 0 | 0 | 0 | 0 | 0/1 | 1 | 0 | 0 | 1 | 1 | 1/1 (100) |
| HLA‐A03:01_P1 | CoV_Prot_M_149‐163 | CoV_Prot_M_135‐183 | HLA‐A03:01 | RIAGHHLGR | 6 | 1 | 0 | 0 | 1 | 1/6 (17) | 7 | 1 | 0 | 1 | 2 | 2/7 (29) |
| HLA‐A03:01_P2 | CoV_Prot_S_345‐359 | CoV_Prot_S_342‐390 | HLA‐A03:01 | SVYAWNRKR | 8 | 0 | 0 | 1 | 1 | 1/8 (13) | 11 | 2 | 0 | 2 | 4 | 4/11 (36) |
| HLA‐A03:01_P3 | CoV_Prot_S_376‐390 | CoV_Prot_S_342‐390 | HLA‐A03:01 | KCYGVSPTK | 6 | 0 | 0 | 4 | 4 | 4/6 (66,67) | 8 | 1 | 0 | 7 | 8 | 8/8 (100) |
| HLA‐A03:01_P4 | CoV_Prot_S_526‐539 | CoV_Prot_S_521‐567 | HLA‐A03:01 | KSTNLVKNK | 7 | 0 | 0 | 1 | 1 | 1/7 (14,29) | 10 | 0 | 0 | 2 | 2 | 2/10 (20) |
| HLA‐A03:01_P5 | CoV_Prot_S_41‐55 | CoV_Prot_S_41‐91 | HLA‐A03:01 | KVFRSSVLH | 6 | 0 | 0 | 0 | 0 | 0/6 | 7 | 0 | 0 | 2 | 2 | 2/7 (28,57) |
| HLA‐A03:01_P6 | CoV_Prot_S_138‐152 | CoV_Prot_S_121‐175 | HLA‐A03:01 | GVYYHKNNK | 8 | 1 | 0 | 0 | 1 | 1/8 (12,5) | 10 | 1 | 0 | 0 | 1 | 1/10 (10) |
| HLA‐A11:01_P1 | CoV_Prot_S_669‐683 | CoV_Prot_S_665‐707 | HLA‐A11:01 | QTQTNSPRR | 1 | 0 | 1 | 0 | 1 | 1/1 (100) | 1 | 0 | 0 | 0 | 0 | 0/1 |
| HLA‐A11:01_P2 | CoV_Prot_S_719‐733 | CoV_Prot_S_698‐747 | HLA‐A11:01 | EILPVSMTK | 1 | 0 | 0 | 0 | 0 | 0/1 | 1 | 0 | 0 | 1 | 1 | 1/1 (100) |
| HLA‐A11:01_P3 | CoV_Prot_S_753‐765 | CoV_Prot_S_737‐779 | HLA‐A11:01 | GSFCTQLNR | 1 | 0 | 0 | 0 | 0 | 0/1 | 1 | 0 | 0 | 1 | 1 | 1/1 (100) |
| HLA‐A11:01_P4 | CoV_Prot_S_546‐560 | CoV_Prot_S_521‐567 | HLA‐A11:01 | GVLTESNKK | 2 | 0 | 0 | 0 | 0 | 0/2 | 1 | 0 | 0 | 0 | 0 | 0/1 |
| HLA‐A11:01_P5 | CoV_Prot_S_781‐795 | CoV_Prot_S_769‐813 | HLA‐A11:01 | QIYKTPPIK | 1 | 0 | 0 | 0 | 0 | 0/1 | 1 | 0 | 0 | 1 | 1 | 1/1 (100) |
| HLA‐A11:01_P6 | CoV_Prot_S_1093‐1107 | CoV_Prot_S_1089‐1135 | HLA‐A11:01 | GTHWFVTQR | 1 | 0 | 0 | 0 | 0 | 0/1 | 1 | 0 | 0 | 0 | 0 | 0/1 |
| HLA‐A11:01_P7 | CoV_Prot_S_364‐378 | CoV_Prot_S_342‐390 | HLA‐A11:01 | NSASFSTFK | 1 | 0 | 0 | 0 | 0 | 0/1 | 1 | 0 | 0 | 0 | 0 | 0/1 |
| HLA‐A23:01_P1 | CoV_Prot_M_042‐056 | CoV_Prot_M_37‐73 | HLA‐A23:01 | LYIIKLIFL | 1 | 0 | 0 | 0 | 0 | 0/1 | 1 | 0 | 0 | 0 | 0 | 0/1 |
| HLA‐A23:01_P2 | CoV_Prot_M_89‐103 | CoV_Prot_M_65‐105 | HLA‐A23:01 | YFIASFRLF | 1 | 0 | 0 | 0 | 0 | 0/1 | 1 | 0 | 0 | 0 | 0 | 0/1 |
| HLA‐A25:01_P1 | CoV_Prot_M_12‐26 | CoV_Prot_M_1‐47 | HLA‐A25:01 | ELKKLLEQW | 1 | 0 | 0 | 0 | 0 | 0/1 | 2 | 0 | 0 | 0 | 0 | 0/2 |
| HLA‐A26:01_P1 | CoV_Prot_S_044‐058 | CoV_Prot_S_41‐91 | HLA‐A26:01 | STQDLFLPF | 1 | 0 | 0 | 0 | 0 | 0/1 | 1 | 0 | 0 | 0 | 0 | 0/1 |
| HLA‐A29:02_HLA‐C02:02_P1 | CoV_Prot_S_024‐038 | CoV_Prot_S_1‐51 | HLA‐A29:02 | NSFTRGVYY | 1 | 0 | 0 | 0 | 0 | 0/1 | 1 | 0 | 0 | 0 | 0 | 0/1 |
| HLA‐A29:02_HLA‐C02:02_P1 | CoV_Prot_S_024‐038 | CoV_Prot_S_1‐51 | HLA‐C02:02 | NSFTRGVYY | 1 | 0 | 0 | 0 | 0 | 0/1 | 2 | 0 | 0 | 0 | 0 | 0/2 |
| HLA‐A68:01_P1 | CoV_Prot_M_137‐151 | CoV_Prot_M_135‐183 | HLA‐A68:01 | LVIGAVILR | 2 | 0 | 0 | 0 | 0 | 0/2 | 1 | 0 | 0 | 0 | 0 | 0/1 |
| HLA‐A68:01_P2 | CoV_Prot_S_673‐687 | CoV_Prot_S_665‐707 | HLA‐A68:01 | QTNSPRRAR | 2 | 0 | 0 | 0 | 0 | 0/2 | 2 | 0 | 0 | 0 | 0 | 0/2 |
| HLA‐B07:02_P1 | CoV_Prot_N_102‐116 | CoV_Prot_N_78‐127 | HLA‐B07:02 | SPRWYFYYL | 5 | 0 | 0 | 0 | 0 | 0/5 | 6 | 1 | 1 | 2 | 4 | 4/6 (67) |
| HLA‐B07:02_P2 | CoV_Prot_N_37‐51 | CoV_Prot_N_1‐51 | HLA‐B07:02 | RPQGLPNNT | 6 | 0 | 0 | 0 | 0 | 0/6 | 6 | 0 | 0 | 0 | 0 | 0/6 |
| HLA‐B07:02_P3 | CoV_Prot_N_253‐267 | CoV_Prot_N_224‐274 | HLA‐B07:02 | KPRQKRTAT | 6 | 0 | 0 | 0 | 0 | 0/6 | 7 | 0 | 1 | 1 | 2 | 2/7 (28,57) |
| HLA‐B07:02_P4 | CoV_Prot_S_678‐692 | CoV_Prot_S_665‐707 | HLA‐B07:02 | SPRRARSVA | 8 | 0 | 0 | 1 | 1 | 1/8 (12,5) | 7 | 0 | 0 | 4 | 4 | 4/7 (57,14) |
| HLA‐B07:02_P5 | CoV_Prot_S_202‐216 | CoV_Prot_S_165‐216 | HLA‐B07:02 | TPINLVRDL | 7 | 0 | 0 | 0 | 0 | 0/7 | 8 | 0 | 0 | 0 | 0 | 0/8 |
| HLA‐B08:01_P1 | CoV_Prot_M_144‐158 | CoV_Prot_M_135‐183 | HLA‐B08:01 | HLRIAGHHL | 4 | 0 | 0 | 0 | 0 | 0/4 | 5 | 0 | 1 | 0 | 1 | 1/5 (20) |
| HLA‐B08:01_P2 | CoV_Prot_S_869‐883 | CoV_Prot_S_841‐891 | HLA‐B08:01 | MIAQYTSAL | 4 | 0 | 0 | 0 | 0 | 0/4 | 6 | 0 | 1 | 1 | 2 | 2/6 (33,33) |
| HLA‐B14:01_P1 | CoV_Prot_S_37‐51 | CoV_Prot_S_1‐51 | HLA‐B14:01 | DKVFRSSVL | 1 | 0 | 0 | 0 | 0 | 0/1 | 1 | 0 | 0 | 0 | 0 | 0/1 |
| HLA‐B14:01_P2 | CoV_Prot_S_41‐55 | CoV_Prot_S_41‐91 | HLA‐B14:01 | FRSSVLHST | 1 | 0 | 0 | 0 | 0 | 0/1 | 1 | 0 | 0 | 0 | 0 | 0/1 |
| HLA‐B15:01_P1 | CoV_Prot_S_494‐508 | CoV_Prot_S_461‐508 | HLA‐B15:01 | FQPTNGVGY | 2 | 0 | 0 | 0 | 0 | 0/2 | 2 | 0 | 0 | 2 | 2 | 2/2 (100) |
| HLA‐B15:01_P2 | CoV_Prot_S_799‐813 | CoV_Prot_S_769‐813 | HLA‐B15:01 | SQILPDPSK | 1 | 0 | 1 | 0 | 1 | 1/1 (100) | 1 | 0 | 0 | 1 | 1 | 1/1 (100) |
| HLA‐B15:01_P3 | CoV_Prot_S_178‐192 | CoV_Prot_S_165‐216 | HLA‐B15:01 | GNFKNLREF | 2 | 0 | 0 | 0 | 0 | 0/2 | 2 | 0 | 0 | 0 | 0 | 0/2 |
| HLA‐B27:05_HLA‐C07:02_P1 | CoV_Prot_S_994‐1008 | CoV_Prot_S_981‐1023 | HLA‐B27:05 | GRLQSLQTY | 1 | 0 | 0 | 0 | 0 | 0/1 | 1 | 0 | 0 | 1 | 1 | 1/1 (100) |
| HLA‐B27:05_HLA‐C07:02_P1 | CoV_Prot_S_994‐1008 | CoV_Prot_S_981‐1023 | HLA‐C07:02 | GRLQSLQTY | 7 | 0 | 0 | 1 | 1 | 1/7 (14) | 9 | 0 | 0 | 2 | 2 | 2/9 (22) |
| HLA‐B27:05_P1 | CoV_Prot_S_077‐091 | CoV_Prot_S_41‐91 | HLA‐B27:05 | KRFDNPVLP | 1 | 1 | 0 | 0 | 1 | 1/1 (100) | 1 | 0 | 0 | 0 | 0 | 0/1 |
| HLA‐B27:05_P2 | CoV_Prot_S_342‐356 | CoV_Prot_S_342‐390 | HLA‐B27:05 | TRFASVYAW | 1 | 0 | 0 | 0 | 0 | 0/1 | 1 | 0 | 0 | 0 | 0 | 0/1 |
