Cave‐dwelling bats of Carajás National Forest: New cytogenetic data of threatened species
Jéssica Barata da Silva, Thayse Cristine Melo Benathar, Ramerson Lucas Ferreira Azevedo, Leonardo Carreira Trevelin, Cleusa Yoshiko Nagamachi, Guilherme Oliveira, Julio Cesar Pieczarka

TL;DR
This study explores the chromosomal diversity of cave-dwelling bats in Brazil's Carajás National Forest to support conservation efforts and better understand their biodiversity.
Contribution
The study presents the first karyotype descriptions for five bat species in Brazil, including Natalus macrourus and Peropteryx kappleri.
Findings
Cytogenetic analyses revealed chromosomal diversity in eight bat species from Carajás National Forest.
Five species had their karyotypes described for the first time in Brazilian specimens.
The findings support biodiversity conservation and genome sequencing efforts.
Abstract
In Brazil, bat species that use caves as shelters are constantly susceptible to anthropic impacts. Of the 181 species of bats in Brazil, 81 are recorded in caves, 13 of which are considered essentially cave dwellers. The Carajás National Forest Conservation Unit, located in the southeastern region of Pará, serves as a refuge for these species, among others. The present work sought to reveal the chromosomal diversity of bats in six caves of the Carajás National Forest. Cytogenetic analyses were carried out for some species as an additional tool for taxonomic identification. We present here the conventional karyotype of eight species, five being described for the first time in Brazilian specimens. We also described for the first time the karyotypes of Natalus macrourus and Peropteryx kappleri. These findings may be helpful to support conservation guidelines, and for the knowledge of these…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
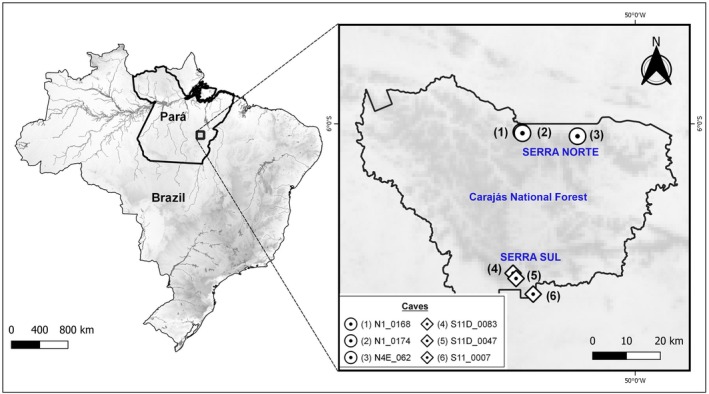 FIGURE 1
FIGURE 1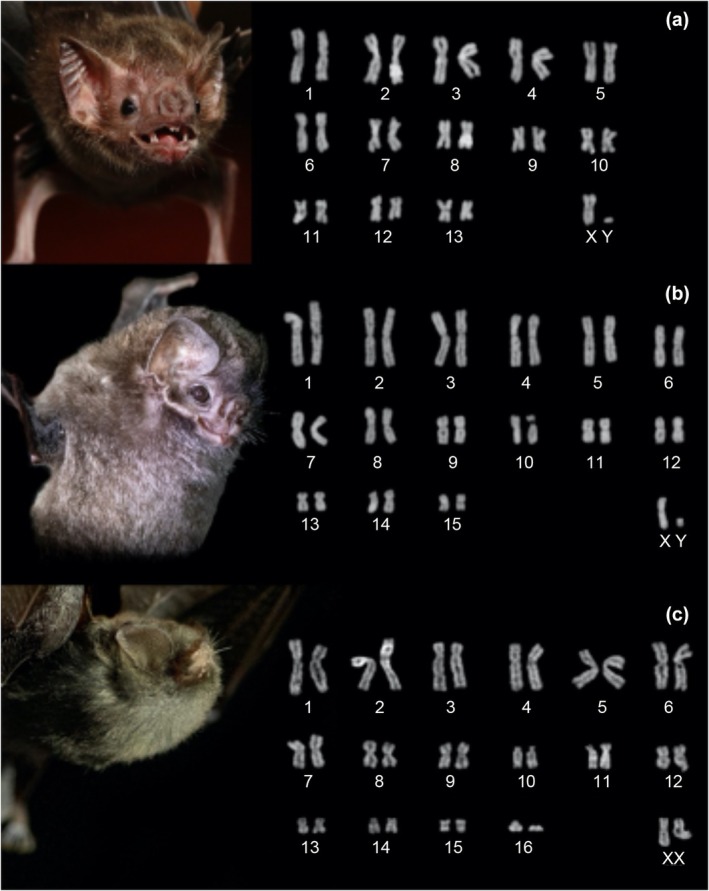 FIGURE 2
FIGURE 2 FIGURE 3
FIGURE 3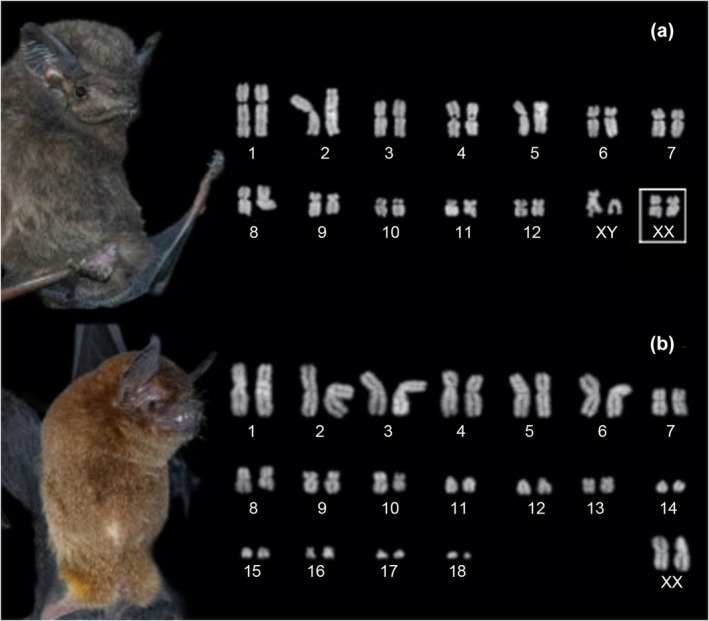 FIGURE 4
FIGURE 4| Family | Species | 2 | FNa | Locality | References | |
