Prostaglandins limit nuclear actin rod formation during Drosophila oogenesis
Tina L. Tootle

TL;DR
This study shows that prostaglandins help control nuclear actin rod formation in fruit fly egg development.
Contribution
The study demonstrates that prostaglandin signaling limits nuclear actin rod formation in Drosophila oogenesis.
Findings
Loss of prostaglandin synthesis increases the frequency of nuclear actin rods in Drosophila oogenesis.
PG signaling affects rod length and number, suggesting a regulatory role in nuclear actin dynamics.
Abstract
Expression of GFP-Actin results in nuclear actin rod formation during specific stages of Drosophila melanogaster oogenesis. Loss of prostaglandin (PG) synthesis and signaling results in an increased frequency of cells with nuclear actin rods; there are less rods per cell, but the rods are longer. These findings suggest that loss of PGs results in increased nuclear actin and are consistent with prior findings assessing the roles of PGs in modulating endogenous nuclear actin. Thus, GFP-Actin rod formation can be used as a tool to screen for new regulators of nuclear actin.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
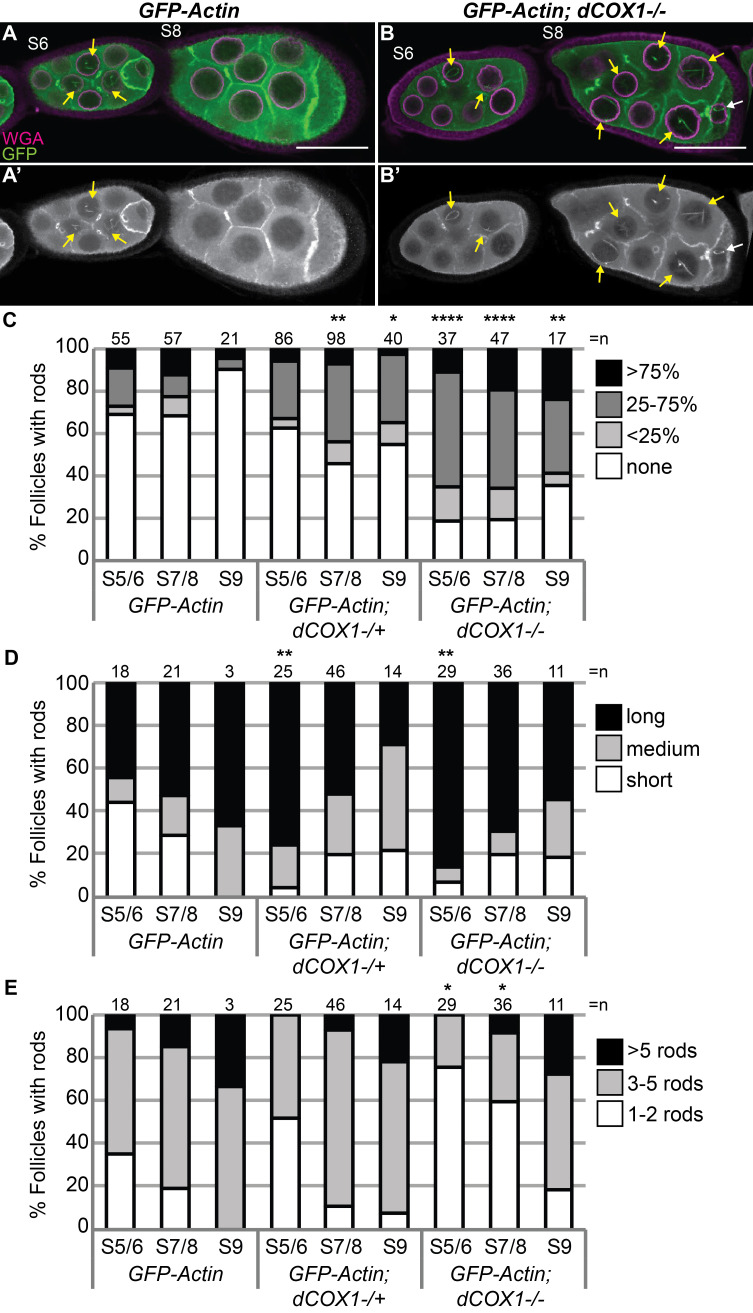 Figure 1
Figure 1|
| |||
|
|
|
|
|
|
|
y 1 w 1 |
RRID: BDSC_1495 |
Bloomington Drosophila Stock Center (BDSC) |
|
|
w 1118 ; P{osk-GAL4::VP16}A11/CyO |
FBtp0083699 Also: RRID: BDSC_44241 |
from A. Ephrussi Also available from: BDSC |
|
UAS-
|
w[*]; P{w[+mC]=UASp-GFP.Act5C}2-1 |
RRID: BDSC_9258 |
BDSC |
|
(referred to as
|
PBac{WH}pxt f01000 |
FBal0178293 |
Harvard Exelixis Collection; Thibault, 2004 PMID: 14981521 |
|
| |||
|
|
|
| |
|
rabbit polyclonal anti-GFP (TP401) |
RRID: AB_10013661 |
Torry Pines Biolabs, Inc | |
|
| |||
|
|
|
| |
|
AlexaFluor 555 Wheat Germ Agglutinin |
Cat# |
ThermoFisher Invitrogen | |
|
4’,6-Diamidino-2-phenylindole (DAPI) |
Cat# D9542 |
Millipore Sigma | |
|
bovine serum albumin |
Cat# A7284 |
Sigma-Aldrich | |
|
Grace's Insect Media |
Cat# 04-457F |
Lonza | |
- —U.S. National Science Foundation (United States)https://ror.org/021nxhr62
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsHER2/EGFR in Cancer Research · Colorectal Cancer Treatments and Studies · Cardiac Imaging and Diagnostics
Description