| HLA‐B35:03_P1 | CoV_Prot_S_081‐095 | CoV_Prot_S_81‐130 | HLA‐B35:03 | LPFNDGVYF | 1 | 0 | 0 | 1 | 1 | 1/1 (100) | 1 | 0 | 0 | 0 | 0 | 0/1 |
| HLA‐B35:03_P2 | CoV_Prot_S_1257‐1271 | CoV_Prot_S_1222‐1273 | HLA‐B35:03 | EPVLKGVKL | 1 | 0 | 0 | 0 | 0 | 0/1 | 1 | 0 | 0 | 1 | 1 | 1/1 (100) |
| HLA‐B35:03_P3 | CoV_Prot_S_18‐32 | CoV_Prot_S_1‐51 | HLA‐B35:03 | LPPAYTNSF | 1 | 0 | 0 | 0 | 0 | 0/1 | 1 | 0 | 0 | 0 | 0 | 0/1 |
| HLA‐B38:01_P1 | CoV_Prot_S_769‐783 | CoV_Prot_S_769‐813 | HLA‐B38:01 | EQDKNTQEV | 1 | 0 | 0 | 0 | 0 | 0/1 | 1 | 0 | 1 | 0 | 1 | 1/1 (100) |
| HLA‐B40:01_P1 | CoV_Prot_S_1149‐1163 | CoV_Prot_S_1123‐1166 | HLA‐B40:01 | KYFKNHTSP | 1 | 0 | 0 | 0 | 0 | 0/1 | 2 | 0 | 0 | 0 | 0 | 0/2 |
| HLA‐B40:01_P2 | CoV_Prot_S_1175‐1189 | CoV_Prot_S_1157‐1205 | HLA‐B40:01 | KEIDRLNEV | 4 | 0 | 0 | 0 | 0 | 0/4 | 3 | 0 | 0 | 0 | 0 | 0/3 |
| HLA‐B44:02_P1 | CoV_Prot_S_1203‐1215 | CoV_Prot_S_1195‐1230 | HLA‐B44:02 | YEQYIKWPW | 2 | 0 | 0 | 0 | 0 | 0/2 | 2 | 0 | 0 | 0 | 0 | 0/2 |
| HLA‐B44:02_P2 | CoV_Prot_S_825‐839 | CoV_Prot_S_802‐852 | HLA‐B44:02 | ADAGFIKQY | 2 | 0 | 0 | 0 | 0 | 0/2 | 2 | 0 | 0 | 1 | 1 | 1/2 (50) |
| HLA‐B44:02_P3 | CoV_Prot_S_1197‐1211 | CoV_Prot_S_1195‐1230 | HLA‐B44:02 | QELGKYEQY | 2 | 0 | 0 | 0 | 0 | 0/2 | 2 | 0 | 0 | 0 | 0 | 0/2 |
| HLA‐B44:03_P1 | CoV_Prot_N_321‐335 | CoV_Prot_N_305‐347 | HLA‐B44:03 | MEVTPSGTW | 1 | 0 | 0 | 0 | 0 | 0/1 | 1 | 0 | 0 | 0 | 0 | 0/1 |
| HLA‐B51:01_P1 | CoV_Prot_S_005‐019 | CoV_Prot_S_1‐51 | HLA‐B51:01 | LPLVSSQCV | 4 | 0 | 1 | 0 | 1 | 1/4 (25) | 5 | 0 | 0 | 0 | 0 | 0/5 |
| HLA‐B51:01_P2 | CoV_Prot_S_709‐723 | CoV_Prot_S_698‐747 | HLA‐B51:01 | IPTNFTISV | 5 | 0 | 0 | 0 | 0 | 0/5 | 5 | 0 | 0 | 1 | 1 | 1/5 (20) |
| HLA‐C02:02_DRB1_0701_P1 | CoV_Prot_M_165‐179 | CoV_Prot_M_135‐183 | HLA‐C02:02 | VATSRTLSY | 1 | 0 | 0 | 0 | 0 | 0/1 | 2 | 1 | 0 | 0 | 1 | 1/2 (50) |
| HLA‐C02:02_DRB1_0701_P1 | CoV_Prot_M_177‐191 | CoV_Prot_M_173‐222 | DRB1_0701 | VATSRTLSY | 1 | 0 | 0 | 0 | 0 | 0/1 | 3 | 0 | 0 | 0 | 0 | 0/3 |
| HLA‐C02:02_HLA‐C12:02_P1 | CoV_Prot_S_683‐696 | CoV_Prot_S_665‐707 | HLA‐C02:02 | VASQSIIAY | 1 | 0 | 0 | 0 | 0 | 0/1 | 2 | 0 | 0 | 0 | 0 | 0/2 |
| HLA‐C02:02_HLA‐C12:02_P1 | CoV_Prot_S_683‐696 | CoV_Prot_S_665‐707 | HLA‐C12:02 | VASQSIIAY | 1 | 0 | 0 | 0 | 0 | 0/1 | 2 | 0 | 0 | 0 | 0 | 0/2 |
| HLA‐C02:02_P1 | CoV_Prot_M_193‐207 | CoV_Prot_M_173‐222 | HLA‐C02:02 | YSRYRIGNY | 1 | 0 | 0 | 0 | 0 | 0/1 | 2 | 0 | 0 | 0 | 0 | 0/2 |
| HLA‐C02:02_P3 | CoV_Prot_S_170‐184 | CoV_Prot_S_165‐216 | HLA‐C02:02 | VSQPFLMDL | 1 | 0 | 0 | 0 | 0 | 0/1 | 2 | 0 | 0 | 0 | 0 | 0/2 |
| HLA‐C02:02_P4 | CoV_Prot_S_304‐319 | CoV_Prot_S_285‐327 | HLA‐C02:02 | KGIYQTSNF | 1 | 0 | 0 | 0 | 0 | 0/1 | 2 | 0 | 0 | 0 | 0 | 0/2 |
| HLA‐C02:02_P5 | CoV_Prot_S_369‐383 | CoV_Prot_S_342‐390 | HLA‐C02:02 | ASFSTFKCY | 1 | 0 | 0 | 0 | 0 | 0/1 | 2 | 0 | 0 | 1 | 1 | 1/2 (50) |
| HLA‐C02:02_P7 | CoV_Prot_M_34‐47 | CoV_Prot_M_1‐47 | HLA‐C02:02 | FAYANRNRF | 1 | 0 | 0 | 0 | 0 | 0/1 | 2 | 1 | 0 | 1 | 2 | 2/2 (100) |
| HLA‐C02:02_P8 | CoV_Prot_M_27‐37 | CoV_Prot_M_1‐47 | HLA‐C02:02 | LTWICLLQF | 1 | 0 | 0 | 0 | 0 | 0/1 | 2 | 0 | 1 | 1 | 2 | 2/2 (100) |
| HLA‐C02:02_P9 | CoV_Prot_S_1259‐1273 | CoV_Prot_S_1222‐1273 | HLA‐C02:02 | VLKGVKLHY | 1 | 0 | 0 | 0 | 0 | 0/1 | 2 | 0 | 0 | 0 | 0 | 0/2 |
| HLA‐C03:04_P1 | CoV_Prot_S_881‐895 | CoV_Prot_S_881‐927 | HLA‐C03:04 | WTFGAGAAL | 2 | 0 | 1 | 0 | 1 | 1/2 (50) | 3 | 0 | 0 | 1 | 1 | 1/3 (33) |
| HLA‐C03:04_P2 | CoV_Prot_S_865‐879 | CoV_Prot_S_841‐891 | HLA‐C03:04 | IAQYTSALL | 5 | 0 | 0 | 0 | 0 | 0/5 | 5 | 0 | 1 | 0 | 1 | 1/5 (20) |
| HLA‐C04:01_DRB1_1501_P1 | CoV_Prot_S_165‐179 | CoV_Prot_S_165‐216 | HLA‐C04:01 | TFEYVSQPF | 4 | 0 | 0 | 0 | 0 | 0/4 | 4 | 0 | 0 | 0 | 0 | 0/4 |
| HLA‐C04:01_DRB1_1501_P1 | CoV_Prot_S_161‐175 | CoV_Prot_S_121‐175 | DRB1_1501 | TFEYVSQPF | 5 | 0 | 0 | 1 | 1 | 1/5 (20) | 6 | 0 | 0 | 0 | 0 | 0/6 |
| HLA‐C04:01_DRB1_1501_P1 | CoV_Prot_S_161‐175 | CoV_Prot_S_121‐175 | HLA‐C04:01 | TFEYVSQPF | 1 | 0 | 0 | 1 | 1 | 1/1 (100) | 1 | 0 | 0 | 1 | 1 | 1/1 (100) |
| HLA‐C04:01_HLA‐C07:02_P1 | CoV_Prot_S_72‐86 | CoV_Prot_S_41‐91 | HLA‐C04:01 | RFDNPVLPF | 4 | 0 | 0 | 1 | 1 | 1/4 (25) | 3 | 0 | 1 | 0 | 1 | 1/3 (33,33) |
| HLA‐C04:01_HLA‐C07:02_P1 | CoV_Prot_S_72‐86 | CoV_Prot_S_41‐91 | HLA‐C07:02 | RFDNPVLPF | 6 | 2 | 0 | 0 | 2 | 2/6 (33,33) | 6 | 0 | 0 | 1 | 1 | 1/6 (16,67) |
| HLA‐C04:01_P1 | CoV_Prot_M_105‐119 | CoV_Prot_M_97‐147 | HLA‐C04:01 | SFNPETNIL | 2 | 0 | 0 | 0 | 0 | 0/2 | 2 | 0 | 0 | 1 | 1 | 1/2 (50) |
| HLA‐C04:01_P2 | CoV_Prot_N_78‐92 | CoV_Prot_N_78‐127 | HLA‐C04:01 | SPDDQIGYY | 2 | 0 | 0 | 1 | 1 | 1/2 (50) | 2 | 0 | 0 | 0 | 0 | 0/2 |
| HLA‐C04:01_P3 | CoV_Prot_N_333‐347 | CoV_Prot_N_305‐347 | HLA‐C04:01 | KLDDKDPNF | 2 | 0 | 0 | 0 | 0 | 0/2 | 2 | 1 | 0 | 0 | 1 | 1/2 (50) |
| HLA‐C04:01_P6 | CoV_Prot_S_1136‐1149 | CoV_Prot_S_1123‐1166 | HLA‐C04:01 | VYDPLQPEL | 4 | 0 | 0 | 0 | 0 | 0/4 | 4 | 0 | 0 | 1 | 1 | 1/4 (25) |
| HLA‐C05:01_P1 | CoV_Prot_S_285‐299 | CoV_Prot_S_285‐327 | HLA‐C05:01 | ITDAVDCAL | 2 | 0 | 0 | 0 | 0 | 0/2 | 2 | 0 | 0 | 0 | 0 | 0/2 |
| HLA‐C05:01_P2 | CoV_Prot_S_581‐595 | CoV_Prot_S_557‐607 | HLA‐C05:01 | ILDITPCSF | 2 | 0 | 0 | 0 | 0 | 0/2 | 2 | 0 | 0 | 0 | 0 | 0/2 |
| HLA‐C05:01_P3 | CoV_Prot_S_713‐727 | CoV_Prot_S_698‐747 | HLA‐C05:01 | FTISVTTEI | 2 | 0 | 0 | 0 | 0 | 0/2 | 2 | 0 | 0 | 0 | 0 | 0/2 |