|---|---|---|---|---|---|---|
| Phyllostomidae |
| 32 | 60 | Pernambuco, Brazil | Santos et al. ( | |
| Phyllostomidae |
| 28 | 52 | Pernambuco, Brazil | Santos et al. ( | |
|
| 32 | 60 | Nickerie, Suriname | Baker et al. ( | ||
| Furipteridae |
| 34 | 62 | Saramacca, Suriname | Baker et al. ( | |
| Furipteridae |
| 34 | 60 | Carajas, Brazil | Present study | |
| Vespertilionidae |
| 44 | 50 | Huánuco, Peru | Bickham et al. ( | |
| Natalidae |
| 36 | 54 | CNF, Parauapebas, Pará, Brazil | Present study | |
| Emballonuridae |
| 26 | 48 | CNF, Parauapebas, Pará, Brazil | Present study | |
| Mormoopidae |
| 38 | 60 | Veracruz, Mexico | Farias et al. ( | |
| Species | Individuals/sex | Classification | Locality | Cave | Geographic Coordinate (datum WGS‐84) |
|---|---|---|---|---|---|
|
| 1♂ | EC | Serra Sul | S11_0007 (6) | −50.242, −6.456 |
|
| 1♂ | OC | Serra Sul | S11_0007 (6) | −50.242, −6.456 |
|
|
1♀ 1♂ | EC | Serra Norte | N1_0168 (1) | −50.302, −6.022 |
|
| 1♀ | EC | Serra Sul | S11_0007 (6) | −50.242, −6.456 |
|
| 1♂ | OC | Serra Norte | N1_0168 (1) | −50.302, −6.022 |
|
|
1♂ 1♀ | EC | Serra Sul | S11D_0083 (4) | −50.324, −6.397 |
|
|
1♀ 1♂ | OC | Serra Norte |
N4E_062 (3) S11D_0047 (5) | −50.153, −6.033 |
|
| 1♀ | EC | Serra Norte | N1_0174 (2) | −50.299, −6.025 |
- —Instituto Brasileiro de Desenvolvimento e Sustentabilidade
- —Banco Nacional de Desenvolvimento Econômico e Social 10.13039/501100017654
- —Conselho Nacional de Desenvolvimento Científico e Tecnológico 10.13039/501100003593
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBat Biology and Ecology Studies · Evolution and Paleontology Studies · Chromosomal and Genetic Variations
INTRODUCTION
1
Underground ecosystems are widely found in the Neotropical region (Gnaspini & Trajano, 2000; Ladle et al., 2012). Brazil is rich in caves, with an estimated 310,000 such environments in the Brazilian territory, although <5% have been officially recorded (Piló & Auler, 2011). In a given diversity of subterranean environments, many organisms inhabit them (Kunz, 1982).