Actin localizes and functions within the nucleus. The import and export of actin from the nucleus is highly regulated (Borkuti et al., 2022; Dopie et al., 2012; Stuven et al., 2003), supporting that it has important nuclear functions. Indeed, recent work reveals nuclear actin has a wide variety of activities, including regulating transcription, being a component of chromatin remodeling complexes, mediating DNA damage repair, and contributing to nuclear structure (Green et al., 2021; Hurst et al., 2019; Kelpsch & Tootle, 2018; Misu et al., 2017). The functions of nuclear actin, like cytoplasmic actin, are regulated by its form – monomeric, polymeric, filamentous, and networks of filaments (Fukui & Katsumaru, 1979; Grosse & Vartiainen, 2013; Hendzel, 2014; Kristo et al., 2016; Vartiainen, 2008). In some cases, thick nuclear actin filaments are observed; these are often referred to as nuclear actin rods. Such rods form when there is a large influx of actin into the nucleus; this occurs in the contexts of cellular stress (Fukui & Katsumaru, 1979; Iida et al., 1986; Munsie et al., 2012; Nishida et al., 1987; Osborn & Weber, 1980; Vartiainen et al., 2007). Thus, nuclear actin rods are indicative of high nuclear actin levels. While nuclear actin plays critically important roles in cellular functions, the mechanisms regulating its level and form remain poorly understood.
We established Drosophila oogenesis as a new, in vivo system for studying nuclear actin regulation and function. Female flies have two ovaries, each of which contains 15-20 ovarioles or chains of sequentially developing egg chambers or follicles. There are fourteen stages of follicle development (Giedt & Tootle, 2023). Each follicle is comprised of one oocyte, fifteen germline-derived nurse cells and a layer of epithelial cells termed follicle cells. We previously found that during follicle development there are multiple pools of endogenous nuclear actin that exhibit unique developmental patterns (Kelpsch et al., 2016; Wineland et al., 2018). Further, germline expression of exogenous actin labeling tools (Lifeact-GFP, Utrophin-GFP, and GFP-Actin) perturbs nuclear actin dynamics and results in stage-dependent nuclear actin rod formation (Kelpsch et al., 2016; Spracklen, Fagan, et al., 2014). These rods form due to the tools artificially stabilizing endogenous nuclear actin (Lifeact-GFP or Utrophin-GFP) and/or increasing nuclear actin levels (GFP-Actin).