| HLA‐C05:01_P4 | CoV_Prot_S_745‐759 | CoV_Prot_S_737‐779 | HLA‐C05:01 | STECSNLLL | 2 | 0 | 0 | 0 | 0 | 0/2 | 2 | 0 | 0 | 0 | 0 | 0/2 |
| HLA‐C05:01_P5 | CoV_Prot_S_841‐855 | CoV_Prot_S_841‐891 | HLA‐C05:01 | LGDIAARDL | 2 | 0 | 0 | 0 | 0 | 0/2 | 2 | 0 | 0 | 1 | 1 | 1/2 (50) |
| HLA‐C05:01_P6 | CoV_Prot_S_1117‐1131 | CoV_Prot_S_1089‐1135 | HLA‐C05:01 | FVSGNCDVV | 2 | 0 | 0 | 0 | 0 | 0/2 | 2 | 0 | 0 | 0 | 0 | 0/2 |
| HLA‐C05:01_P7 | CoV_Prot_S_289‐303 | CoV_Prot_S_285‐327 | HLA‐C05:01 | ALDPLSETK | 2 | 0 | 0 | 0 | 0 | 0/2 | 2 | 0 | 1 | 1 | 2 | 2/2 (100) |
| HLA‐C07:01_P1 | CoV_Prot_S_553‐567 | CoV_Prot_S_521‐567 | HLA‐C07:01 | KKFLPFQQF | 4 | 0 | 0 | 1 | 1 | 1/4 (25) | 4 | 0 | 0 | 1 | 1 | 1/4 (25) |
| HLA‐C07:01_P2 | CoV_Prot_M_102‐116 | CoV_Prot_M_97‐147 | HLA‐C07:01 | ARTRSMWSF | 4 | 0 | 0 | 0 | 0 | 0/4 | 6 | 0 | 0 | 0 | 0 | 0/6 |
| HLA‐C07:01_P3 | CoV_Prot_S_785‐799 | CoV_Prot_S_769‐813 | HLA‐C07:01 | YKTPPIKDF | 5 | 0 | 0 | 1 | 1 | 1/5 (20) | 6 | 0 | 0 | 0 | 0 | 0/6 |
| HLA‐C07:02_P11 | CoV_Prot_S_322‐335 | CoV_Prot_S_313‐351 | HLA‐C07:02 | VRFPNITNL | 8 | 1 | 1 | 0 | 2 | 2/8 (25) | 9 | 0 | 1 | 0 | 1 | 1/9 (11,11) |
| HLA‐C07:02_P2 | CoV_Prot_M_169‐183 | CoV_Prot_M_135‐183 | HLA‐C07:02 | SRTLSYYKL | 7 | 1 | 0 | 0 | 1 | 1/7 (14,29) | 7 | 0 | 1 | 1 | 2 | 2/7 (28,57) |
| HLA‐C07:02_P3 | CoV_Prot_N_158‐172 | CoV_Prot_N_158‐203 | HLA‐C07:02 | LQLPQGTTL | 7 | 0 | 1 | 1 | 2 | 2/7 (28,57) | 7 | 0 | 1 | 0 | 1 | 1/7 (14,29) |
| HLA‐C07:02_P4 | CoV_Prot_S_438‐452 | CoV_Prot_S_421‐455 | HLA‐C07:02 | SKVGGNYNY | 8 | 0 | 0 | 2 | 2 | 2/8 (25) | 9 | 0 | 0 | 1 | 1 | 1/9 (11,11) |
| HLA‐C07:02_P5 | CoV_Prot_S_486‐500 | CoV_Prot_S_461‐508 | HLA‐C07:02 | YFPLQSYGF | 8 | 0 | 0 | 1 | 1 | 1/8 (12,5) | 7 | 0 | 1 | 2 | 3 | 3/7 (42,86) |
| HLA‐C07:02_P6 | CoV_Prot_S_573‐587 | CoV_Prot_S_557‐607 | HLA‐C07:02 | VRDPQTLEI | 8 | 0 | 0 | 2 | 2 | 2/8 (25) | 9 | 0 | 1 | 0 | 1 | 1/9 (11,11) |
| HLA‐C07:02_P7 | CoV_Prot_S_165‐179 | CoV_Prot_S_165‐216 | HLA‐C07:02 | EYVSQPFLM | 8 | 1 | 0 | 0 | 1 | 1/8 (12,5) | 9 | 0 | 0 | 2 | 2 | 2/9 (22,22) |
| HLA‐C07:02_P8 | CoV_Prot_S_18‐32 | CoV_Prot_S_1‐51 | HLA‐C07:02 | TRTQLPPAY | 7 | 0 | 0 | 1 | 1 | 1/7 (14,29) | 9 | 0 | 1 | 0 | 1 | 1/9 (11,11) |
| HLA‐C08:02_P1 | CoV_Prot_S_865‐879 | CoV_Prot_S_841‐891 | HLA‐C08:02 | LTDEMIAQY | 1 | 0 | 0 | 0 | 0 | 0/1 | 1 | 0 | 0 | 1 | 1 | 1/1 (100) |
| HLA‐C12:02_P1 | CoV_Prot_S_1189‐1203 | CoV_Prot_S_1157‐1205 | HLA‐C12:02 | VAKNLNESL | 1 | 0 | 0 | 0 | 0 | 0/1 | 2 | 0 | 0 | 0 | 0 | 0/2 |
| HLA‐C12:03_HLA‐B51:01_P1 | CoV_Prot_S_117‐130 | CoV_Prot_S_81‐130 | HLA‐B51:01 | NATNVVIKV | 5 | 0 | 0 | 0 | 0 | 0/5 | 5 | 0 | 0 | 3 | 3 | 3/5 (60) |
| HLA‐C12:03_HLA‐B51:01_P1 | CoV_Prot_S_121‐135 | CoV_Prot_S_121‐175 | HLA‐C12:03 | NATNVVIKV | 1 | 0 | 0 | 0 | 0 | 0/1 | 2 | 0 | 0 | 0 | 0 | 0/2 |
| HLA‐C12:03_HLA‐B51:01_P1 | CoV_Prot_S_121‐135 | CoV_Prot_S_121‐175 | HLA‐B51:01 | NATNVVIKV | 4 | 0 | 1 | 0 | 1 | 1/4 (25) | 5 | 0 | 0 | 3 | 3 | 3/5 (60) |
| HLA‐C12:03_P2 | CoV_Prot_S_249‐263 | CoV_Prot_S_245‐295 | HLA‐C12:03 | SSSGWTAGA | 2 | 0 | 0 | 0 | 0 | 0/2 | 4 | 1 | 0 | 0 | 1 | 1/4 (25) |
| HLA‐C14:02_P1 | CoV_Prot_M_193‐207 | CoV_Prot_M_173‐222 | HLA‐C14:02 | RYRIGNYKL | 2 | 0 | 0 | 0 | 0 | 0/2 | 2 | 0 | 0 | 0 | 0 | 0/2 |
| HLA‐C14:02_P2 | CoV_Prot_S_698‐712 | CoV_Prot_S_698‐747 | HLA‐C14:02 | LGAENSVAY | 2 | 0 | 0 | 0 | 0 | 0/2 | 2 | 0 | 0 | 1 | 1 | 1/2 (50) |
| HLA‐C15:02_P1 | CoV_Prot_S_024‐038 | CoV_Prot_S_1‐51 | HLA‐C15:02 | YTNSFTRGV | 1 | 0 | 0 | 0 | 0 | 0/1 | 1 | 0 | 0 | 0 | 0 | 0/1 |
| HLA‐C15:02_P2 | CoV_Prot_S_057‐071 | CoV_Prot_S_41‐91 | HLA‐C15:02 | VTWFHAIHV | 2 | 0 | 0 | 0 | 0 | 0/2 | 2 | 0 | 0 | 2 | 2 | 2/2 (100) |
| HLA‐C15:02_P3 | CoV_Prot_S_93‐107 | CoV_Prot_S_81‐130 | HLA‐C15:02 | ASTEKSNII | 3 | 0 | 0 | 0 | 0 | 0/3 | 3 | 1 | 0 | 1 | 2 | 2/3 (66,67) |
| HLA‐C16:01_DRB1_1501_P1 | CoV_Prot_S_154‐168 | CoV_Prot_S_121‐175 | HLA‐C16:01 | YSSANNCTF | 1 | 0 | 0 | 0 | 0 | 0/1 | 1 | 0 | 0 | 0 | 0 | 0/1 |
| HLA‐C16:01_DRB1_1501_P1 | CoV_Prot_S_154‐168 | CoV_Prot_S_121‐175 | DRB1_1501 | YSSANNCTF | 5 | 0 | 0 | 1 | 1 | 1/5 (20) | 6 | 0 | 0 | 0 | 0 | 0/6 |
- —North Rhine‐Westphalia
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsSARS-CoV-2 and COVID-19 Research · vaccines and immunoinformatics approaches · COVID-19 Clinical Research Studies
INTRODUCTION
1
There are two main approaches to predict T‐cell epitopes, and thereby strength and specificity. One approach is the identification and mapping of T‐cell epitopes, which are presented via major histocompatibility complex (MHC) class I and class II molecules. Such peptides have average lengths of 8 to 11 amino acids and 13 to 18 amino acids for MHC class I and MHC class II, respectively [1]. There are two main approaches to predict T‐cell epitopes. One is the in silico analysis of peptide binding to specific HLA molecules via software tools such as NetMHCpan. The other is the in vitro stimulation of T cells with peptides of interest and the correlation of an individual's HLA haplotype (i.e., the sum of all HLA alleles of that individual) with the T‐cell response. Feeding into both approaches, all potential peptides derived from an antigen from the N‐ to the C‐terminal part of the protein (referred to as sequential walk) provide information about the immunogenic regions of that specific antigen.