Caves are important as shelters for bats because they provide a favorable environment for the ecological and physiological maintenance of these mammals (De Sousa Barros et al., 2020). Many bat species are dependent on subterranean ecosystems, as they provide protection against predation, act as sites of reproduction and mating, and facilitate social exchange and the adaptive maintenance of body temperature during torpor or hibernation (De Sousa Barros et al., 2020; Furey & Racey, 2015; Kunz, 1982).
Brazil has comparatively strict legislation protecting underground ecosystems (and bat caves), where environmental licensing processes are required for any economic activity with the potential to impact these ecosystems (Brasil). However, the Brazilian government deals with pressures to make this protective legislation more flexible, which could threaten diverse populations of fauna and flora and the ecosystem services they provide.
The southeastern region of Pará state (Brazilian east Amazonian region) has a wide history of anthropogenic environmental interference, leaving only fragments of forest remnants (Souza‐Filho et al., 2016). The Carajás region, where there is the largest iron ore mine in the world, is home to approximately 1000 caves, being of great speleological importance (Piló et al., 2015). The Carajás National Forest, where many of these caves are located, is a conservation unit that presents a wide range of outstanding landforms and is one of the most important regions in the Brazilian Amazon (Piló et al., 2015). Its management plan establishes areas for scientific research, conservation, and mining, among other uses (Martins et al., 2012). In addition to the regional richness in subterranean environments, the Carajás National Forest is a portion of the Amazon biome composed of a mosaic of dominant vegetation types. Where caves are surrounded by the savanna physiognomy of canga vegetation, such caves are the main stable shelter for bats (Martins et al., 2012).
Caves must be evaluated as nodes in a network of underground environments in karstic landscapes. Therefore, the evaluation and relevance classification of these environments in environmental studies need to consider a broader scale (Delgado‐Jaramillo et al., 2018; Jaffe et al., 2016). Studies aimed at understanding the genetic diversity in bioindicator taxa can help to demonstrate the connectivity between subterranean environments and guide management and conservation decisions.
Of the 181 bat species in Brazil, 81 were recorded in caves and two of the four endangered species (Barros & Bernard, 2023) are dependent on these locations (De Oliveira et al., 2018; Garbino et al., 2020). Conservation strategies for these environments depend on knowledge of species distribution, diversity patterns, levels of endemism, and taxonomy (Moreira et al., 2009). However, the scarcity of knowledge, mainly due to the lack of comprehensive inventories and low taxonomic resolution of the available data, are the main obstacles to proposing conservation measures (Gallão & Bichuette, 2015).
Karyotype can be thought of as a map of the nuclear genome. Far from being only a descriptive branch of knowledge, cytogenetics is valuable as a powerful tool that contributes to bat systematics and taxonomy (Rieseberg, 2001; Sotero‐Caio et al., 2017). As most mammals' species show a species‐specific karyotype, descriptions of karyotypes serve an important role in species characterization and also in defining chromosomal rearrangements, which can provide information about genetic barriers and the processes involved in speciation (Baker, 1967; Faria & Navarro, 2010). In this regard, investigative chromosome studies can provide evidence of karyotypic variation of poorly studied species, such as the threatened cave‐dwelling bat species. Therefore, describing the chromosomal diversity in sensitive environments such as caves and documenting the genetic variation of endangered species are useful tools integrated with other approaches in conservation studies.
