Gama rays mediated improvement of catalytic efficiency and thermostability of glucoamylase by replacing active site leucine to isoleucene from super koji (Aspergillus oryzae)
Anam Saqib, Saif -ur-Rehman, Hazrat Ali, Noor Hassan, Asad Ali, Muhammad Hamid Rashid, Habibullah Nadeem, Habibullah Nadeem, Habibullah Nadeem

TL;DR
This study uses gamma rays to improve the efficiency and heat stability of a key food industry enzyme by altering its active site.
Contribution
A novel point mutation (Leu203 → Ile) in glucoamylase from Aspergillus oryzae enhances catalytic efficiency and thermostability via gamma ray-induced mutation.
Findings
Mutant glucoamylase M-60(5) showed 1.62-fold higher kcat and 4.75-fold higher kcat/Km compared to the parent enzyme.
The mutant enzyme had a 1.92-fold longer half-life at 55°C and improved thermodynamic parameters for starch hydrolysis.
The point mutation altered the enzyme's active site conformation, stabilizing the transition state and enhancing performance.
Abstract
Glucoamylase is considered as an essential enzyme in food industry. However, lowere catalytic efficiency and weak thermostability confine its application in food industry. Therefore, the current study was aimed to improve catalytic efficiency and thermostability of glucoamylase by replacing active site leucine to isoleucene from Super Koji (Aspergillus oryzae) using gama rays mediated point mutation. High catalytic efficiency and thermostability of glucoamylase from mutant Aspergillus oryzae M-60(5) (screened from 51 mutants) was achieved due to a point mutation, i.e., Leu203 → lle in active site. The SDS-PAGE molecular mass of parent and mutant glucoamylase was 63.1 kDa, while mutant glucoamylase showed; productivity = 9.7 U ml‒1, kinetic constants kcat = 118 (1.62 fold), (kcat/Km) = 1899 (4.75 fold) and half-life at 55 °C for 45 min (1.92 fold). Thermodynamics parameters for starch…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
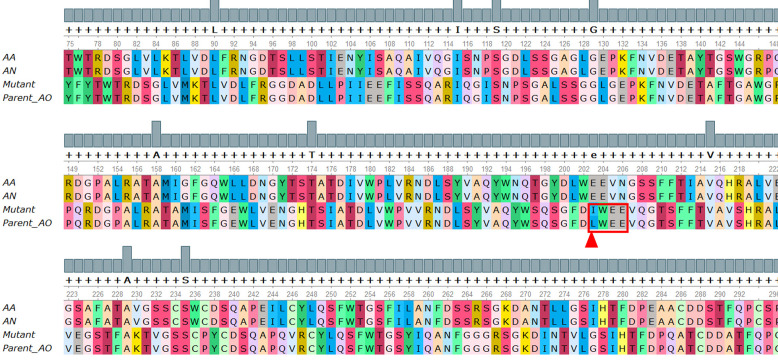 Fig 1
Fig 1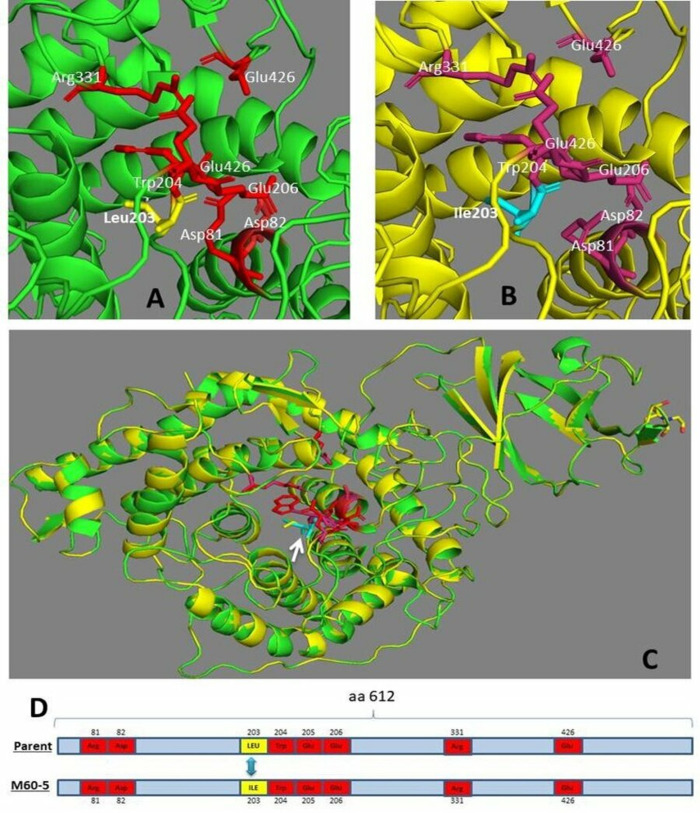 Fig 2
Fig 2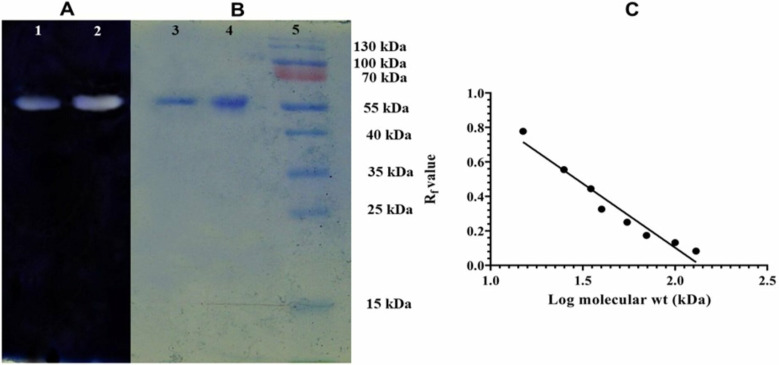 Fig 3
Fig 3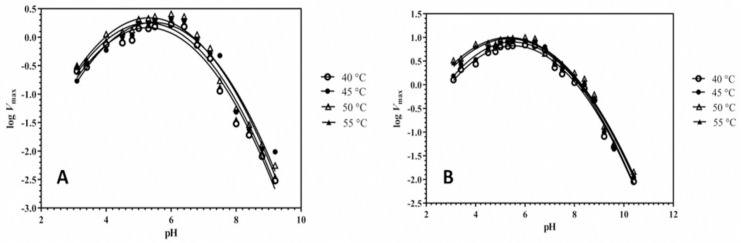 Fig 4
Fig 4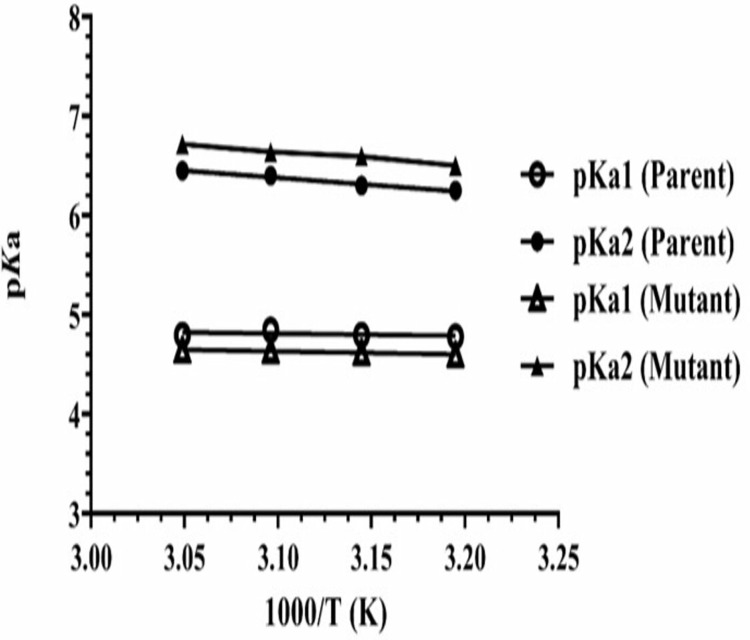 Fig 5
Fig 5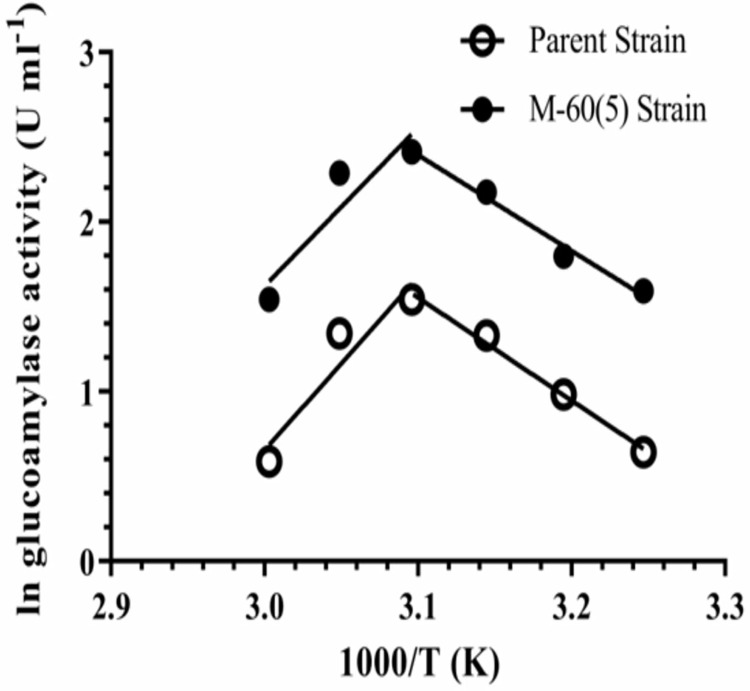 Fig 6
Fig 6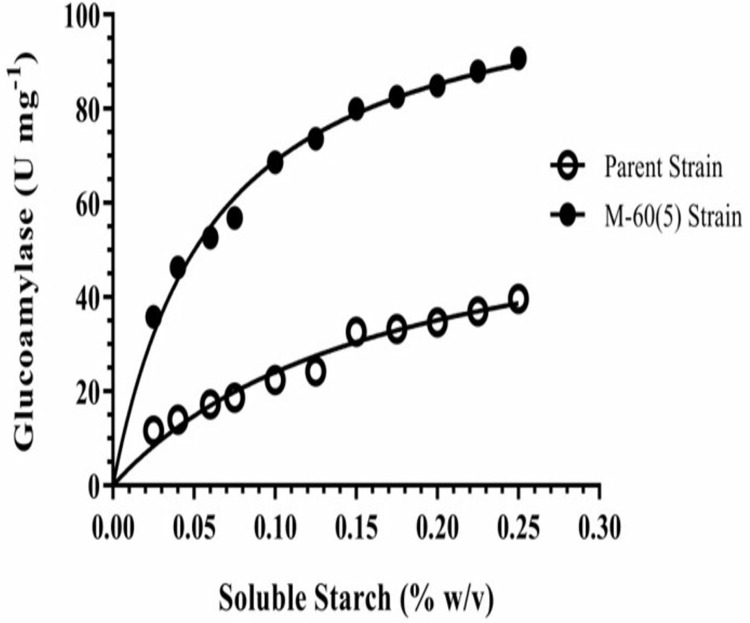 Fig 7
Fig 7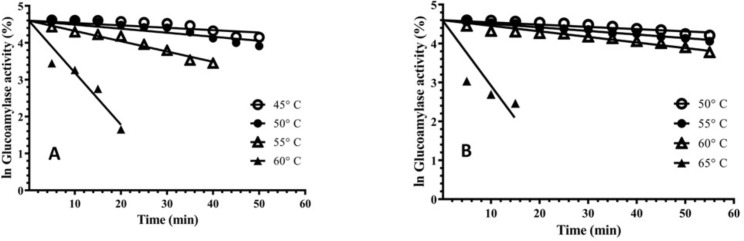 Fig 8
Fig 8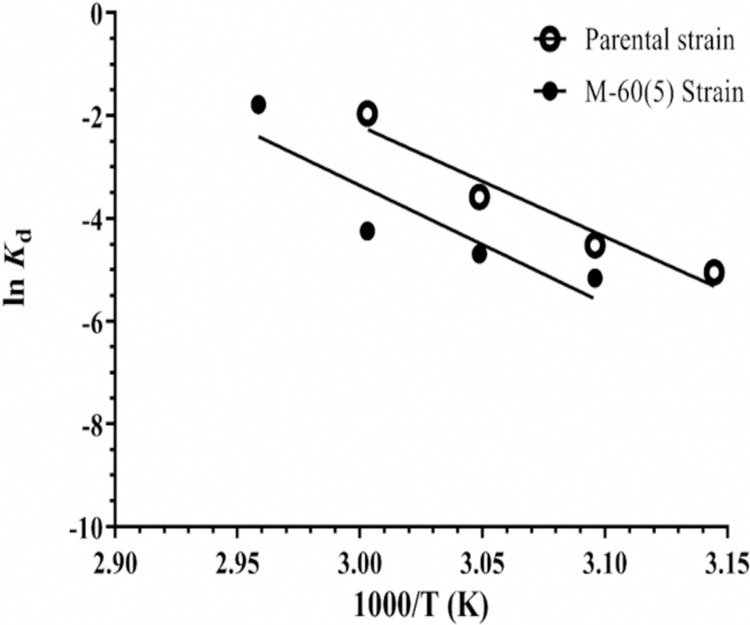 Fig 9
Fig 9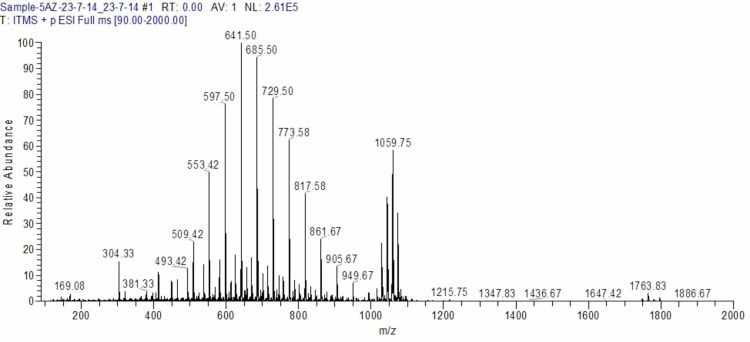 Fig 10
Fig 10Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEnzyme Production and Characterization · Protein Hydrolysis and Bioactive Peptides · Enzyme Catalysis and Immobilization
Introduction
