Complete mitogenome of the critically endangered Asian king vulture (Sarcogypscalvus) (Aves, Accipitriformes, Accipitridae): evolutionary insights and comparative analysis
Wannapol Buthasane, Sithichoke Tangphatsornruang, Piroon Jenjaroenpun, Thidathip Wongsurawat, Saowaphang Sanannu, Vorasuk Shotelersuk, Gunnaporn Suriyaphol

TL;DR
This study sequenced the complete mitochondrial genome of the critically endangered Asian king vulture and analyzed its evolutionary relationships with other vulture species.
Contribution
The first complete mitogenome of Sarcogypscalvus is assembled and compared with other vulture groups.
Findings
The mitogenome of S.calvus is 17,750 base pairs long and includes 13 protein-coding genes.
S.calvus is closely related to other Gypini vultures, with a divergence time of 16.7 million years ago.
Conserved amino acid substitutions in the ND1 gene distinguish Gypini from Cathartidae vultures.
Abstract
The Asian king vulture (Sarcogypscalvus), also known as the red-headed vulture, is an Old World vulture (Gypini) facing severe population declines. This study aimed to assemble the complete mitogenome of S.calvus, explore its phylogenetic relationships, estimate divergence times, and examine genetic distances and amino acid substitutions. The mitogenome was de novo assembled from genomic DNA extracted from the blood of a female S.calvus. Phylogenetic and pairwise genetic distance analyses were conducted, comparing S.calvus with other members of Gypini, New World vultures (Cathartidae) and various other birds. The assembled mitogenome was 17,750 base pairs in length, comprising 13 protein-coding genes (PCGs), 22 transfer RNA genes, two ribosomal RNA genes and two control regions. Most PCGs used the ATG start codon, except for cytochrome c oxidase subunit 1 (COX1), which employed GTG.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
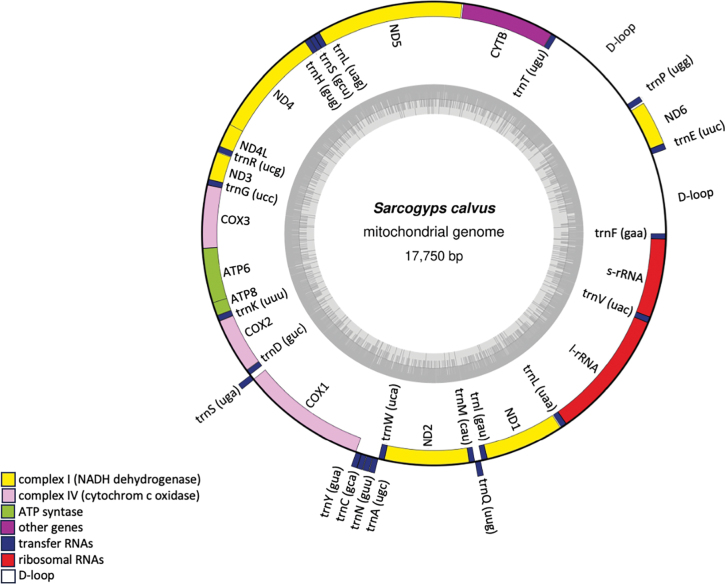 Figure 1
Figure 1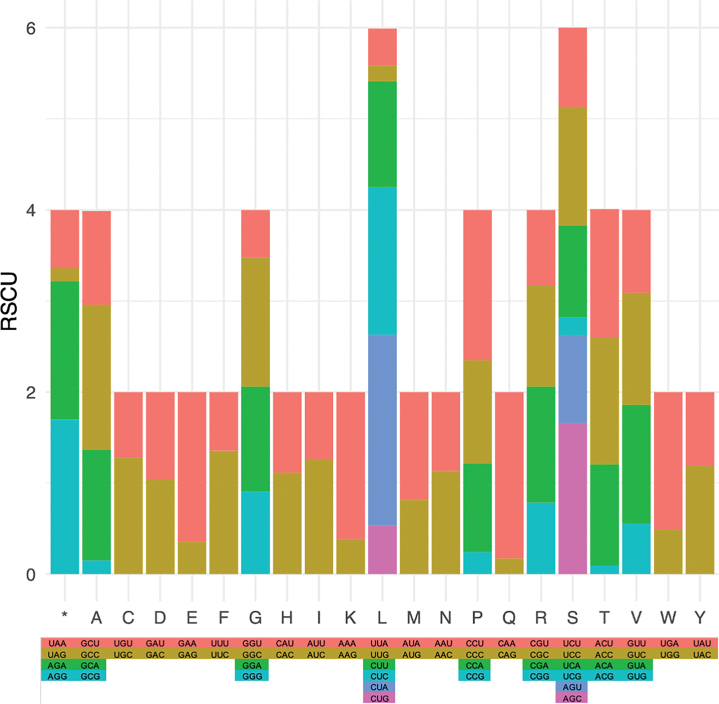 Figure 2