Cytogenetic studies in South American bats have a long history, since the 1960s (Baker, 1967), but only in this century the chromosome panting research began (Pieczarka et al., 2005). So, most of the literature on South American bat's karyotypes is on classical cytogenetics analysis. Many species still do not have their karyotypes described. This is especially relevant because cytogenetic information on bat species that inhabit difficult‐to‐access environments, such as caves, is scarce (Table 1).
In this paper we report the results of a cytogenetics study of bats obtained from six caves, aiming to describe their chromosomal diversity in natural caves in the Carajás National Forest. We provided karyotypes of eight important species for the cave ecosystem, two of than classified in the IUCN red list as Least Concern (Furipterus horrens), and Not Assessed (Natalus macrourus) (IUCN). We described for the first time the karyotype of N. macrourus, an important species that is threatened. We compared our data with previously published data from the same or closely related species, to verify possible karyotypic variations.
MATERIALS AND METHODS
2
Study area
2.1
This study was carried out in Carajás National Forest (CNF), which is in the southeastern region of the State of Pará in domains of the Itacaiúnas River basin, affluent of the Tocantins River, whose confluence occurs in the city of Marabá (Figure 1). CNF occupies an area of 411,949 ha, in lands in the municipalities of Parauapebas, Canaã dos Carajás and Água Azul do Norte (Campos & Castilho, 2012). The map was made using QGIS v.3.10.7. The shapefiles containing geographic data (elevation, hydrography, and country limits) were obtained from DIVA‐GIS (Hijmans et al., 2004). We used the limits provided by Braga et al. (2008) and we created the shapefiles on QGIS v.3.10.7.
Location of the study region and zoom of the study area showing the spatial distribution of the sampled caves, whose symbols indicate localities in the “Serra Norte” and “Serra Sul” places. The numbers refer to localities in Table 2.
Collected material, identification, and ethic statements
2.2
We collected bats in two expeditions of three consecutive days in November 2021 and March 2022 that visited three caves in Serra Norte and three in Serra Sul. Captures were conducted using harp traps placed at the caves' entrance and Ecotone Inc. 12 × 3‐m mist nets outside, in the understory of nearby forests. Details of cave names are in Table 2. The traps were open in the late afternoon and were closed after 4 h of sampling.
The captures were authorized by the Brazilian Environment Department under license (IBAMA). JCP has a permanent field permit (number 13248) and a special field permit for collecting samples of cave biodiversity (number 76429) from “Instituto Chico Mendes de Conservação da Biodiversidade”. The Cytogenetics Laboratory from UFPA has a special permit number 19/2003 from the Ministry of Environment for sample transport and 52/2003 for using the samples for research. We followed the guidelines for the use of wild mammals in research (Sikes, 2016), collection, and documentation of specimens (Pitt, 2008). The sample sacrifice was performed by animal welfare guidelines established by Brazilian resolution CFMV n.1000/2012 and in accordance with animal welfare guidelines established by the Animal Ethics Committee (Comitê de Ética Animal) from Universidade Federal do Pará (Permit 68‐2015).
Samples
2.3
Eleven specimens were collected (Table 2) and the vouchers were deposited at the mammal collection of Museu Paraense Emílio Goeldi (MPEG). Each captured bat was identified according to their morphological features and external and craniodental measurements using keys and other available literature describing these species (Gardner, 2008; Simmons, 2005). Bats were classified considering classification as essentially cavernicolous (EC) and occasionally cavernicolous (OC), concerning shelter use strategies of caves (Arita, 1993).
Chromosomal preparations and karyotype analysis
2.4
Chromosomal preparations were obtained through direct bone marrow extraction, according to the procedure described (Baker et al., 2003) and primary culture of fibroblasts (Moratelli et al., 2002) with minor adjustments. A 5% Giemsa in phosphate buffer solution was used to stain the metaphases. For karyotype description and diploid number, 10 metaphases were analyzed per species. The best metaphases were selected for karyotyping. An Olympus BX41 microscope and a CCD 1300QDS digital camera were used to obtain digital images from conventional staining, which were analyzed using the GenA‐SIs software v. 7.2.7.34276.