Glucoamylase (GA) is the most widely used enzyme [1] and has distinctive properties of hydrolyzing starch into subunits of oligosaccharide and commonly used for production of high corn, glucose and fructose syrups as well as for alcohol production [2–4]. The GA produces glucose directly by scarification of α-1,3, α-1,4 and α-1,6 glycosidic bonds present at non-reducing end of starch molecule [5,6]. Aspergillus oryzae is the main focus of food industries for hydrolytic enzymes production such as α-amylase, GA and other food grade enzymes. Different fungal species, e.g., Aspergillus niger, Rhizopus niveus and Rhizopus delemar, are used to produce GA commercially [7]. However, their lower catalytic activity leads to consume high energy and restrictions in starch processing and fermentation [8]. Hence, it is a dire need to develop novel fungal GA suitable for evolving industrial applications.
The A. oryzae is considered as Generally Recognized As Safe (GRAS), therefore their enzymes are safe for food and feed industry [8,9]. In recent era, strain improvement by site directed and random mutagenesis of fungi is being used widely for industrial applications. Strain improvement usually targets for elevated yields of the enzymes and biomass production, broad spectrum uses of industrially suitable substrates and efficient physiological properties [10]. Implementation of various molecular methods like recombinant DNA technology and γ-rays dependent mutagenesis is being carried out to evolve strains in order to meet increase industrial demands for food grade enzymes [11].
The complete open reading frame of A. oryzae GA contained 612 amino acids residues and GA protein is composed of carbohydrate binding module (CBM) of 106 amino acid residues from 506–612 [12]. The A. oryzae GA showed 67% and 30% homology with A. niger and R oryzae, respectively [13,14]. Five highly conserved regions were reported in GA enzyme of A oryzae and A. niger with 78–95% similarity, whereas, the active site of A. oryzae was homologous to A. niger [15].
Random mutagenesis induced by γ-rays may contribute in enhancement of industrial enzymes production; hence γ-ray mediated mutagenesis of A. niger made the mutant GA highly efficient in substrate hydrolysis (Aleem et al. 2018). Kinetics and thermodynamics of GA suggested that the mutated enzyme have potential for commercial scale glucose production in starch processing as well as in the food industry [16,17]. Previously, we developed novel Aspergillus oryzae mutants by γ-rays’ mutagenesis for the enhanced production of thermostable α-amylases, which were also highly specific in the α-amylase production [18]. The increase in production, catalytic efficiency and thermostability of α-amylases proved that the γ-rays might have altered the α-amylases conformation [18]. In the current study, we further have screened the previously generated koji mutants for the thermostable GA hyper producer strains. Consequently, mutant M-60(5) was identified as the potent hyper producer of thermostable GA.