The underlying reasons for severe COVID‐19 disease courses are poorly understood. Accumulation of virus‐specific CD4^+^ T cells was reported in severe cases [2, 3, 4] and increased production of IFN‐γ by CD8^+^ T cells and a higher degree of T‐cell clonal expansion was described to be favorable for moderate COVID‐19 [5]. Additionally, the presence of bystander T‐cell activation in early anti‐viral defense was demonstrated in mild early‐onset disease [6], while elevated circulating plasmablasts and reduced germinal center responses were repeatedly observed in severe COVID‐19 [7]. To which extent the general strength of the T‐cell response might be dampened by the bystander T‐cell activation and its correlation with disease severity is still not fully understood. Since the underlying mechanisms leading to severe COVID‐19 infections are yet to be determined, the identification of individuals with a higher risk for severe disease is pending. Little work has been done to systematically associate peptide presentation with the severity of the disease and to compare the potential of HLA haplotypes from severe and mildly infected people with each other. Both analyses could support the identification of reliable clinical prediction markers.
In addition, novel virus strains can dampen the T‐cell responses gained via vaccination with first‐generation vaccines or natural infection with pre‐VOC viral strains. Certain strains are even able to completely escape it if they are presented in the context of certain HLA molecules [8, 9]. The extent to which this is supported by a dampened peptide presentation in different individuals or whether certain HLA molecules are predestined for this viral escape is still controversial. It is common understanding to correlate known T‐cell epitopes with areas undergoing high mutation rates in order to make assumptions about the degree to which T‐cell responses will be potentially diminished in the future.
In this study, T‐cell epitopes were identified and the T‐cell epitope landscape within various SARS‐CoV‐2 proteins was determined using groups of vaccinated and/or convalescent individuals. As these immunogenic protein regions did not overlap with areas, which show enhanced mutation rates, a relatively stable T‐cell response is expected toward upcoming virus strains. Furthermore, we could not find evidence that peptide presentation capacity from the immunogenic regions of HLA haplotypes from severe and mild diseases are strongly different. However, in line with previous studies [7], in long and more severe disease courses a bystander activation of unspecific CD8^+^ T cells, represented by waning frequencies of virus‐reactive CD8^+^ T cells, accompanied by significantly increased cell counts and a strong CD4^+^ T‐cell response were detected. Thus, we conclude that differences in disease courses (duration and severity) were likely not induced by the ability of certain HLA molecules to present specific peptides and to induce T‐cell responses, but rather the specificity of the T‐cell response directly.
Taken together, we suggest that the T‐cell response is likely to remain relatively stable in future SARS‐CoV‐2 virus strains. In addition, the main cause of severe disease courses is rather the result of intrinsic differences in T‐cell receptor (TCR)/HLA‐induced signaling and activation.
RESULTS
2
Composition of Cohort Study Groups and Epitope Analysis Strategy
2.1
To determine the T‐cell epitope landscape, three cohort groups were analyzed: (A) vaccinated (n = 27), (B) convalescent (n = 33), and (C) convalescent vaccinated (n = 12) study subjects (Figure 1). For all study subjects background information is provided (Figure 1A,B; Figure S1, Table S1).
Experimental design and cohort description. (A) For cohorts A, B, and C, the cohort‐specific SARS‐CoV‐2 background, including the number of participants, the sex distribution ratio (female:male), the mean age of the participants, the interim days between vaccination or infection to the date of the blood donation as well as the administered vaccines are listed (B) Exemplary distribution of HLA‐A allotypes for cohorts A, B, and C. Shown is the frequency of each allele amongst the total number of HLA‐A allotypes expressed within the cohort. (C) Given the protein sequence of the SARS‐CoV‐2 spike, nucleocapsid, and membrane proteins, 53 peptide pools covering approximately 50 amino acid‐long, overlapping sequences were designed (top). These peptide pools include 10 consecutive 15‐mer peptides, with an 11 amino acid overlap to each other. Vaccinated (cohort A), convalescent (cohort B) as well as vaccinated and convalescent individuals (cohort C) were recruited to assess (I) the CD4+ and CD8+ T‐cell response against these aforementioned 50‐mer sequences, and (IIa) the CD4+ and CD8+ T‐cell response against 15‐mer peptides that are derived from immunogenic peptide pools by conducting in vitro stimulation approaches. Furthermore, (IIb) the binding affinity of peptides to MHC Class I and II molecules were in silico predicted using NetMHCpan with regard to the HLA background of the individual study subjects. Lastly, (III) T‐cell responses from core peptides were validated in vitro. Study subjects of cohort D, comprising independent convalescent individuals, were HLA phenotyped. The frequency as well as absolute number of SARS‐CoV‐2 responsive T cells, neutralizing antibody (nAB) titers, and their clinical phenotype were assessed. All parameters were included in correlation analyses.
We established an experimental workflow comprised of three analysis levels (Figure 1C, bottom left graphic): On the first level, T cells were stimulated in vitro. For this purpose, 53 peptide pools were used, and all pools together covered the entire S, N, and M antigens from their N‐ to the C‐termini. They comprised consecutive peptides, mostly 15‐mers which had 11 amino acids overlap with each other. Each single peptide pool covered a 50 amino acid‐long region of the indicated SARS‐CoV‐2 antigens (Figure 1C, “I”). This first analysis level provided information on whether and which 50‐amino acids long region were immunogenic and, thus, could mount a T‐cell response (Table 1). On the second analysis level, the available 15‐mer peptides for the previously identified immunogenic 50‐amino‐acid‐long regions were used for T‐cell stimulation to further narrow down a potential T‐cell epitope. In addition on the second level, an in silico, prediction of potential MHC‐binders was conducted using the software NetMHC(II)pan and correlated with in vitro findings (Figure 1C, “IIa/IIb”). Therewith, we determined theoretical binding affinities of 15mer single peptides to the HLA alleles of interest. Simultaneously the accompanying 9‐mer MHC binding core peptide sequences were defined.
The latter is verified on the third and last analysis level. Here, 9‐mer immunogenic core peptides, within 15‐mer verified and predicted immunogenic peptides (Tables 2 and 3), were in vitro validated for their potential to induce T‐cell responses, and NetMHC(II)pan‐predicted MHC restrictions were confirmed or refuted (Figure 1C, “III”; Table 4). An exemplary gating strategy or the flow cytometric identification of T‐cell reactivity upon stimulation with a 9‐mer core peptide (HLA‐A01:01_P2) and SARS‐CoV‐2 peptide pools (SARS‐CoV‐2 PepTivator S/M/N) is provided in Figure S2.
Identification of Immunogenic Protein Regions and Peptides
2.2
To identify SARS‐CoV‐2‐specific immunity gained through vaccination or infection, the absolute numbers of study subjects, who showed T‐cell responses to the indicated 50‐amino‐acid‐long regions of the SARS‐CoV‐2 S, M, and N proteins were counted (Figure 2). T‐cell reactivity was measured based on CD8^+^IFN‐γ^+^TNF‐α^+^ T cells (from now on referred to as “reactive CD8^+^ T cells”) (Figure 2B, upper graph) and CD4^+^CD154^+^TNF‐α^+^ T cells (from now on referred to as “reactive CD4^+^ T cells”) (Figure 2B, lower graph). Cohort C (red bars) was negligible due to low n numbers and little overlap in HLA distributions to cohorts A and B (Figure 1B). Comparison of cohort A (orange bars) and B (petrol bars) unveiled differences between vaccination and infection: vaccinated individuals showed peaks of reactivity within the mid‐region of the S1‐subunit for both, reactive CD4^+^ and CD8^+^ T cells, and at the N‐terminal region of the S1‐receptor binding domain (RBD) and S2‐subunit for reactive CD8^+^ T cells. In contrast, convalescent individuals displayed a generally more homogeneously distributed T‐cell reactivity along the S, N, and M proteins among CD8^+^‐reactive T cells. This was slightly different for reactive CD4^+^ T cells, as here convalescent individuals display a partial lack of reactivity within the S1 RBD subunit, as well as a peaking region of reactivity at the C‐terminus of the Membrane protein.