RESULTS
3
We described diploid chromosome number (2n), and autosomal fundamental number (FNa) of a sample of 11 specimens of eight species. The male specimen of Desmodus rotundus has 2n = 28, FNa52, and all autosomes had meta/submetacentric morphology; the X is a mid‐sized submetacentric and Y is a punctiform chromosome (Figure 2a). The karyotype of Diphylla ecaudata shown 2n = 32, FNa = 60, all autosomes had meta/submetacentric morphology, and chromosome X is submetacentric and Y is a small submetacentric (Figure 2b). The karyotype of a female F. horrens shows 2n = 34, FNa = 60, composed of 14 pairs of meta/submetacentric chromosomes (1–11, 13, 16–17) and two pairs of acrocentric chromosomes (10, 16); since no banding was done, we supposed that the X is a mid‐sized submetacentric (Figure 2c). The male and female specimens of Lonchorhina aurita have 2n = 32, FNa = 60, all autosomes had meta/submetacentric chromosomes; chromosome X is submetacentric and Y is small acrocentric (Figure 3a).
Conventional Giemsa staining of chromosomes of selected species from Carajás National Forest. (a) Desmodus rotundus (2n = 28, FNa = 52); (b) Diphylla ecaudata (2n = 32, FNa = 60); (c) Furipterus horrens (2n = 34, FNa = 60). The Giemsa‐stained chromosomes had their color inverted.
Conventional Giemsa staining of chromosomes of selected species from Carajás National Forest. (a) Lonchorhina aurita (2n = 32, FNa = 60); (b) Myotis albescens (2n = 44, FNa = 50); (c) Natalus macrourus (2n = 36, FNa = 56). Chromosome pair into a square: sex chromosomes from another specimen. The Giemsa‐stained chromosomes had their color inverted.
The karyotype of Myotis albescens shown 2n = 44, FNa = 50, composed of four pairs of meta/submetacentric (1–3, 16) and 17 acrocentric chromosomes (4–15, 17–21); the chromosome X is submetacentric and Y is small acrocentric (Figure 3b). The analyzed N. macrourus specimens presented 2n = 36, FNa = 56, with 11 pairs meta/submetacentric chromosomes (1–9, 13) and 6 pairs acrocentric chromosomes (10–12, 14–17); the chromosome X is submetacentric and Y is small acrocentric (Figure 3c). A male specimen of Peropteryx kappleri has 2n = 26, FNa = 48; all autosomes had meta/submetacentric morphology; the X is a submetacentric chromosome and Y is acrocentric about half the size of the X‐chromosome (Figure 4a). Finally, the karyotype of Pteronotus personatus presented 2n = 38, FNa = 60, composed of 12 meta/submetacentric chromosome pairs (1–10, 13, 16) and 6 acrocentric pairs (11–12, 14–15, 17–18); the chromosome X is mid‐sized metacentric (Figure 4b).
Conventional Giemsa staining of chromosomes of selected species from Carajás Nacional Forest. (a) Peropteryx kappleri (2n = 26, FNa = 48); (b) Pteronotus personatus (2n = 38, FNa = 60). Chromosome pair into a square: sex chromosomes from another specimen. The Giemsa‐stained chromosomes had their color inverted.
DISCUSSION
4
The Carajás National Forest is an excellent target for studying bats because it consists of an area of high biological and landscape diversity, harboring over 1000 caves, one of the largest concentrations of caves in Brazil (Martins et al., 2012; Piló et al., 2015). Of the 23 bat species registered in the CNF caves (Tavares et al., 2012), our study revealed a representativeness of ~35% of this diversity. Furthermore, the reported data on bat occurrence in 269 Brazilian caves classified the species as essentially cavernicolous, opportunistic cavernicolous, and non‐cavernicolous (Guimarães & Ferreira, 2014). In the present study, over half of the karyotyped species investigated are essentially cavernicolous (Table 1).
It is essential to improve the representativeness of cave bats´ cytogenetic data since <5% of the caves in Brazil have been comprehensively explored (Jansen et al., 2012). Furthermore, most studies have focused on areas other than the Brazilian Amazon, leaving the region with <1% of its known cave bat population investigated (Cajaiba et al., 2021; De Oliveira et al., 2018; Guimarães & Ferreira, 2014; Sites Jr et al., 1981). This representatively is further worrying, as Amazonian caves show more variable bat assemblage compositions than caves in other regions of Brazil (De Oliveira et al., 2018).