Novelty of current report is as it for the first time explains about effect of point mutation, i.e., replacement of catalytic center leucine to iso-leucine on the active site conformation; kinetics and thermodynamics of stability-function of the GA. Moreover, mutation’s effect on the GA was further attributed through evaluation of active site microenvironment by determining the heat of ionization of active site residues.
Methods
Culture maintenance and mutant preparation
The super Koji (A. oryzae cmc1) and its mutant derivative M-60(5) strain (hyper producer of thermostable GA) were obtained from Industrial Enzymes & Biofuels Group, Industrial Biotechnology Division, National Institute for Biotechnology and Genetic Engineering (NIBGE), Faisalabad. The Koji M-60(5) mutant was screened from the mutants of super Koji, which were previously developed by γ-rays treatment [18]. Briefly, the mutants were generated by preparing fresh inoculum of A. oryzae in six 15 ml falcon tubes and irradiated with Caesium-137 (Cs-137) γ-ray and γ-rays source fitted in gamma cell radiation chamber (Mark-IV Irradiator/Gamma Cell-220). Main stock of mutants (51 variants) was exposed to various level of γ-ray exposure, e.g., 60, 80, 100, 120 & 140 kRad (0.6, 0.8, 1.0, 1.2 & 1.4 kGray). The potato dextrose agar was used to maintain culture at 30 °C as described by Aleem et al. [18].
Production and purification of GA
The GA from parent and mutant M-60(5) strains was produced in 10L bioreactor. Briefly, six liter of liquid fungal growth medium (LFGM) containing Rafhan starch (2% w/v) with pH 6.5 was prepared. The media was autoclaved for 40 min at 121 °C, 30 psi. Afterwards, the temperature was lowered down to 30 °C and stirred continuously at 150 rpm. The inoculum was grown in 500 ml flask as described [18]. The cell density of inoculum in triplicate was measured and transferred to bioreactor aseptically (0.3% w/v pack cells). The fermenter was run for 72 hrs and aliquots were taken from the bioreactor after every 6 hrs and centrifuged (25,900 × g) for 10 mints to clear the sample. Afterwards, the total cell mass and GA activity was determined in the samples. The muslin cloth (maximum pore size 2 mm) was used to filter growth medium and the filtrate was centrifuged (25,900 × g) for 20 min at 4 °C. Finally, the supernatant was lyophilized using Benchtop Freeze Dryers, (Labconco™ FreeZone™, 115V US Models) to concentrate the enzyme as described [19].