Using Drosophila oogenesis as a model, we identified prostaglandin (PGs) signaling as a novel regulator of nuclear actin. PGs are locally acting lipid signaling molecules that are produced in a multi-step process (Funk, 2001; Tootle, 2013). Briefly, the fatty acid arachidonic acid, the substrate for PG production, is enzymatically released from lipid stores. This substrate is acted on by cyclooxygenase (COX) enzymes and PG-type specific synthases to produce bioactive PGs; these PGs then activate G protein-coupled receptors to mediate their downstream functions. Flies have a single COX-like enzyme termed Pxt/dCOX1, subsequently referred to as dCOX1 (Tootle & Spradling, 2008). We previously found that during Drosophila oogenesis PGs regulate both actin cytoskeletal remodeling and nuclear actin (Groen et al., 2012; Spracklen, Kelpsch, et al., 2014; Talbot et al., 2023; Tootle & Spradling, 2008). Specifically, PG synthesis and signaling normally limits the amount and regulates the forms of nuclear actin (Talbot et al., 2023).
To further test the role of PG signaling in controlling nuclear actin, we asked whether PGs regulate nuclear actin rod formation. Germline expression of GFP-Actin increases nuclear actin levels, and this, in turn, induces nuclear actin rod formation in a low frequency of nurse cells during Stages 5-9 (S5-9) (Kelpsch et al., 2016). As loss of dCOX1 increases endogenous nuclear actin (Talbot et al., 2023), we hypothesized that *dCOX1 * mutant follicles will exhibit increased GFP-Actin rod formation. Indeed, that is what we observe (Fig 1B-B’ compared to A-A’). To quantify this change follicles were binned into 4 categories based on the percentage of nurse cells exhibiting actin rods – 0, <25%, 25-75%, and >75%. Both reduction in (heterozygosity) or loss of dCOX1 increases the frequency of follicles exhibiting actin rods (Fig 1C). We also assessed rod length (short, medium, or long), and number of rods per nuclei. Reduction or loss of dCOX1 results in longer rods during S5/6 (Fig 1D), and in *dCOX1 * mutants the number rods are fewer during S5/6 and S7/8 (Fig 1E). These data are consistent with the idea that small rods are joined to create larger and fewer rods as nuclear actin levels increase (Ishikawa-Ankerhold et al., 2017); however additional experiments are needed to formally test this possibility. The increased GFP-Actin rod formation observed when dCOX1 is lost correlates with our prior finding that PG synthesis and signaling are required to limit the level of endogenous nuclear actin (Talbot et al., 2023). Therefore, alterations in GFP-nuclear actin rod formation can be used to screen for regulators of nuclear actin.
Methods
Fly stocks
Fly stocks were maintained on cornmeal-agar-yeast food at 21°C. The following stocks were obtained from the Bloomington Drosophila Stock Center (Bloomington, IN): yw (RRID: BDSC_1495) and UASp GFP-Actin5C (RRID: BDSC_9258). * pxt ^f01000^ * (referred to as dCOX1-/- ) was obtained from the Harvard Exelixis collection (Boston, MA). The *oskar GAL4 * line was a generous gift from Anne Ephrussi (Telley et al., 2012); this stock can also be obtained from the Bloomington Drosophila Stock Center (RRID: BDSC_44241). Expression of UASp GFP-Actin 5C was achieved by crossing to oskar GAL4 flies, maintaining fly crosses at 21°C, and maintaining progeny at 25°C for 5-6 days, during which the flies were fed wet yeast paste daily.
Immunofluorescence
Whole-mount
- Drosophila* ovary samples were dissected into room temperature Grace’s insect medium (Lonza, Walkersville, MD). Samples were fixed for 10 min at room temperature in 4% paraformaldehyde in Grace’s insect medium. Samples were blocked by washing in antibody wash (1X phosphate-buffered saline, 0.1% Triton X-100, and 0.1% bovine serum albumin) six times for 10 mins each at room temperature. The primary antibody was incubated overnight at 4°C; rabbit anti-GFP 1:2000 (pre-absorbed on *yw * ovaries at 1:20 and used at 1:100; RRID: AB_10013661, Torrey Pines Biolabs, Inc., Secaucus, NJ). After 6 washes in antibody wash (10 min each), secondary antibodies were incubated overnight at 4°C or for ~4 hours at room temperature. The following secondary antibody was used at 1:250-1:500: AlexaFluor 488:goat anti-rabbit (RRID:AB_2576217). AlexaFluor 555-conjugated wheat germ agglutinin, WGA (1:500; ThermoFisher Invitrogen) was included with the primary and secondary antibodies. Following six washes in antibody wash (10 min each), 4′,6-diamidino-2-phenylindole (DAPI, 5 mg/ml) staining was performed at a concentration of 1:5000 in 1X PBS for 10 min at room temperature. Samples were then rinsed in 1X PBS and mounted in 1 mg/mL phenylenediamine in 50% glycerol, pH 9 (Platt & Michael, 1983). All experiments were performed a minimum of three independent times.