The karyotypes recovered from our samples were similar to those already published (see Table 1), however, there was no previous knowledge of the karyotypes of Brazilian populations of F. horrens, M. albescens, N. macrourus, P. kappleri and P. personatus. Our sample of F. horrens has a divergent karyotype concerning chromosome structure compared to the karyotyped specimen from Suriname (Baker et al., 1981). In the present study, F. horrens was found to have a karyotype of 2n = 34/FN = 60, while in the previous description the FNa = 62 (Baker et al., 1981). The difference results from the fact that pair 10 is acrocentric in our karyotype while is subtelocentric in the previous study (Baker et al., 1981). However, in both studies, the sexual chromosome pair was not different since in both situations only females were analyzed, so it is possible that the FNa described is different from that suggested. The same happens here since we are not sure that the pair designated as X is the sex chromosome.
Natalus is an infrequently encountered species and is threatened by the practice of extermination of cave bat colonies (e.g., in campaigns against rabies) (Tejedor, 2011; Tejedor et al., 2005). The genus Natalus has a complex taxonomic history and studies based on morphological and molecular characters indicate cryptic diversity in some species (López‐Wilchis et al., 2012). Currently, eight valid species with Neotropical distribution are recognized (Tejedor, 2011). In Brazil, there was some uncertainty in the classification, as N. stramineus has been historically applied to populations of the genus Natalus from virtually the entire Neotropics. Later the populations in the south of the Amazon River were classified as N. espiritosantensis, and the samples of the North, in the shields of the Guyanas as N. tumidirostris (Garbino & Tejedor, 2013). As N. macrourus was considered a senior synonym of N. espiritosantensis (Garbino & Tejedor, 2013), this became the valid name of the species occurring south of the Amazon River.
Baker (1970) described the karyotype of what is supposed to be Natalus stramineus, from populations in Mexico with 2n = 36, FN = 56. Assessing the geographic location of the reported studies, the karyotype of the populations of Mexico is N. mexicanus or N. lanatus (however, some authors disagree that there are two species of Natalus in Mexico, see (López‐Wilchis et al., 2012)), since after review Baker and Jordan (1970) and Tejedor (2011) characterized N. tumidirostris from Trinidad and Tobago with the same chromosome number and morphology. However, N. stramineus is reported only in the northern Lesser Antilles. So, the described karyotype from Trinidad and Tobago may be N. stramineus. By their turn, Volleth et al. (2023) described the karyotypes of N. mexicanus from Cueva de las Vegas, Tenampulco, Mexico having 2n = 36 and FNa = 56, and N. tumidirostris from Candelaria, Venezuela, as having also 2n = 36 and FNa = 56. Those authors state that 2n = 36 and FNa = 56 is the most common form, although some authors describe FNa = 54. To summarize, due to this extensive history of changes in the taxonomy of Natalus, it was assumed that we already knew the karyotype of the species that is distributed in the Brazilian Amazon (Dos Reis et al., 2007), but this was not true until the present work. The karyotype here described is similar to the others previously described, with 2n = 36, and FN = 56. This means that the speciation in this genus is related to geographic isolation. The populations must be large, with a reasonable ability to travel over wide regions, avoiding inbreeding (Dobigny et al., 2015).
Although Natalus species have the same chromosomal number and morphology, knowledge of the cytogenetics of the group can provide useful data in further studies to differentiate the species from the analysis of repetitive portions of the genome using the FISH (Fluorescence in situ hybridization) technique and assist in the complex evolutionary history of this taxon.
In the present study, we described for the first time the karyotype of P. kappleri. This species is not an obligate cave dweller and can also roost in tree hollows and rock crevices (Voss et al., 2016). Previously, cytogenetic data was only available for P. macrotis, with a karyotype composed of 2n = 26 and FN = 48. The same diploid and fundamental numbers were found in our study, without evidence of chromosomal differences when compared to the congener P. macrotis from other Amazonian locations (Baker et al., 1981). The only difference would be that the X chromosome of P. macrotis was described as near acrocentric (Baker et al., 1981) while in our study P. kappleri has a submetacentric X. Regarding the other species, we demonstrate that the specimens sampled in this study show karyotypes characteristic expected for what is already known in the literature for these species, evidencing taxonomical stability.