The GA was purified by fractional precipitation. Briefly, the solid ammonium sulphate was added steadily to 1.0 ml of concentrated crude GA to achieve saturation ranging from 10–90% at 4 °C. The GA preparations containing ammonium sulfate were then placed overnight at 4 °C and centrifuged (25,900 × g) for 20 min. The pallet was discarded and GA activity was checked in the supernatants. Salt concentrations relating to onset and the complete precipitation of GA were selected. Hence, for large scale purification of GA, solid ammonium sulfate was added slowly to achieve 40% saturation and placed overnight at 4 °C and centrifuged as described above. After that pellet was discarded and 75% saturation was achieved by adding more salt. Again, supernatant was discarded and pellet containing GA was kept for next step. Distilled water was added to pellet and dialyzed for removing salt at 4 °C for 24 hrs.
Sequencing and in-silico point mutation identification in GA gene
The freshly grown mycelia of parent and mutant strain M-60(5) were used for genomic DNA extraction, which was extracted as reported [20]. The PCR fragments were obtained by using 5′ATGCGGAACAACCTTCTTTTTTCC3′ and 5′CTACCACGACCCAACAGTTGGG3′ as primers, according to thermo profile of 94 °C for 5 min; 35 cycles of 94 °C for 60 s; 61 °C for 1 min, 30 s; 72 °C for 2 min and 72 °C for 10 min. The amplified DNA fragment was applied on poly acrylamide gel for the gene identification and PCR product was purified by using PCR purification kit (GeneJET PCR Purification Kit, Catalog number: K0701, Fermentas, Thermoscitific®) and sequenced by Macrogen, Republic of Korea.
The nucleotide sequence alignment of mutant M-60(5) was performed by Clone Manager 10 tool [21] against parent koji strain. The local and global protein alignment tools at European Molecular Biology Open Software Suite (EMBOSS) was used for converting cDNA to amino acid [22]. The InterPro at European Molecular Biology Laboratory- European Bioinformatics Institute (EMBL-EBI) and National Center for Biotechnology Information (NCBI) conserved domain database were utilized for deducing mutant domain information [23]. After domain localization, 3D structure was constructed by Swiss modeling [24] and superimposing of parent and mutant M-60(5) structures was done by PyMol [25] to observe mutational change in active site of the GA encoding protein.
Glucoamylase (GA) assay
The GA activity was determined using soluble starch (1% w/v) as substrate. The assay reaction consisting 100 µl enzyme extract, 1.0 ml of soluble starch (1% w/v) in sodium acetate buffer pH 5.0 and incubated at 50 °C for 40 min. The quenched reaction mixture (QRM) in 1.0 ml of Glucose oxidase/Peroxidase kit (Fluitest® GLU, Biocon, Bangalore, India) was used to determine released glucose [20,26]. One unit of GA activity was defined as the amount of GA required to release one μmol of glucose min^‒1^ from soluble starch under defined assay conditions of temperature and pH. The GA activity units were calculated by using the formula as described by Aleem et al. [18].
Protein assay
The extracellular proteins released were estimated by using Bradford assay and bovine serum albumin was used as a standard [27].
Molecular mass determination
The purity of purified GA and their subunit molecular mass was determined by 10% sodium dodecyl sulphate denaturing-renaturing polyacrylamide gel electrophoresis (SDS-DR-PAGE). Protein markers from Thermoscientific^®^ ranging in size between 10–180 kDa were used and run as standard. Coomassie brilliant blue R-250 solution (0.1%) was used to stain gel containing enzyme and molecular markers. Apparently pure GAs (0.5 μL of 0.6 mg mL ‒1) was spotted separately on SDS-PAGE gel and incubated at 50 °C for 90. Later, replica copy of the SDS-PAGE gel was cut and stained with Iodine solution [28]. A transparent band appeared after 20 min of staining with blue back ground.
Optimum pH, pKa & heat of ionization (ΔHI) of active site residues
The purified GA extracted from mutant M-60(5) was checked against various pH using different buffer systems, e.g., Citrate buffer: pH 3.0–6.2, Sorenson’s buffer: pH 5.8–8.0 & Glycine-NaOH buffer: pH 8.6–10.6. Variable temperatures (40–55 °C) were used to find the optimum pH of GA. In addition, Dixon and Webb [29] protocol was used to determine the dissociation constants (pKa_1_ and pKa_2_) of active site ionizable residues of GA forming ES-complex. Finally, the ΔHI was calculated by using the equation below:
Temperature optimum, temperature quotient (Q10) & activation energy
GA activity was checked against different temperature ranging from 45–60 °C. The assay was conducted at pH 5 (50 mM Na-acetate buffer) for 45 min. The activation energy (Ea) was calculated using Arrhenius plot, while temperature quotient (Q10) was calculated as described [29].
Kinetics & thermodynamics of starch hydrolysis
Different defined amount of GA with various conc. of substrate (soluble starch) ranging from 0.025% to 0.25% (w/v) were used to determine the Michaelis Menten kinetic constants (Km, Vmax, kcat and kcat/Km) for soluble starch hydrolysis by Koji’s GAs at 50 °C, pH 5. The graphpad prism version 7.04 was used to fit date to non-linear regression. Furthermore, Eyring’s absolute rate equation was used to calculate the thermodynamic parameters for substrate hydrolysis usingformula described by Eyring and Stearn [30].
Thermodynamics of irreversible thermal stability
To determine irreversible thermal inactivation of Koji’s GAs 15 ml of the parental and mutated GA solutions were taken in falcon tubes and incubated at various temperatures like: 45, 50, 55 and 60 °C in a water bath. The GA aliquots were taken from each sample after regular time intervals, i.e., 0, 5, 10, 15, 20, 25, 30, 35, 40, 45 & 50 min, which were then placed in ice cold water for 30 min for the refolding. Afterwards, the withdrawn GA aliquots were assayed for % residual activity of the GA. The data was fitted to first order plot and rate constants for irreversible thermal inactivation (Kd) of GAs were calculated.