Immunogenic protein regions within the SARS‐CoV‐2 spike‐, nucleocapsid‐, and membrane‐protein among cohorts A, B, and C. (A) Illustration of SARS‐CoV‐2 Spike (S), Nucleocapsid (N) and Membrane (M) protein sequences. Given are the total numbers of amino acids covered by the protein (subunits) as well as their respective positions. (B) Bar graphs (aligned to (A)) for cohort A (vaccinated; orange), cohort B (convalescent; petrol) and cohort C (vaccinated and convalescent; red) showing the absolute numbers of reactive study subjects amongst CD8+TNF‐α+IFN‐γ+ (upper graphs) and CD4+CD154+TNF‐α+ T cells (lower graphs) (left y‐axes) upon stimulation with peptide pools covering the Spike protein (left graphs), the nucleocapsid protein (middle graphs) and the Membrane protein (right graphs), plotted against the respective protein region (x‐axes). Aligned to these bars, the absolute number of mutations that are found within the SARS‐CoV‐2 B.1.1.529/Omicron sublineages E.G.1, XBB, BQ.1, BA.2.12.1, BA.5, BA.4 and BA.1 (pink curve; left y‐axes) is shown.
To uncover whether virus mutations would occur preferentially in regions that could mount a T‐cell response, the number of mutations emerging from the B.1.1.529/Omicron virus strain lineages (EG.5.1, XBB, BQ.1, BA.2.75, BA.2.12.1, BA.5, BA.4, and BA.1) within the respective protein regions (Figure 2B, left y‐axes, pink lines) was aligned. Only partial overlap with the identified regions of T‐cell reactivities was observed.
Taken together, T‐cell responses in a cohort of convalescent individuals were mostly distributed over the entire sequences of the S, M, and N proteins, while vaccinated individuals rather reacted to distinct regions of the S protein. Our analyses provided a comprehensive overview of the epitope landscape of the S, M, and N protein of SARS‐Cov‐2 and identified certain differences between vaccinated and convalescent individuals. Recent mutations in SARS‐CoV‐2 did not indicate preferential mutations in regions capable of mounting strong T‐cell responses. Furthermore, a considerable number of potential MHC‐binding peptides were predicted and detected in all antigens. Hence, we conclude that T‐cell responses ought to be rather resilient to viral variants with the current rate of mutation of SARS‐CoV‐2 and a T‐cell escape variant affecting a general public would require a major reorganization of the viral protein structure, which is in line with previous studies [10]
(Un)favorable Haplotypes Do Not Influence HLAs’ Peptide Presentation Capacity
2.3
Fischer et al. [11] recently correlated certain HLA haplotypes with a longer disease duration and enhanced severity. To uncover whether the cause of a severe disease course lies in an impaired potential of the HLAs to present SARS‐CoV‐2 S, M, and N‐derived peptides, an independent cohort of convalescent donors, the so‐called cohort D, was recruited (Figure 1C). Cohort D was included to represent the first and second line of infections (“Ischgl‐cohort”). Therefore, this cohort is a cohort, with known immune and infection status, as well as disease status. Moreover, the patients are young and do not have co‐morbidities as they are potential convalescent blood donors. These individuals were sequenced for their HLA haplotypes and assessed for their cellular and humoral immune responses. Thereafter, the identified HLA haplotypes expressed by the individuals of cohort D were correlated to the individual disease duration and severity, which was assessed based on a self‐questionnaire (see extract in the M&M part). This correlation and the previous study by Fischer et al. [11] including a much bigger cohort, allowed a classification of the HLA haplotypes into two major groups: favorable and unfavorable haplotypes. The two groups were classified by calculating a hazard score for all individuals of cohort D based on their disease course (Table S3 and S4).
Having identified those haplotypes that are linked to mild and more severe COVID‐19 disease (Table S5), individuals from cohorts A to C were re‐grouped according to their expression of favorable‐ and unfavorable‐HLA haplotypes and subsequently re‐analyzed to see whether those HLAs elicit an altered T‐cell‐response pattern. However, the resulting T‐cell immunogenic protein regions were comparable for individuals of all cohorts and both subgroups (Figure S3). This assertion is applicable to both, reactive CD8^+^ and CD4^+^ T cells, pointing toward a similar MHC‐mediated peptide presentation capacity within both groups of individuals. A reciprocal analysis comparing the number of presented epitopes in people, who underwent severe or mild disease courses only revealed minimal differences, with significantly higher peptide responses for the more favorable MHC II haplotypes (Figure S4), providing further evidence for similar peptide presentation potentials in mild and severe diseases.
HLA Haplotypes Linked to Unfavorable Disease Courses Correlate With Reduced CD8+ T‐Cell Responses
2.4
As we did not find indications pointing to differences in peptide‐presentation capacity between individuals expressing favorable and unfavorable HLA alleles for cohorts A to C, we hypothesized that T‐cell intrinsic differences and varying strengths of the T‐cell responses were the underlying cause of varying disease courses. Hence, cellular responses in patients with favorable and unfavorable HLA haplotypes were analyzed for cohort D and the frequencies of CD3^+^CD8^+^INF‐γ^+^ as well as CD3^+^CD4^+^INF‐γ^+^ effector memory T cells were assessed and correlated with the individual disease duration or severity. This analysis revealed that patients with an unfavorable HLA‐haplotype showed lower frequencies of INF‐γ‐expressing SARS‐CoV‐2‐specific CD8^+^ T cells (with around 3%), although these patients suffered longer from COVID‐19 (Figure 3A,B). Noteworthy, blood withdrawal was always around the same time point (see material and methods section). Simultaneously, the same group suffering from long disease duration presented with high frequencies of INF‐γ‐expressing SARS‐CoV‐2‐specific CD4^+^ T cells (with up to approx. 28%), (Figure 4A,B). However, when the study subjects were not specifically categorized into favorable or unfavorable HLA‐haplotypes, disease duration as well as disease severity aligned with an increased number of CD3^+^CD8^+^INF‐γ^+^ as well as CD3^+^CD4^+^INF‐γ^+^ antigen‐specific T cells for all SARS‐CoV‐2 proteins (S, M, and N) after re‐stimulation (Figures 3C and 4C). Considering only the unfavorable HLA‐haplotypes for all disease stages, a significant increase in the absolute number of CD3^+^CD8^+^INFγ^+^ T cells was identified for those individuals that suffered from a long disease duration (Figures 3D and 4D), pointing toward a bystander T‐cell activation [17].
*Long disease courses are dominated by a weaker virus‐specific CD8+ T‐cell response and unfavorable HLA‐haplotypes. Correlation of disease duration with the frequency of PepTivator‐specific CD3+CD8+IFNγ+ T cells of (A) study subjects with mild (yellow), moderate (green), short (blue), and long disease (red) (n ≥9/group) and (B) all study subjects (n = 40) show an accumulation of unfavorable HLA haplotypes and low frequencies of reactive CD8+ T cells for prolonged disease courses. White spheres indicate unfavorable HLA‐haplotypes, while blue spheres represent favorable HLA‐haplotypes for disease severity Frequencies were determined within PBMCs, gated on CD45, CD3, CD8, and the expression of IFN‐γ. (C) Flow cytometric assessment of the total cell count of virus‐reactive CD3+CD8+ T cells after stimulation with the three different SARS‐CoV‐2 PepTivators S, M, and N as well as a combination of all shows significantly increased numbers of reactive CD8+ T cells. (D) Assessment of the total cell count of virus‐reactive CD3+CD8+ T cells of study subjects bearing exclusively unfavorable (left) or favorable (right) HLA‐haplotypes after combinatorial restimulation with SARS‐CoV‐2‐PepTivator pool S, M and N shows significant differences for individuals with unfavorable HLA haplotypes only. p < 0.05; Error bars represent mean values ± SEM; Mann–Whitney U‐test.
*Long disease courses show higher frequencies of virus‐specific CD4+ T cells and unfavorable HLA‐haplotypes. (A) Correlation of disease duration with the frequency of PepTivator‐specific CD3+CD4+IFNγ+ T cells of (A) study subjects with mild (yellow), moderate (green), short (blue), and long disease (red) (n ≥9/group) and (B) all study subjects (n = 40) show an accumulation of unfavorable HLA haplotypes and a tendency toward elevated frequencies of reactive CD4+ T cells for prolonged disease courses. White spheres indicate unfavorable HLA haplotypes, while blue spheres represent favorable HLA haplotypes for disease severity. Frequencies were determined within PBMCs, gated on CD45, CD3, CD8, and the expression of IFN‐γ. (C) Flow cytometric assessment of the total cell count of virus‐reactive CD3+CD4+ T cells after stimulation with the three different SARS‐CoV‐2 PepTivators S, M, and N as well as a combination of all, shows only minor differences in reactive CD4+ T cell counts. (D) Assessment of the total cell count of virus‐reactive CD3+CD4+ T cells of study subjects bearing exclusively unfavorable (left) or favorable (right) HLA‐haplotypes after combinatorial restimulation with SARS‐CoV‐2‐PepTivator pool S, M and N demonstrates no significant differences. p < 0.05; Error bars represent mean values ± SEM; Mann–Whitney U‐test.
HLA‐alleles Correlated With Disease Duration and CD4+ T‐Cell Response
2.5
Although the unfavorable HLA‐haplotypes did not show a direct impact on CD4^+^ T‐cell responses within cohort D, the question was raised whether other parameters might play a role. Therefore, we looked at the age, and expression of CCR5, a chemokine receptor that is crucial for the humoral response of mucosal immunity [11], as well as the humoral response itself. There were no differences within cohort D under consideration of age or the expression of CCR5. As virus‐specific CD4^+^ T helper‐cell responses were elevated during long COVID‐19 disease courses within the study subjects of cohort D, we assessed their neutralizing antibody (nAB) titers. Titers of ≥1:160 were considered as a positive reaction against SARS‐CoV‐2 infections. Here we correlated the CD4^+^ T‐cell response of all study subjects with their individual disease duration and the associated HLA‐haplotypes, showing that those with the unfavorable HLA‐haplotypes presented also with the highest counts of CD4^+^ T cells (Figure 5A). Considering the level of nABs in correlation to the haplotypes, those individuals with unfavorable haplotypes correlated also with the highest titers of neutralizing antibodies (Figure 5B).