Figure 2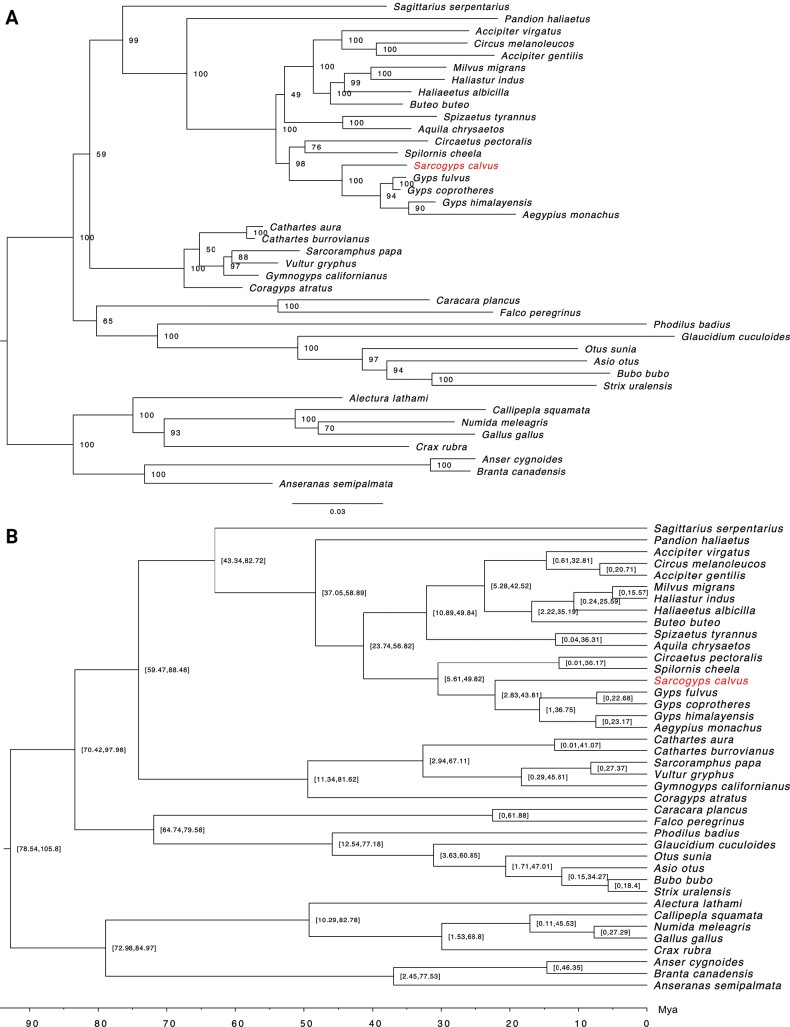 Figure 3
Figure 3| Scientific name | Order | Family | Accesion number | Sequence length ( | Reference |
|---|---|---|---|---|---|
|
|
|
|
| 18,266 | Unpublished |
|
|
|
|
| 17,952 |
|
|
|
|
|
| 17,811 |
|
|
|
|
|
| 17,332 |
|
|
|
|
|
| 18,674 |
|
|
|
|
|
| 17,473 |
|
|
|
|
|
| 17,749 | Unpublished |
|
|
|
|
| 16,908 |
|
|
|
|
|
| 18,094 |
|
|
|
|
|
| 17,381 |
|
|
|
|
|
| 17,719 |
|
|
|
|
|
| 19,055 |
|
|
|
|
|
| 18,016 |
|
|
|
|
|
| 18,291 | Unpublished |
|
|
|
|
| 17,479 |
|
|
|
|
|
| 19,285 |
|
|
|
|
|
| 19,329 |
|
|
|
|
|
| 19,302 |
|
|
|
|
|
| 16,808 |
|
|
|
|
|
| 16,870 | Harrison et al. 2004 |
|
|
|
|
| 16,870 |
|
|
|
|
|
| 16,779 |
|
|
|
|
|
| 17,864 |
|
|
|
|
|
| 16,760 |
|
|
|
|
|
| 16,773 |
|
|
|
|
|
| 16,739 |
|
|
|
|
|
| 17,077 |
|
|
|
|
|
| 18,068 |
|
|
|
|
|
| 16,698 |
|
|
|
|
|
| 16,666 |
|
|
|
|
|
| 16,701 |
|
|
|
|
|
| 16,784 | Unpublished |
|
|
|
|
| 16,785 | Unpublished |
|
|
|
|
| 17,555 |
|
|
|
|
|
| 18,952 |
|
|
|
|
|
| 17,392 | Unpublished |
|
|
|
|
| 17,835 |
|
|
|
|
|
| 18,708 |
|
|
|
|
|
| 17,086 |
|
| Start | End | Length ( | Direction | Type | Gene name | Gene product | Anti-codon | Start codon | Stop codon |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 987 | 987 | + | control region | - | - | – | – | – |
| 988 | 1058 | 71 | + | tRNA | tRNA-Glu | TTC | – | – | |
| 1059 | 1577 | 519 | + | CDS |
| NADH dehydrogenase subunit 6 | – | ATG | TAG |
| 1599 | 1668 | 70 | + | tRNA | tRNA-Pro | TGG | – | – | |
| 1669 | 2863 | 1195 | + | control region | |||||
| 2931 | 2864 | 68 | – | tRNA | tRNA-Thr | TGT | |||
| 4076 | 2934 | 1143 | – | CDS |
| cytochrome b | – | ATG | TAA |
| 5903 | 4089 | 1815 | – | CDS |
| NADH dehydrogenase subunit 5 | – | ATG | TAA |
| 5974 | 5904 | 71 | – | tRNA | tRNA-Leu | TAG | – | – | |
| 6039 | 5975 | 65 | – | tRNA | tRNA-Ser | GCT | – | – | |
| 6110 | 6041 | 70 | – | tRNA | tRNA-His | GTG | – | – | |
| 7488 | 6106 | 1383 | – | CDS |