The activation energy for irreversible thermal inactivation [Ea(d)] of GA was determined by using the Arrhenius plot. Rearranged Erying’s absolute rate equation was applied to determine the thermodynamics of irreversible inactivation of GAs as mentioned above with the difference that kcat was replaced with Kd (denaturation constant). Moreover, in entalphy change (Δ*H**) determination the Ea was replaced with Ea(d).
Toxin analysis by Liquid chromatography–mass spectrometry
The sample was prperaed by taking equal volume of mutant GAs and 100% chloroform and mixed in a separating funnel. The chloroform was evaporated in rotary evaporator and 2 ml methanol was used to disolve the residues. After that, the mixture was filtered through 0.45 µm nylon membrane and subjcted to analysis by Quadrupole Linear Ion Trap Mass Spectrometer Finnegan LTQ XL hyphenated with Surveyor Plus LC system (LC-MS) (Thermo Fisher Scientific, USA) using the protocal described by [18] and [31] with slight changes. The parameter were set as capillary temperature 335 °C, voltage 45 V, spray voltage 5 kV, sheath gas flow rate 70 and auxiliary gas flow rate 20 arbitrary units. Different aflatoxins ml^‒1^ (B1, B2, G1 and G2) were used as standards.
Results
Production and purification of GA
In the current study, parent super Koji (A. oryzae) and its mutant derivative M-60(5) strain were screened based on the hyper production of thermostable GA were grown under submerged conditions on raw maize starch in 10L fermenter. The GA produced by mutant M-60(5) was 9.7 U ml^‒1^ which was 2.6 fold higher than the parent (3.6 U ml^‒1^). Whereas, the specific activity of the mutant was 1.83 fold increased (54.9 U mg^‒1^) as compare to parent. The GA produced by super koji parent and mutant strains was subjected to single step purification. Fractional precipitation of GAs by ammonium sulfate gave single band on 10% SDS-PAGE, which confirmed the GAs had purity apparently at homogeneity level. The precipitation trend of the mutant enzyme was slightly faster than the control depicting that the γ-rays might has changed the surface of the glubular protein. After purification, the specific activity of the purified GAs from parent and mutant M-60(5) Koji strains was increased to 51.8 and 96.2 U mg^‒1^, respectively.
Sequencing and point mutation identification in GA gene of M-60(5)
The sequenced data analysis revealed the size of genomic DNA and cDNA of GA genes as 2,039 bp and 2,241 bp, respectively, with the difference of four introns varying in sizes of 49 bp, 52 bp, 45 bp and 56 bp. The open reading frame (ORF) of mutant and parent GA genes consisted of 2,039 bp encoding 612 amino acid residues. The multiple sequence alignment of amino acid sequences from parent A. oryzae and mutant M-60(5) along with A. niger and A. Awamori as a reference has shown that γ-rays treatment resulted into a point mutation at nucleotide position 703, where the cytosine was replaced by adenine that resulted in a change of amino acid in catalytic site, i.e., Leu at position 203 into Isoleucine (Fig 1).
Multiple sequence alignment of A.Awamori, A. nigar, A. oryzae parent and Mutant M-60(5) amino acid sequences. The point mutation is highlighted in a red colored rectangle pointed by red triangle at the bottom.
Furthermore, the 3D structure of parent and mutant M-60(5) GA enzyme showed that replacement of single amino acid has made a slight change in the microenvironment of the active site (Fig 2).
Structural comparison between GAs from A. oryzae parent and mutant M-60(5).(A) Active site of parent GA with Leucine at 203 position, (B) Active site of Mutant M-60(5) GA with Ile at 203 position and (C) superimposition of the predicted GA model of Parent (green) and the structure of GA from Mutant M-60(5) (yellow). Residues involved in substrate recognition and in catalytic site are shown in magenta and red, while the replacement of Leucine in parent into Isoleucine in mutant are shown in cyan and yellow and indicated with the arrow (white). (D) The schematic diagram to point out the mutation with yellow color.
Molecular mass determination
Similar subunit molecular mass (63.1 kDa) of GAs from Parent and M-60(5) strain was found on 10% SDS-DR-PAGE, which was further confirmed by activity staining of GA. Moreover, the accuracy was maintained by a standard curve between molecular mass and R_f_ values of the protein ladder (Fig 3, S1 Fig ). The calculated subunit mass was nearly same as 65 kDa predicted from the deduced amino acid sequences of GA mutant M-60(5) and parent strain.
Determination of subunit molecular mass of glucoamylase (GA) produced by A. oryzae parent and mutant M-60(5).A): Zymographic analysis: Lane-1 Parent GA, Lane-2 M-60(5) GA. B): Lane-3 purified parent GA, Lane-4 M60(5) GA, Lane-5 protein markers [130, 100, 70(Red band), 55, 40, 35, 25 kDa] stained by Coomassie brilliant blue R-250 stain. C): Standard curve for protein markers ladder to determine the subunit molecular mass of GAs produced by A. oryzae strains.
Effect of pH & enthalpy (ΔHi) of active site residues ionization
The similar optimum pH (6.0) for GA activity was observed for both A. oryzae parent and mutant M-60(5), while optimum pH range with about 70% activity for mutant was 3.4–6.5, whereas for the parental enzyme it was 5.0–6.5. The ionizable groups of active side residues involved in the maximum velocity (Vmax) were determined by applying Dixon plot (Fig 4). The pKa values define as ionization constant which describes the effect of pH on chemical shift or of an enzyme’s activity in a reaction. The present study revealed the pKa1 of proton-donating ionizable group of parent and mutant were 4.5 and 4.55, respectively, while pKa_2_ of proton-receiving group for the parent was 6.5 and for mutant M-60(5) was 6.7. Carboxylic acid and imidazole were acting as ionizable groups for acidic and basic limbs, respectively in active site of both the parent and mutated Koji GA at 50 °C.
Dixon plots for the determination of pKa values of active site residues of GAs from (A) A. oryzae parent, (B) A. oryzae M-60(5) controlling maximum velocity for soluble starch hydrolysis at various temperatures.