Image acquisition and processing
Microscope images of fixed Drosophila follicles were obtained using Zen software on a Zeiss 700 mounted on an Axio Observer.Z1 using a Plan-Aprochromat 20x/0.8 WD=.55 M27 (Carl Zeiss Microscopy, Thornwood, NY). Maximum projections, merged images, rotation, and cropping were performed using ImageJ software. All fluorescent images were brightened by 30% in Photoshop (Adobe) to improve visualization.
Quantification of nuclear actin rod frequency, length, and number
Quantification of nuclear actin rods was performed on confocal image stacks of follicles stained with anti-GFP, WGA, and Phalloidin. Genotypically de-identified images were analyzed using ImageJ; as necessary, brightness and contrast were adjusted to score all the actin rods present. Data was collected for S5-6, S7-8, and S9; follicle staging was assigned based on morphology and size. For each follicle the percentage of nurse cells exhibiting nuclear actin rods was assessed and binned into four categories: none, ≤25%, 25-75%, or ≥75%. For each follicle with nurse cells exhibiting nuclear actin rods, the number of rods was counted and binned into 3 categories: 1 to 2 rods, 3 to 5 rods, or >5 rods per nuclei. The rods were then scored for length: short (≤1/4 diameter of nucleus), medium (~1/2 diameter of nucleus), or long (≥1 diameter). Data was analyzed using Excel (Microsoft) and statistical analysis was preformed using R (Vienna, Austria).
Reagents
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Borkúti P KristóI SzabóA Bajusz C Kovács Z Réthi-Nagy Z Lipinszki Z Lukácsovich T Bogdan S Vilmos P 2022819 Parallel import mechanisms ensure the robust nuclear localization of actin in Drosophila.Front Mol Biosci 92296-889X 96363596363510.3389/fmolb.2022.96363536060241 PMC 9437273 · doi ↗ · pubmed ↗
- 2Dopie J Skarp KP RajakyläEK TanhuanpääK Vartiainen MK 201229 Active maintenance of nuclear actin by importin 9 supports transcription.Proc Natl Acad Sci U S A 10990027-8424 E 544E 55210.1073/pnas.111888010922323606 PMC 3295300 · doi ↗ · pubmed ↗
- 3Fukui Y Katsumaru H 197951 Nuclear actin bundles in Amoeba, Dictyostelium and human He La cells induced by dimethyl sulfoxide.Exp Cell Res 12020014-482745145510.1016/0014-4827(79)90412-9571346 · doi ↗ · pubmed ↗
- 4Funk CD 20011130 Prostaglandins and leukotrienes: advances in eicosanoid biology.Science 29455480036-80751871187510.1126/science.294.5548.187111729303 · doi ↗ · pubmed ↗
- 5Giedt MS Tootle TL 2023 The Vast Utility of Drosophila Oogenesis.Methods Mol Biol 26261064-374513610.1007/978-1-0716-2970-3_136715897 · doi ↗ · pubmed ↗
- 6Green Nicole M Kimble Garrett C Talbot Danielle E Tootle Tina L 2021323 Nuclear Actin Encyclopedia of Life Sciences 95896710.1002/9780470015902.a 0028471 · doi ↗
- 7Groen CM Spracklen AJ Fagan TN Tootle TL 20121010 Drosophila Fascin is a novel downstream target of prostaglandin signaling during actin remodeling.Mol Biol Cell 23231059-15244567457810.1091/mbc.E 12-05-041723051736 PMC 3510018 · doi ↗ · pubmed ↗
- 8Grosse R Vartiainen MK 2013103 To be or not to be assembled: progressing into nuclear actin filaments.Nat Rev Mol Cell Biol 14111471-007269369710.1038/nrm 368124088744 · doi ↗ · pubmed ↗