*Unfavorable HLA‐haplotypes correlate with a higher CD4+ T‐cell response and elevated levels of neutralizing antibodies. (A) Correlation of disease duration with the CD4+ T‐cell response shows positive association with unfavorable HLA‐haplotypes. (B) Positive correlation of disease duration with the titer of neutralizing antibodies (nABs) within unfavorable HLA‐haplotypes. 0 corresponds to the median response of unfavorable and favorable haplotypes that were linked to disease duration; *p < 0.05, **p < 0.005, **p < 0.0005. Error bars represent mean values ± SEM.
DISCUSSION
3
Mapping of Immunogenic Protein Regions Reveals Distinct Patterns for Vaccinated and Naturally Infected Individuals
3.1
Virus‐specific T‐cell responses are linked to fast viral clearance and mild disease courses. Improving our understanding of SARS‐CoV‐2‐specific T‐cell responses is crucial for the ability to treat and prevent severe disease courses. The present study provides a comprehensive overview of the epitope landscape of the N, M, and S proteins of SARS‐CoV‐2.
Dörnte et al. could previously show via both in vitro peptide stimulation and in silico NetMHCpan analyses that SARS‐CoV‐2 mutations may not lead to a complete escape from the T‐cell response on a population level. Since December 2020 [12], multiple vaccines have been developed to protect the population from severe infections [13]. However, through the continuous mutation of SARS‐CoV‐2 antigens novel virus strains arise with altered pathogenicity, as these strains bear the potential to escape SARS‐CoV‐2 specific immunity gained via vaccination with first‐generation vaccines or via natural infection with ancestral virus‐strains [1]. Therefore, predicting the effect of mutations on T cell‐mediated immune responses is important to estimate the potential loss of immunoprotection, for example, caused by a novel heavily mutated SARS‐CoV‐2 strain.
The present study demonstrates that vaccinated individuals have a tendency to mount T‐cell responses to slightly more distinct regions of the Spike protein, while T‐cell responses in naturally infected individuals were induced via more uniformly distributed regions of the S protein. Note, that here T‐cell responses have been evaluated independently from the individual response strength, and that the number of T‐cell epitopes might not correlate with the latter. This uniform distribution of protein regions that are recognized by T cells of convalescent individuals could also be observed for the M and the N proteins. Notably, this difference between vaccinated and convalescent individuals was especially pronounced within the CD4^+^ T‐cell compartment. Whether CD4^+^ T‐cell activation could directly affect the quantity and quality of the humoral immune response remains to be investigated, since follicular T helper (T_FH_) cells were not analyzed. However, the direct correlations between CD4^+^ T‐cell and humoral responses have been described [14, 15]. Therefore, a lack of CD4^+^ T cells that specifically recognize cognate peptides presented by the pre‐activated B cells may result in an impaired formation of long‐lived memory B cells. The broad and more evenly distributed T‐cell response presented by convalescent individuals might support an improved B‐cell response and memory phenotype compared with the immunity gained by vaccinated individuals, but as mentioned above differences between these two groups were rather mild, and an in‐depth T_FH_ analysis would be required.
Both the in silico prediction of potential MHC binders and the alignment of mutations found in the S, M, and N proteins suggest relative stability of the T‐cell response also for novel virus strains unless the protein structure of SARS‐CoV‐2 undergoes a major reorganization. This hypothesis is based on (I) the high number of potential MHC binding peptides predicted throughout the entire sequence of the Spike protein [16, 17], (II) the evenly distributed areas, which can mount T‐cell responses in naturally infected individuals, (III) the high number of 15‐mer and core peptides identified in the present study and other previously published literature [18, 19, 20, 21, 22, 23], and (IV) the absence of a correlation of past SARS‐CoV‐2 mutations with areas mounting strong T‐cell responses. This hypothesis is further supported by previously described findings, which demonstrated stable T‐cell responses to the B.1.1.529‐variant of SARS‐CoV‐2 in individuals vaccinated two or three times with first‐generation vaccines against all evaluated proteins [24].
To determine whether an impaired capacity of MHC alleles to present peptides to T cells was associated with more severe and prolonged disease, we reevaluated the T cell‐immunogenic protein regions of cohorts A to C. This analysis revealed that there is no difference between the T‐cell recognition patterns of individuals, expressing favorable or unfavorable HLA alleles. Based on that we evaluated the T‐cell response strength in an additional cohort D, for which clinical parameters for disease length and severity were available, which allowed us to perform additional correlations. To exclude the possibility of an impaired recognition of pMHC‐complexes by T cells future subsequent studies could comprise TCR avidity evaluations. Considering the T‐cell response of the naturally infected individuals categorized based on their disease duration and severity we identified a clear pattern for CD8^+^ T‐cell responses. Those individuals with a short disease duration mounted the highest frequencies of CD8^+^ T cells while those with a prolonged disease course had rather low frequencies of virus‐specific CD8^+^ T cells. Contrastingly, we observed high numbers of reactive, virus‐specific CD8^+^ T cells after re‐stimulation with SARS‐CoV‐2 derived peptides although the same cell numbers were applied for the assays, which points toward a high bystander activation of CD8^+^ T cells in severe COVID‐19. Additionally, comparing those observations to the T‐cell responses assessed for vaccinated‐only patients, the virus‐specific T cells generated under infection were directed against all proteins of the SARS‐CoV‐2 strain (S, M, and N).
This data indicates a lack of a virus‐specific MHC class I response in those patients with more severe symptoms, which is also supported by the literature [25, 26, 27]. Assessment of the virus‐specific CD4^+^ T‐cell response reveals an equal distribution in all four different subgroups of convalescent subjects, with elevated frequencies only in a combined cocktail of all proteins pointing toward a rather CD4‐mediated immune response in those patients who suffered longer. These findings are in line with the results found for neutralizing antibody titers, where those patients with a higher CD4^+^ T‐cell response also aligned with higher antibody titers, indicating a likely CD4^+^ T‐cell linked B‐cell response as described above. However, further characterization of CD4^+^ T‐cell subsets is required in the future.
Several publications have shown that infection with SARS‐CoV‐2 is, indeed, linked to the expression levels of HLA molecules. The group of Bahlmann et al. transfected two human cell lines, Caco‐2 (human colorectal adenocarcinoma cells) and Calu‐3 (human airway epithelial cells) with different SARS‐Cov2 virus strains and showed, that the virus‐positive cells expressed a significantly lower number of HLA‐molecules than nontransfected cells, indicating that SARS‐CoV‐2 inhibits the induction of the HLA class I pathway [28]. In a cohort of vaccinated patients, Mentzer et al. analyzed the humoral response by determining the antibody titers after vaccination and associated the response with the HLA class I pathway as they found a clear association with genes regulated on Chromosome 6, where the gene for HLA is located. Moreover, they identified that patients with HLA‐DQB1:06 presented with the highest antibody titers [29].
The correlation of the antibody titers with the severity of the disease revealed that while IgG titers against the Spike protein or the Nucleocapsid were clearly associated with the course of the disease, as neutralizing antibodies were only highly produced by those patients with a rather moderate/long infection (WHO grade 3) [30]. A more detailed study performed by Lehmann et al. investigated a so‐called “house‐cohort” in which some individuals of the same household underwent COVID‐19 infection, while others did not. They identified that those patients that got infected were homozygous for specific HLA molecules. Looking for those HLA‐molecules in convalescent study subjects of cohort D, carrying HLA‐DQB05:01 was considered a “protective” HLA‐molecule, resulting in a shorter disease duration, while those lacking this molecule suffered from a longer infection [31]. Moreover, individuals with HLA‐DQB05:01 expression had lower IgG antibody titers compared with those without, but there was no difference in the neutralizing antibody titers, indicating that carriers of HLA‐DQB*05:01 have a tendency to form antibodies [32].
Considering another parameter, that might play a role in the differing immune responses in cohort D, the cohort was divided into male and female donors to assess the effect of the sex on the cellular and humoral immune response. It is known that men generally develop slightly dominating Th1‐driven immune responses, while female responses are dominated by Th2‐cells [33]. Moreover, it has been reported that women mount much higher antibody levels due to more proliferative B cells [33]. An effect that was mirrored in the results of convalescent study subjects from cohort D. Through breaking down the HLA‐haplotypes to the most common alleles and looking at the duration of the SARS‐CoV‐2 infection, in females HLA class I haplotypes were associated with shorter, while HLA class II are associated with longer disease duration. Contrastingly, in males, HLA class II is associated with a shorter disease duration. Hence, a shorter disease duration is associated with a more favorable HLA‐haplotype (Table S3) [32]. These results are in line with the cellular response, in which unfavorable HLA‐haplotypes show a weaker CD8^+^ T‐cell response (Figure 3), while there was no association with CD4^+^ T cells (Figure 4). However, considering the disease duration, CD4^+^ T cells were elevated and were associated with elevated neutralizing antibody titers (Figures 4 and 5).
The results presented in this study underline the requirement of a proper cellular response, which is mandatory to clear the infection, thereby also supporting previous publications [34]. A strong T‐helper cell link within more severe and longer infections, which support the B cells in producing higher antibody titers is likely, however, requires further corroboration by future studies, in particular, the analysis of the T_FH_ subset. In addition, despite a relatively stable T‐cell response toward novel SARS‐CoV‐2 strains is expected, next‐generation vaccines should in the future aim for preserving a sufficient cellular T‐cell response and therefore should specifically include T‐cell antigens in addition to the formation of antibodies against SARS‐CoV‐2.
Limitations of the Study
3.2
-
Cohorts A, B, and C include nonsimilar numbers of participants as cohort C is considerably smaller than A and B (n A = 27, n B = 33, n c = 12). Therefore, a direct comparison of obtained T‐cell reactivity patterns with cohort C was not done in this study, although it has no effect on the information about epitope candidates and core peptides obtained from the analysis of cohort C.