Furthermore, effect of γ-rays on the conformation of GA active site was assessed indirectly by evaluating the heat of ionization (ΔHi) of ionizable groups of active site residues (Fig 5). The ΔHi of proton donating residue for parent and mutant GA was 1054 cal mol^‒1^ and 1461 cal mol^‒1^, while for the proton receiving residue was 6576 cal mol^‒1^ and 6581 cal mol^‒1^, respectively. Therefore, we considered the conformational change in active site of mutant’s GA might be due to the π MO ionization of aromatic histidine and NH_2_ nitrogen lone pair ionization of Glu/Asp residues.
Determining heat of ionization (ΔHi) of the active site residues of GAs from A. oryzae parent and mutant M-60(5) strains by Dixon plot.
Temperature optimum, activation energy & temperature quotient
The GA from A. oryzae parent and mutant M-60(5) exhibited similar optimum temp, i.e., 50 °C with optimum temp range of 45–55 °C (~80% of optimal activity). The working ability of an enzyme at elevated temperature is referred as its thermophilicity, while the resistance against unfolding at higher temperature is termed as thermostability of an enzyme. The activation energy (Ea) determined by Arrhenius plot for the soluble starch hydrolysis for parent GA was 50.4 kJ mol^‒1^, while for the mutant M-60(5) it was 46.9 kJ mol^‒1^ (Fig 6). Lower energy requirement by the mutant GA indicated that it was better as compared to the parent GA. Hence, we considered the mutation due to γ-rays treatment might have changed the microenvironment of the active site, resulting into improved conformation, which made the mutant M-60(5) GA more efficient in making the transition-state complex (ES*).
Arrhenius plot to determine the activation energy for soluble starch hydrolysis by GA from A. oryzae parent and mutant M-60(5) strains.
Kinetics & thermodynamics of substrate hydrolysis
Effect of mutation on the Michaelis Menten kinetics constants (Km, Vmax, kcat & kcat/Km) of soluble starch hydrolysis by the Koji GAs were analyzed by fitting the data to non-linear regression using the Graphpad Prism software (Fig 7). The kinetics of soluble starch hydrolysis by mutant GA was drastically improved due to replacement of Leu203 with Ile203. The Km, which is the measure of binding affinity of substrate with the enzyme, of mutant M-60(5) GA for soluble starch hydrolysis at 50 °C, pH 5.0 was lower (0.06 mg ml^‒1^) than the parental GA having 0.17 mg ml^‒1^. Hence, the γ-rays induced mutation in Koji mutant M-60(5) has changed the conformation of active site and made it 2.8 folds more efficient to make the ES complex (Table 1).
Table 1: Kinetics and thermodynamics of soluble starch hydrolysis by GA produce of A. oryzae parent and mutant M-60(5) at 50 °C and pH 5.0.
Michaelis-Menten kinetic (Vmax, Km) constants for the soluble starch hydrolysis by GA from A. oryzae parent and mutant M-60(5) at 50 °C, pH 5.0.
The Kcat (turn over) catalytic events performed by active site of mutated GA were 1.7 fold higher than the parent GA, i.e., 118 s^‒1^ (Table 1). Furthermore, substrate specificity constant (kcat/Km) of mutated GA was 1899 and was 4.7 fold higher than control. Since it gives information about substrate specificity when its concentration is extremely low (<<Km), the results pointed towards the alternation in active site conformation. We concluded that mutation in active site residues made the mutant GA highly specific for starch hydrolysis. The catalytic efficiency of GA produce of A. oryzae mutant M-60(5) was extremely higher by having Km of GA (Km = 0.062 mg ml^‒1^).
The Gibbs free energy for soluble starch hydrolysis (ΔG**) of M-60(5) GA was decreased as compared to parental GA, hence, confirmed that it required lower amount of functional energy to convert the transition complex (ES) into products. Moreover, reduction in ΔH * of mutated GA with 3.492 kJ mol^‒1^ also confirmed that it required lower energy to make the activated transition complex (ES*). On the other hand, ΔS * for starch hydrolysis of mutant GA was decreased, which confirmed that the activation in substrate hydrolysis was not entropically driven (Table 1).
Kinetics and thermodynamics of irreversible thermostability
The thermostability of mutant GA was increased about two fold at 55 °C than the parental enzyme, while at 45 °C & 50 °C a modest increase in the stability was observed (Fig 8). The increase in thermal stability of M-60(5) GA was due to higher Gibbs free energy, which was increased with a difference of 3.43 kJ mol^‒1^. The higher free energy helped the mutated GA to resist against thermal unfolding of its transition state (U*) into an inactive enzyme.
This figure presents irreversible thermal inactivation of GA from (A): A. oryzaeparent, (B): A. oryzae M-60(5) at variable temperatures using Pseudo first order plots.
Moreover, activation energy Ea(d) to make the transition state ‘U*’ of mutant GA was also higher than the parental one, pointing towards higher stability of the mutated GA (Fig 9). The change in entropy (ΔS**) of mutated GA at temperatures ranging from 45°C–60°C was higher than the parental GA, which indicated towards the increase in disorder of its active site conformation (Table 2). It was concluded that the increase in thermostability of mutant GA was due to higher ΔG* * and was not entropically driven.
Table 2: Kinetics and thermodynamics of irreversible thermal stability of GA from parent and mutant M-60(5) A. oryzae.
Determining activation energy for irreversible thermal denaturation of GA from A. oryzae parent and mutant M-60(5) strains by Arrhenius plot.
Toxin analysis on LC-MS
The Mutant M-60(5) GA was analyzed for toxin analysis on LC-MS against standard and already reports control values. It was confirmed that the M-60(5) GA did not produce any aflatoxins (Fig 10).
Aflatoxin analysis of Mutant M- 60(5) through LC-MS.
Discussion
A. oryzae is used in food processing industries for centuries. Therefore, numerous attempts have been made to improve the A. oryzae strains for making process more feasible. Various methods are used for strain improvement, e.g., random mutagenesis [18]. Hence, in the current study, point mutation, i.e., replacement of catalytic center leucine to iso-leucine on the active site conformation was introduced in A. oryzae for improving catalytic efficiency and thermostability of Glucoamylase (GA) by γ-rays induced random mutagenesis.