| NADH dehydrogenase subunit 4 | – | ATG | AGG |
| 7778 | 7482 | 297 | – | CDS |
| NADH dehydrogenase subunit 4L | – | ATG | TAA |
| 7848 | 7780 | 69 | – | tRNA | tRNA-Arg | TCG | – | – | |
| 8204 | 7854 | 354 | – | CDS |
| NADH dehydrogenase subunit 3 | – | ATG | AGG |
| 8273 | 8205 | 69 | – | tRNA | tRNA-Gly | TCC | – | – | |
| 9057 | 8274 | 784 | – | CDS |
| cytochrome c oxidase subunit III | – | ATG | CCT |
| 9740 | 9057 | 684 | – | CDS |
| ATP synthase F0 subunit 6 | – | ATG | TAA |
| 9898 | 9731 | 168 | – | CDS |
| ATP synthase F0 subunit 8 | – | ATG | TAA |
| 9970 | 9900 | 71 | – | tRNA | tRNA-Lys | TTT | – | – | |
| 10655 | 9972 | 684 | – | CDS |
| cytochrome c oxidase subunit II | – | ATG | TAA |
| 10726 | 10658 | 69 | – | tRNA | tRNA-Asp | GTC | – | – | |
| 10731 | 10802 | 72 | + | tRNA | tRNA-Ser | TGA | – | – | |
| 12344 | 10794 | 1551 | – | CDS |
| cytochrome c oxidase subunit I | – | GTG | AGG |
| 12346 | 12415 | 70 | + | tRNA | tRNA-Tyr | GTA | – | – | |
| 12416 | 12482 | 67 | + | tRNA | tRNA-Cys | GCA | – | – | |
| 12485 | 12557 | 73 | + | tRNA | tRNA-Asn | GTT | – | – | |
| 12560 | 12628 | 69 | + | tRNA | tRNA-Ala | TGC | – | – | |
| 12702 | 12630 | 73 | – | tRNA | tRNA-Trp | TCA | – | – | |
| 13747 | 12701 | 1047 | – | CDS |
| NADH dehydrogenase subunit 2 | – | ATG | TAG |
| 13816 | 13748 | 69 | – | tRNA | tRNA-Met | CAT | – | – | |
| 13816 | 13886 | 71 | + | tRNA | tRNA-Gln | TTG | – | – | |
| 13971 | 13900 | 72 | – | tRNA | tRNA-Ile | GAU | – | – | |
| 14947 | 13970 | 978 | – | CDS |
| NADH dehydrogenase subunit 1 | – | ATG | AGG |
| 15030 | 14957 | 74 | – | tRNA | tRNA-Leu | TAA | – | – | |
| 16634 | 15030 | 1605 | – | rRNA | 16S ribosomal RNA | – | – | – | |
| 16706 | 16635 | 72 | – | tRNA | tRNA-Val | TAC | – | – | |
| 17681 | 16706 | 976 | – | rRNA |
| 12S ribosomal RNA | – | – | – |
| 17750 | 17681 | 70 | – | tRNA | tRNA-Phe | GAA | – | – |
| Region | MRS | DRS | TriRS | TetRS | Microsatellite sequences | Total |
|---|---|---|---|---|---|---|