-
Cohort A comprises 3.5 times more females compared with males. Hence, a discrimination between T‐cell responses in male and female individuals for cohorts A, B, and C was not performed.
-
The mean interval between vaccination and/or natural infection and the time of blood collection is not the same between the cohorts with a median of 50 ± 9 days. Furthermore, the majority (63%) of cohort A received BNT162b2 vaccines, while B was more diverse. Correlation of T‐cell responses with vaccine type and time since vaccination and/or infection was not possible.
-
Differences between HLA allotypes expressed by each cohort were observed for all HLA class I and II genes. Although these differences are inevitable due to the natural diversity of the human HLA gene locus, unequal T‐cell reactivity against SARS‐CoV‐2 structural proteins may be influenced by the different peptide binding affinities of the expressed HLAs rather than only by the immunization routes represented by the different cohorts.
The evaluation of intrinsic differences in T‐cell response strengths was done on convalescent individuals with mild to moderate infections and no severe infections. Nevertheless, our results are in line with studies including severe outcomes. A deeper characterization of T_FH_ cells as a potential link between the CD4^+^ T cell immune response and antibody‐producing B cells is necessary. This could be the focus of future studies.
Materials and Methods
4
Study Participants and Whole Blood Donations
4.1
Study participants were recruited at Miltenyi Biotec via an internal blood donation program, and the Medical School Hannover (MHH) (cohort A–C), as well as from volunteers of the University Clinic Düsseldorf (cohort D). The individual SARS‐CoV‐2 background of all study participants was assessed via questionnaires and summarized in Table S1. Participants 1 to 73 were assigned to the study's cohorts comprising (A) vaccinated, (B) convalescent, and (C) vaccinated and convalescent individuals (Figure 1C). Whole blood donations were drawn from all volunteers after informed consent (Miltenyi Biotec: 20200151; Medical School Hannover: 9255_BO_K2020; University Clinic Düsseldorf) and following the WMA Declaration of Helsinki regarding the ethical principles for medical research involving human subjects. Therefore, 100 mL whole blood was drawn using an EDTA‐coated BD Vacutainer (BD, Franklin Lakes, USA, cat. no. 367525). Donations from the MHH hospital were transported overnight to Miltenyi Biotec. Donations from Miltenyi employees as well as from volunteers of the University Clinic Düsseldorf were directly processed after blood drawing. There was a mean of 57 days for cohort A, a mean of 59 days for cohort B, and a mean of 41 days for cohort C between the date of vaccination or infection and the date of sample drawing. For cohort D, patients donated their blood samples as convalescent donors within 2 months after recovery.
It is important to note that study participants of cohort D were recruited and analyzed independently from cohorts A to C, for the evaluation of SARS‐CoV‐2–specific T‐cell response strengths of the 40 donors depicted in this study. The categorization of those study participants into subgroups of mild, moderate, as well as short and long disease was done according to information assessed by a self‐questionnaire. Hereby, the different disease durations/strengths were subdivided based on disease duration and symptoms (mild: sore throat, running nose, no fever/ short: 1–10 days/moderate: sore throat, running nose, fever over 3 days/long: 20–120 days). We insert here an extract from the questionnaire for recording the degree of illness:
Sequential Walk
4.2
To identify SARS‐CoV‐2 specific T‐cell epitopes an in vitro stimulation approach was utilized to assess the immunogenicity of SARS‐CoV‐2 derived peptides (for sequences for SARS‐CoV‐2 peptides see GenBank QHD43416.1, QHD43423.2, QHD43419.1). By using an experimental design aiming for a stepwise downsizing/filtering of potential immunogenic regions, the number of necessary experiments and blood material could be reduced. Hence, in the first step, 15mer‐peptide pools, followed by a downstream assessment of single, 15mer peptides out of reactive pools, and finally the evaluation of 9mer core peptides derived from reactive single peptides have been evaluated for their individual potential to initiate a peptide‐specific T‐cell response.
In more detail, first, a sequential walk over the virus’ spike, nucleocapsid, and membrane protein was performed using mostly 15mer consecutive peptides having 11 amino acids overlapping each other. Due to a limited number of available peripheral blood mononuclear cells (PBMCs), this first peptide stimulation approach was done using pools, comprising 10 to 12 peptides. Together 36 peptide pools (362 single 15mer peptides) for the Spike, 11 peptide pools (110 single 15mer peptides) for the Nucleocapsid, and 6 peptide pools (60 single 15mer peptides) representing the Membrane protein cover the complete sequence of the respective protein. After evaluation of immunogenic peptide regions covered by the tested pool, the respective single peptides’ immunogenicity was determined following the similar in vitro stimulation procedure, on expanded PBMCs of the same study subject. From an additional in silico, NetMHC (II)pan‐based analysis of observations from these first two in vitro approaches, the peptides’ HLA restrictions and core peptide sequences could be extracted and used in a third, and final validation of core peptides’ immunogenicity for certain HLA alleles.
For each of the described stimulation experiments, an unstimulated control, as well as two positive controls stimulated with CytoStim (Miltenyi Biotec, cat. no. 130‐092‐172) and SARS‐CoV‐2 PepTivators (PepTivator SARS‐CoV‐2 Prot_S Complete, cat. no. 130‐127‐951; PepTivator SARS‐CoV‐2 Prot_M, cat. no 130‐126‐709; PepTivator SARS‐CoV‐2 Prot_N, cat. no 130‐126‐698) covering the complete sequence of the before mentioned structural proteins, were prepared.
Isolation of Peripheral Blood Mononuclear Cells
4.3
To isolate the PBMCs from whole blood, density gradient centrifugation using Pancoll (Pan Biotec, Aidenbach, Germany, cat. no. P04‐60500) was utilized according to the manufacturer's instructions using CliniMACS PBS/EDTA buffer (Miltenyi Biotec, Bergisch Gladbach, Germany, cat. no. 200‐070‐025). To complete the isolation procedure, cells collected in a 50 mL Falcon tube were diluted with CliniMACS PBS/EDTA buffer and centrifuged twice at 200g for 15 min to remove the remaining thrombocytes. Afterward, cells were counted using the Sysmex XP‐300 device (Sysmex, Norderstedt, Germany) and diluted to a concentration of 1 × 10^7^ cells/mL using RPMI‐1640 Medium (Biowest, Nuaillé, France, cat. no. L0501‐500) supplemented with 5% human AB serum (Capricorn, Ebsdorfergrund, Germany, cat. no. HUM‐3B, Lot. CP20‐3472) and 1× Gibco Anti‐Anti (Thermo Fisher Scientific, Waltham, USA, cat. no. 11580486) (from now on referred to as “supplemented medium”). Using this cell suspension, an appropriate number of wells of 96‐well flat bottom plate (Falcon, New York, USA, cat. no. 353072) were filled to a concentration of 1 × 10^6^ PBMCs per well. The plate was then incubated overnight at 37°C, 5% CO_2_.
In Vitro Stimulation Approach and Subsequent Staining of Activity Markers and Intracellular Cytokines
4.4
Each peptide (pool) was separately added at a final concentration of 1 µg/mL/peptide/1E6 PBMCs to the priorly prepared cell samples. Stimulation was done for a total of 6 h at 37°C, 5% CO_2._ After 2 h, 2 µg/mL Brefeldin A (Sigma‐Aldrich, St. Louis, USA, cat. no. B7651) was added to each well to prevent cells from cytokine secretion.
After complete stimulation, staining was performed in a 96‐well V‐bottom plate (Sigma‐Aldrich, cat. no. Z667234) into which cells were transferred after adding 100 µL PBS/EDTA (2 mM) buffer to each sample. Afterward, the plate was centrifuged at 300g for 5 min at room temperature (RT) and the supernatant was discarded. Next, cells were stained with Viobility 450/452 Fixable Dyes (Miltenyi Biotec, cat. 130‐109‐816), according to the manufacturer's instructions. To enable subsequent intracellular staining of cytokines and activation markers, cell fixation was performed using Inside Fix (Inside Stain Kit, Miltenyi Biotec, cat. no. 130‐090‐477), according to the manufacturer's instruction, followed by resuspension in 250 µL/well Inside Perm (Inside Stain Kit, Miltenyi Biotec, cat. no. 130‐090‐477) and centrifugation at 300g for 5 min at RT. Finally, cells were stained using the following antibody‐cocktail: anti‐CD3—APC (Miltenyi Biotec, cat. no. 130‐113‐135), anti‐CD14—VioBlue (Miltenyi Biotec, cat. no. 130‐110‐525), anti‐CD20—VioBlue (Miltenyi Biotec, cat. no. 130‐111‐531), anti‐CD4—VioBright515 (Miltenyi Biotec, cat. no. 130‐114‐535), anti‐CD8—VioGreen (Miltenyi Biotec, cat. no. 130‐110‐684), anti‐IFN‐γ—PE (Miltenyi Biotec, cat. no. 130‐113‐496), anti‐TNF‐α—PEVio770 (Miltenyi Biotec, cat. no. 130‐120‐492), anti‐CD154—APCVio770 (Miltenyi Biotec, Cat. No.130‐114‐130), and anti‐IL‐2—PEVio615 (Miltenyi Biotec, cat. no.130‐111‐307). All antibodies were used in a 1:50 dilution. Staining was done according to the manufacturer's instructions. After additional washing with 100 µL Inside Perm and centrifugation at 300g for 10 min at RT, cells are resuspendedin250 µL PBS/EDTA (2 mM) buffer supplemented with 0.5% of MACS BSA Stock Solution (Miltenyi Biotec, cat. no. 130‐091‐376). Flow cytometric analysis was done using the MACSQuant16—flow cytometer (Miltenyi Biotec, cat. no. 130‐109‐803). We defined a positive T‐cell response by the appearance of a distinct population for the chosen activity‐associated markers and a frequency increase of at least 10% with respect to the unstimulated control.