In the current study, the GA was produced 2.6 fold higher (9.7 U ml^‒1^) by mutant M-60(5) as compared to parent (3.6 U ml^‒1^). Whereas, the specific activity of the mutant was 1.83 fold increased (54.9 U mg^‒1^) as compare to parent. The increase in GA enzyme activity in A. oryzae by γ-rays mutagenesis is confirmed in an already reported study [32]. The mutant M-60(5) strain produces much higher GAs than already reported strains, i.e., the maximum units of GA under optimum conditions are reported as 3.5 U ml^‒1^ from A. wentii [33], 8.23 U mL^‒1^ from A. Oryzae [34] and 5.9 U mg^‒1^ from A. Awamori [35]. The crude GAs from A.oryzae parent and Mutant M-60(5) were purified by ammonium sulphate precipitation as described [26,36,]. After purification, the specific activity of the purified GAs from parent and mutant M-60(5) Koji strains was increased to 51.8 and 96.2 U mg^‒1^, respectively which are much higher than the specific activity of purified GA from A. niger, i.e., 16.2 U mg^‒1^ by three steps purification [17], however the specific activity of purified GA from A. fumigatus was reported as 94 U mg^‒1^ [37].
In addition, the multiple sequence alignment of amino acid sequences from parent A. oryzae and mutant M-60(5) along with A. niger and A. Awamori as a reference has shown that γ-rays treatment resulted into a point mutation at nucleotide position 703, where the cytosine was replaced by adenine that resulted in a change of amino acid in catalytic site, i.e., Leu at position 203 into Isoleucine. In previous studies, a single replacement of leucine to isoleucine in active site has drastically effect the enzyme activity of Taq polymerase by converting it into highly cold sensitive enzyme [38], whereas as a result of mutagenesis, a single conserved amino acid replacement, i.e., leucine to isoleucine has affected the ‘g’ protein signals mechanism [39]. Hence, based on the above report we concluded that the replacement of leucine203 into isoleucine in M-60(5) Koji GA might have drastically changed the conformation of its active site, resulting into an efficient and stable mutant GA enzyme. We believe that improvement in catalytic efficiency of GAs in mutant was linked with the change in a Leu203 to Ile203 at active site of GA Mutant M-60(5).
In the present study, the molecular mass of GAs was determined and fund subunit molecular mass (63.1 kDa) of GAs from Parent and M-60(5). Moreover, the accuracy was maintained by a standard curve between molecular mass and R_f_ values of the protein ladder. Mutant M-60(5) showed almost similar molecular mass as of GA from Aspergillus oryzae from Luzhou-flavour Daqu [40] and A. Fumigatus [37]. Already reported studies supports our finding of similar subunit molecular mass of parent and mutant strains from Aspergillus oryzae [41] and from A. flavus [42]. These findings are in accordance with other published results, where the GA from A. flavonus shown protein size of 78 kDa which varied from the calculated size of 55.1 kDa [34].
Furthermore, optimum pH range with showing 70% activity for mutant was between 3.4–6.5. The more acidic pH range of mutant GA indicated that the γ-rays mediated point mutation has slightly changed the microenvironment of the active site of mutant GA. Previous studies showed the fungal GA remains more active at acidic pH [43,44]. The maximum catalytic activity of this enzyme has also been reported at pH 5 to 6, whereas in A. Niger the optimum reported as pH 4.8 [45]. The present study revealed the pKa1 of proton-donating ionizable group of parent and mutant were 4.5 and 4.55, respectively, while pKa_2_ of proton-receiving group for the parent was 6.5 and for mutant M-60(5) was 6.7. The difference in pKa values of acidic and basic limbs of the active site residues of mutated GA confirmed about the changed configuration of the active site due to random mutagenesis resulting into more efficient catalytic activity of mutant M-60(5). Literature confirms that the change in microenvironment around the catalytic domain resulting into improvement in catalytic efficiency and protein stability at low pH [46]. The reported ionizable groups values of amino acids located in proteins indicated the presence of glutamic/aspartic acid as the proton donating residue, while histidine as the proton receiver [29].
The ΔHi of proton donating residue for parent and mutant GA was 1054 cal mol^‒1^ and 1461 cal mol^‒1^, while for the proton receiving residue was 6576 cal mol^‒1^ and 6581 cal mol^‒1^, respectively. Similar findings regarding proton donating residues were presented by [15], where three Glutamic acid (Glu) and one Aspartic acid (Asp) residues participated as the electron donor in GA active site. Effect of temp on pKa_1_ & pKa_2_ and enthalpy or heat of ionization (ΔHi) of active site residues ionizable groups was determined as described by Dixon and Web [29] (Fig 5). The amino acid composition of mutant’s active site was not changed, however, the increase in ΔHi of proton donating (407 cal mol^‒1^) and receiving (5 cal mol^‒1^) residues evidenced about the change in conformation of active site. Dehareng and Dive [47] evaluated the ionization energies (IE) as a function for the conformation of α-L-amino acids and optimized three to five conformations for the arginine, lysine, isoleucine, tyrosine and tryptophan in their study.
Similarly, the GA from A. oryzae parent and mutant M-60(5) exhibited similar optimum temp, i.e., 50 °C with optimum temp range of 45–55 °C (~80% of optimal activity), which is considered as much higher optimal activity at given range than already reported A. flavus, which showed a decline in optimal activity above 50 °C, pH 5.5 [42]. The optimum temp of GA for soluble starch was reported as 55 °C from Gymnoascella citrina [44], whereas, the purified intracellular GA from A. tritic i WZ99 gave temp optimum of 45 °C [48].
Michaelis Menten kinetics constants (Km, Vmax, kcat & kcat/Km) of soluble starch hydrolysis by the mutant and parent Koji GAs were determined in the current study. The Km of mutant M-60(5) GA for soluble starch hydrolysis at 50 °C and pH 5.0 was lower (0.06 mg ml^‒1^) than the parental GA having 0.17 mg ml^‒1^. Previous studies reported lesser km values of naïve and modified enzyme, i.e., 0.34 and 0.29, respectively [49] and 2.1mg/ml of glucoamylase GA-LZ2 than the km values reported in this study [40]. The Kcat (turn over) catalytic events performed by active site of mutated GA were 1.7 fold higher than the parent GA, i.e., 118 s^‒1^. Moreover, turnover of the mutant GA was higher than that of recently reported novel mesophilic GA having 67.15 s^‒1^ [48] and GA of A. oryzae with 20.3s^‒1^ from Luzhou-flavour Daqu [40]. We believed that the mutation has altered the conformation of active site of mutant GA and made it more flexible and efficient to convert the transition ES * -complex into products. Furthermore, substrate specificity constant (kcat/Km) of mutated GA was 1899 and was 4.7 fold higher than control. Similar findings are reported in literature that γ-rays based mutagenesis of A. niger significantly improved the kinetic properties of GAs from mutant strains for starch hydrolysis [17]. Similar trend in kinetic parameters due to the γ-rays mediated mutagenesis in A. niger for lignocellulose hydrolysis by β-glycosidase was reported by Javed et al [58]. In another report Karim et al. [34] reported Km and Vmax for soluble starch as 5.84 mg ml^‒1^ and 153.85 U mg^‒1^ respectively for GAs from A. flavus NSH9. The current report indicates that mutant M-60(5) is highly substrate specific as compare to the already reported studies.