CONCLUSIONS
5
Cytogenetic data can provide interesting information to help characterize the karyotypes of these species. The karyotypic description has helped in taxonomic studies, including diagnostic characters for delimiting taxa. The study of genetic diversity together with the systematic monitoring of bat populations in the Carajás National Forest region are essential tools in the conservation of these important indicators of biodiversity and the maintenance of cave ecosystems.
AUTHOR CONTRIBUTIONS
Jéssica Barata da Silva: Conceptualization (lead); data curation (lead); formal analysis (equal); writing – original draft (equal). Thayse Cristine Melo Benathar: Formal analysis (supporting); methodology (equal); writing – review and editing (equal). Ramerson Lucas Ferreira Azevedo: Formal analysis (supporting); methodology (equal); writing – review and editing (supporting). Leonardo Carreira Trevelin: Data curation (equal); funding acquisition (supporting); investigation (equal); resources (supporting); writing – review and editing (equal). Cleusa Yoshiko Nagamachi: Data curation (equal); funding acquisition (equal); investigation (equal); resources (equal); writing – review and editing (equal). Guilherme Oliveira: Data curation (equal); funding acquisition (supporting); investigation (equal); resources (supporting); writing – review and editing (equal). Julio Cesar Pieczarka: Project administration (lead); resources (lead); supervision (lead); writing – review and editing (equal).
FUNDING INFORMATION
This research was funded by the Instituto Brasileiro de Desenvolvimento e Sustentabilidade (IABS), grant number 010/2020; by the Conselho Nacional de Desenvolvimento Científico e Tecnológico, grants number 307170/2021‐7 and 307154/2021‐1; and by the Banco Nacional de Desenvolvimento Econômico e Social, grant number 2.318.697.0001.
CONFLICT OF INTEREST STATEMENT
The authors declare no conflict of interest.
INSTITUTIONAL REVIEW BOARD STATEMENT
The animal study protocol was approved by the Ethics Committee (Comitê de Ética Animal) from Universidade Federal do Pará (Permit 68‐2015), which also approved all experimental protocols of this research. The captures were authorized by the Brazilian Environment Department under license (IBAMA 02047.000384/2007‐34). JCP has a permanent field permit (number 13248) and a special field permit for collecting samples of cave biodiversity (number 76429) from “Instituto Chico Mendes de Conservação da Biodiversidade”.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Arita, H. T. (1993). Conservation biology of the cave bats of Mexico. Journal of Mammalogy, 74(3), 693–702.
- 2Baker, R. (1967). Karyotypes of bats of the family Phyllostomidae and their taxonomic implications. The Southwestern Naturalist, 12(4), 407–428.
- 3Baker, R. J. (1970). Karyotypic trends in bats. Biology of Bats, 1, 65–95.
- 4Baker, R. J. , Genoways, H. H. , & Seyfarth, P. A. (1981). Results of the Alcoa Foundation‐Suriname expeditions. VI. Additional chromosomal data for bats (Mammalia: Chiroptera) from Suriname. Mammalogy Papers: University of Nebraska State Museum, 230, 333–344.
- 5Baker, R. J. , Hamilton, M. J. , & Parish, D. A. (2003). Preparations of mammalian karyotypes under field conditions. Occasional Papers, Museum of Texas Tech University, 228, i+1–8.
- 6Baker, R. J. , & Jordan, R. G. (1970). Chromosomal studies of some Neotropical bats of the families Emballonuridae, Noctilionidae, Natalidae and Vespertilionidae. Caryologia, 23(4), 595–604.
- 7Barros, H. M. D. R. , Sotero‐Caio, C. G. , Santos, N. , & de Souza, M. J. (2009). Comparative cytogenetic analysis between Lonchorhina aurita and Trachops cirrhosus (Chiroptera, Phyllostomidae). Genetics and Molecular Biology, 32(4), 748–752.21637449 10.1590/S 1415-47572009005000095 PMC 3036889 · doi ↗ · pubmed ↗
- 8Barros, J. S. , & Bernard, E. (2023). Species richness, occurrence and rarity of bats in Brazilian caves. Austral Ecology, 48, 2144–2170. 10.1111/aec.13453 · doi ↗