Expansion of SARS‐CoV‐2 Spike, Nucleocapsid, Membrane‐Specific Reactive T Cells
4.5
Freshly isolated PBMCs (see previous section) were plated out on a four‐well plate (Falcon, Cat. No.353046), at a concentration of 5E6 PBMCs/5 mL supplemented medium. To support directed cell proliferation, 100 U/mL IL‐2 (Miltenyi Biotec, cat. no. 130‐097‐745) as well as 1 µg/mL of either PepTivator SARS‐CoV‐2 Prot_S Complete, PepTivator SARS‐CoV‐2 Prot_M, or PepTivator SARS‐CoV‐2 Prot_N were added to the cell suspension. Afterward, the cells were incubated for 14 days at 37°C, 5% CO_2_. Meanwhile, the medium was refreshed every 2 to 3 days by gentle removal of half the medium volume and subsequent addition of the same volume‐supplemented medium together with 100 U/mL IL‐2.
In Silico Peptide Binding Prediction
4.6
Using an in silico approach, HLA restrictions, corresponding binding affinities, and core sequences of single 15mer peptides, derived from previously immunogenic peptide pools, were specified. For that, the open‐source NetMHCpan (https://services.healthtech.dtu.dk/service.php?NetMHCpan‐4.1) and NetMHCIIpan (https://services.healthtech.dtu.dk/service.php?NetMHCIIpan‐4.0) prediction‐algorithms were utilized, for interactions with MHC class I and MHC class II molecules, respectively. The eluted ligand percentile ranks (EL‐Rank) are used as a measure of the peptide binding affinity. Therein, EL‐Ranks <2 and >0.5 are considered weak binding peptides, and EL‐Ranks <0.5 are considered strong binding peptides to the respective MHC class I molecule. For MHC class II molecules, instead, EL‐Rank thresholds of <5 and >2 and <2 are applied for the categorization into weak and strong binding peptides, respectively.
Sequencing of HLA‐Alleles
4.7
To genotype the HLA alleles for all cohort participants from the University Clinic Düsseldorf (cohort D), an amplicon‐based NGS‐based approach was used. Six multiplex polymerase chain reactions (PCRs) were used to amplify exons 2, 3, and 4 of the HLA‐Class I and HLA‐DPB1 genes and exons 2 and 3 of HLA‐DRB1, HLA‐DQA1, and HLA‐DQB1. The resulting fragments were supplemented with sample‐specific barcodes and Illumina‐compatible adapters. Sequencing was performed on an Illumina MiSeq device (Illumina Inc., San Diego, USA). A customized software (NGSSequence Analyser, Institute for Transplantation Diagnostics and Cell Therapy, University Hospital Dusseldorf, Dusseldorf, Germany) served for the analysis of the sequence reads using quality control values and high coverage to automate the data analysis. For haplotype phasing of the obtained HLA data, we used the Arlequin software (version 3.5.2.2, available cmpg.unibe.ch/software/arlequin35/), for binding affinities to viral peptides [35].
For all remaining study subjects from other cohorts, HLA sequencing was performed externally by DKMS Life Science Lab GmbH.
Determination of Antibody Levels
4.8
Determination of antibody levels was performed using two serological assays as described before [36]. Spike S1 protein domain‐specific IgA and IgG antibodies were detected by ELISA (Euroimmun, Germany), and Ig antibodies against the Nucleocapsid were verified by Roche Diagnostic Test (Elecsys). Neutralizing antibodies were tested additionally in biological duplicates. Heat‐inactivated donor plasma was diluted. Serum neutralization titer was determined by microscopic inspection as the highest serum dilution without virus‐induced cytopathic effect (CPE). Two previously tested sera from SARS‐CoV‐2 infected individuals served as positive controls for each round. A high‐titer control (NT 1:640) and a medium‐titer control (NT 1:160) were defined to validate each assay. Interassay and intra‐assay variations were determined, and these control sera exhibited maximum variation within only one dilution level. Serum from noninfected individuals and cells without serum served as negative controls to confirm virus‐induced CPE. A neutralization titer of 1:160 was defined as the threshold titer for the binary representation of virus neutralization capacity, allowing the potential plasmapheresis product to serve as a therapeutic agent.
In Vitro Assays for Antigen‐Specific T Cells
4.9
PBMCs of convalescent subjects from four different sub‐groups of disease severity or duration (short, mild, moderate, long) were isolated as described above. The cell suspension was adjusted to a cell titer of 1 × 10^7^ cells/mL X‐Vivo 15 medium (Lonza; cat. no. 11695120) and 100 µL/well was distributed into a 96‐well plate. For each study subject, several approaches were set up: SARS‐CoV‐2 PepTivator Prot_S Complete, Prot_M, and Prot_N as well as a well for all three PepTivators, a negative control (no PepTivators) and the stimulation with PMA/Iono as a positive control. Cells were incubated with the according stimulating agents for 6 h in total. After 30 min of stimulation, 5 ng/mL Brefeldin A (BioLegend; ca. no. 420601) were added to each well. Subsequently, cells were harvested and stained for anti‐CD3‐PE‐Dazzle (BioLegend, cat. no. 300446), anti‐CD4 BV650 (BioLegend, cat. no. 317436), anti‐CD8 VioGreen (Miltenyi Biotec, cat. no. 130‐113‐164), anti‐CD45RO_BV570 (BioLegend, cat. no. 304226), anti‐CD154_PE‐Cy7 (BioLegend, cat. no. 310832) and anti‐IFN‐γ‐BV421 (BioLegend, cat. no. 502532). All antibodies were used in a 1:100 dilution. Staining was done according to the manufacturer's instructions. After washing with PBS and centrifugation at 300g, cells are resuspended in 250 µL PBS/EDTA (2 mM) buffer supplemented with 0.5% of MACS BSA Stock Solution (Miltenyi Biotec, cat. no. 130‐091‐376). Flow cytometric analysis was done using the CytoFlex S–flow cytometer (Beckmann Coulter). Each sample was then normalized to the negative control to exclude unspecific responses or backgrounds.
Author Contributions
Conceptualization: C. Dörnte, A. Datsi, J. Fischer, and M. Schuster. Sample acquisition: J. Fischer, J. Kostyra, C. Lamsfuß, H. Baurmann, B. Eiz‐Vesper, and A. Bonifacius. Investigation: C. Dörnte, A. Datsi, V. Traska, J. Kostyra, M. Wagner, O. Brauns, DS. Visualization: C. Dörnte and A. Datsi. Funding acquisition: H. Baurmann, A. Richter, J. Schmitz, and M. Schuster. Project administration: C. Dörnte, O. Brauns, H. Winkels, H. Baurmann, B. Eiz‐Vesper, A. Bonifacius, R. V. Sorg, C. Dose, J. Schmitz, A. Richter, J. Fischer, and M. Schuster. Supervision: M. Schuster, J. Fischer, and R. V. Sorg. Writing–original draft: C. Dörnte and A. Datsi. Writing–review & editing: C. Dörnte, A. Datsi, J. Fischer, R. V. Sorg, and M. Schuster.
Conflicts of Interest
C. D., V. T., J. K., J. H., M. W., O. B., D. S., C. L., H. B., C. Do., J. S., A. R., and M. S. are employees of Miltenyi Biotec B.V. & Co. KG. The remaining authors declare no conflicts of interest.
Peer Review
The peer review history for this article is available at https://publons.com/publon/10.1002/eji.202451497.
Supporting information
Supporting Information
Supporting Information
Supporting Information
Supporting Information
Supporting Information
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1C. Szeto , C. A. Lobos , A. T. Nguyen , and S. Gras , “TCR Recognition of Peptide–MHC‐I: Rule Makers and Breakers,” International Journal of Molecular Sciences 22, no. 1 (2021): 1–26, 10.3390/ijms 22010068.PMC 779352233374673 · doi ↗ · pubmed ↗
- 2A. Sette and S. Crotty , “Adaptive Immunity to SARS‐Co V‐2 and COVID‐19,” Cell 184, no. 4 (2021): 861–880, 10.1016/j.cell.2021.01.007.33497610 PMC 7803150 · doi ↗ · pubmed ↗
- 3A. T. Tan , M. Linster , C. W. Tan , et al., “Early Induction of Functional SARS‐Co V‐2‐specific T Cells Associates With Rapid Viral Clearance and Mild Disease in COVID‐19 Patients,” Cell reports 34, no. 6 (2021), 10.1016/j.celrep.2021.108728.PMC 782608433516277 · doi ↗ · pubmed ↗
- 4Rydyznski Moderbacher C, S. I. Ramirez , J. M. Dan , et al., “Antigen‐Specific Adaptive Immunity to SARS‐Co V‐2 in Acute COVID‐19 and Associations With Age and Disease Severity,” Cell 183, no. 4 (2020): 996–1012.e 19, 10.1016/j.cell.2020.09.038.33010815 PMC 7494270 · doi ↗ · pubmed ↗
- 5X. Fan , J. W. Song , W. J. Cao , et al., “T‐Cell Epitope Mapping of SARS‐Co V‐2 Reveals Coordinated IFN‐γ Production and Clonal Expansion of T Cells Facilitates Recovery From COVID‐19,” Viruses. 16, no. 7 (2024): 1006, 10.3390/v 16071006.39066169 PMC 11281491 · doi ↗ · pubmed ↗
- 6N. J. Maurice , A. K. Taber , and M. Prlic , “The Ugly Duckling Turned to Swan: A Change in Perception of Bystander‐Activated Memory CD 8 T Cells,” The Journal of Immunology 206, no. 3 (2021): 455–462, 10.4049/jimmunol.2000937.33468558 PMC 7839146 · doi ↗ · pubmed ↗
- 7L. Bergamaschi , F. Mescia , L. Turner , et al., “Longitudinal Analysis Reveals That Delayed Bystander CD 8+ T Cell Activation and Early Immune Pathology Distinguish Severe COVID‐19 From Mild Disease,” Immunity 54, no. 6 (2021): 1257–1275.e 8, 10.1016/j.immuni.2021.05.010.34051148 PMC 8125900 · doi ↗ · pubmed ↗
- 8A. M. Carabelli , T. P. Peacock , L. G. Thorne , et al., “SARS‐Co V‐2 Variant Biology: Immune Escape, Transmission and Fitness,” Nature Reviews Microbiology 21, no. 3 (2023): 162–177, 10.1038/s 41579-022-00841-7.36653446 PMC 9847462 · doi ↗ · pubmed ↗