In addition, the catalytic efficiency of GA produce of A. oryzae mutant M-60(5) was extremely higher than the salt tolerant A. flavus, which had Km of 0.72 mg ml^‒1^ with Vmax of 12.48 μ mol min^‒1^ mg^‒1^. The Km of mutated GA (Km = 0.062 mg ml^‒1^) highlighted that its affinity to soluble starch was about 11.6 fold higher, while maximum velocity (Vmax = 112 μmol min^‒1^ mg^‒1^) was about 9.0 fold higher than that of A. flavus GA [42]. Hence, in comparison to previous studies, the kinetic properties of GA from mutant M-60(5) exhibited excellent catalytic activity in terms of starch hydrolysis that make efficient use of enzyme in industry. Similarly, we already reported γ-rays mutagenesis-based improvement in kinetic properties on thermodynamics of starch hydrolysis, where ΔH * (kJ mol^‒1^), ΔG * (kJ mol^‒1^), ΔS * (J mol^‒1^K^‒1^) for parent and mutant were 41.50, 46.12; 65.69, 63.62; ‒72.65, ‒52.53, respectively [17]. Hence, we concluded that the mutated GA was thermodynamically more efficient in conversion of soluble starch into products.
In the current study, the thermostability of mutant GA was increased about two fold at 55 °C than the parental enzyme, while at 45 °C & 50 °C a modest increase in the stability was observed (Fig 8 A, B). The GA from A. brasiliensis showed half-life of 22 min at 55 °C [50], while half-life of mutant GA of A. awamori at 55 °C was 48 min [51], which is less than that of mutant GA M-60(5). Previously, it was reported that γ-rays mutagenesis has affected A. niger in terms of thermodynamic parameters for cellobiose hydrolysis and improved the thermal stability of mutant enzyme. Where energy of activation (Ea_(d)_) of parent strain was 274 kJ mol^‒1^, while for mutant strain it was 240 kJ mol^‒1^. The ΔH * reported for mutant was as 237.02 kJ mol^‒1^, while for parent was 271.44 kJ mol^‒1^, whereas ΔG * was 105.67 kJ mol^‒1^ for mutant and 105.49 kJ mol^‒1^ for parent. The change in entropy (Δ*S**) for mutant was 407.90 J mol^‒1^K^‒1^, while for parent 515.35 J mol^‒1^K^‒1^ [52].
Conclusions
Random mutagenesis of A. oryzae by γ-ray treatment simultaneously enhanced the productivity, catalytic efficiency and thermostability of GA. The improvement in stability and function of GA was due to a point mutation in active site of GA encoding gene, resulted in replacement of amino acid Leu to Ile at position 203. Hence, the mutation changed the microenvironment of GA active site, resulting into alteration in active site conformation of mutant M-60(5) GA. The changed pKa values, ΔHi of acidic & basic limbs of the active site residues of mutated GA evidenced about the alteration of active site conformation. We concluded conformational change in active site of mutant GA was due to π MO ionization of aromatic histidine and NH_2_ nitrogen lone pair ionization of Glu/Asp residues. Thermostabilization of mutant GA was due to higher ΔG * . We concluded stability-function of enzymes might be simultaneously enhanced by strain improvement through γ-rays treatment. The mutated GA due to its high catalytic efficiency and thermostability proved that it has great potential for application in food industry such as beverages, baking and starch saccharification.
Supporting information
S1 FileRaw images.(PDF)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Zong X, Wen L, Wang Y, Li L. Research progress of glucoamylase with industrial potential. J Food Biochem. 2022;46(7):e 14099. doi: 10.1111/jfbc.14099 35132641 · doi ↗ · pubmed ↗
- 2Abalaka ME, Adetunji CO. Production and optimization of amylase and glucoamylase from Aspergillus niger under solid state fermentation for effective production of glucose syrup. UMYU J Microbiol Res. 2017;2(1):135–46. doi: 10.47430/ujmr.1721.021 · doi ↗
- 3Bilal M, Iqbal HM. State-of-the-art strategies and applied perspectives of enzyme biocatalysis in food sector—current status and future trends. Crit Rev Food Sci Nutr. 2020; 60 (12): 2052–66. doi: 10.1080/10408398.2019.1627284 31210055 · doi ↗ · pubmed ↗
- 4Song W, Tong Y, Li Y, Tao J, Li J, Zhou J, et al. Expression and characterization of a raw-starch glucoamylase from Aspergillus fumigatus. Process Biochem. 2021;11197–104. doi: 10.1016/j.procbio.2021.10.024 · doi ↗
- 5Ibrahim O. Functional oligosaccharide: chemicals structure, manufacturing, health benefits, applications and regulations. J Food Chem Nanotechol. 2018;04(04):. doi: 10.17756/jfcn.2018-060 · doi ↗
- 6Tong L, Zheng J, Wang X, Wang X, Huang H, Yang H, et al. Improvement of thermostability and catalytic efficiency of glucoamylase from Talaromyces leycettanus JCM 12802 via site-directed mutagenesis to enhance industrial saccharification applications . Biotechnol Biofuels. 2021; 14 (1): 1–9. doi: 10.1186/s 13068-021-02052-3 34656167 PMC 8520190 · doi ↗ · pubmed ↗
- 7Abou-Zeid AM, Abd El-Zaher EH, Saad-Allah KM, Ahmed RU. Screening and optimization of some Egyptian soil-born fungi for lipids production as possible source for biofuel. Egypt J Exp Biol (Bot). 2019;(0):1. doi: 10.5455/egyjebb.20190709123823 · doi ↗
- 8Roth C, Moroz OV, Turkenburg JP, Blagova E, Waterman J, Ariza A, et al. Structural and functional characterization of three novel fungal amylases with enhanced stability and p H tolerance. Int J Mol Sci. 2019;20(19):4902. doi: 10.3390/ijms 201949023162330931623309 PMC 6801514 · doi ↗ · pubmed ↗