| (unidentified region) | 7 | 0 | 4 | 0 | 2 | 13 |
|
| 7 | 18 | 14 | 3 | 3 | 45 |
|
| 4 | 4 | 5 | 1 | 0 | 14 |
|
| 11 | 56 | 28 | 7 | 2 | 104 |
|
| 7 | 15 | 14 | 4 | 1 | 41 |
|
| 10 | 27 | 16 | 2 | 1 | 56 |
|
| 22 | 46 | 15 | 3 | 2 | 88 |
|
| 24 | 44 | 16 | 2 | 4 | 90 |
|
| 18 | 37 | 16 | 6 | 3 | 80 |
| 43 | 49 | 18 | 5 | 0 | 115 | |
|
| 17 | 38 | 15 | 2 | 3 | 75 |
|
| 18 | 28 | 20 | 4 | 5 | 75 |
|
| 3 | 9 | 3 | 2 | 0 | 17 |
|
| 27 | 41 | 34 | 2 | 1 | 105 |
|
| 1 | 10 | 3 | 2 | 0 | 16 |
|
| 32 | 53 | 41 | 7 | 2 | 135 |
|
| 14 | 10 | 12 | 5 | 5 | 46 |
|
| 20 | 22 | 10 | 3 | 1 | 56 |
|
| 1 | 1 | 0 | 1 | 0 | 3 |
|
| 0 | 3 | 0 | 0 | 0 | 3 |
|
| 1 | 1 | 0 | 0 | 0 | 2 |
| 2 | 1 | 1 | 0 | 0 | 4 | |
|
| 2 | 5 | 2 | 0 | 0 | 9 |
| 1 | 1 | 0 | 2 | 0 | 4 | |
|
| 0 | 4 | 0 | 0 | 0 | 4 |
|
| 1 | 2 | 4 | 0 | 0 | 7 |
|
| 1 | 3 | 0 | 0 | 0 | 4 |
| 1 | 0 | 0 | 0 | 0 | 1 | |
|
| 2 | 0 | 2 | 0 | 0 | 4 |
|
| 1 | 0 | 0 | 1 | 0 | 2 |
| 2 | 2 | 0 | 0 | 0 | 4 | |
|
| 2 | 3 | 2 | 0 | 0 | 7 |
|
| 0 | 1 | 0 | 1 | 0 | 2 |
|
| 0 | 1 | 1 | 2 | 0 | 4 |
| 2 | 1 | 0 | 0 | 0 | 3 | |
| 2 | 4 | 0 | 0 | 0 | 6 | |
|
| 0 | 1 | 3 | 2 | 0 | 6 |
|
| 1 | 2 | 0 | 0 | 0 | 3 |
|
| 4 | 0 | 0 | 0 | 0 | 4 |
| Total | 311 | 543 | 299 | 69 | 35 | 1257 |
| Gene | Position | OWV | NWV | Side chain property | |
|---|---|---|---|---|---|
| OWV | NWV | ||||
|
| 7 | A | N | Hydrophobic | Polar uncharged |
| 30 | P | S | Special case | Polar uncharged | |
|
| 1 | G | A | Special case | Hydrophobic |
| 3 | S | F | Polar uncharged | Hydrophobic | |
|
| 4 | H | N | Positive | Polar uncharged |
| 43 | A | T | Hydrophobic | Polar uncharged | |
| 90 | N | D | Polar uncharged | Negative | |
| 156 | N | G | Polar uncharged | Special case | |
| 161 | A | S | Hydrophobic | Polar uncharged | |
| 166 | A | T | Hydrophobic | Polar uncharged | |
|
| 152 | M | T | Hydrophobic | Polar uncharged |
| 224 | N | K | Polar uncharged | Positive | |
|
| 5 | P | I | Special case | Hydrophobic |
| 376 | M | T | Hydrophobic | Polar uncharged | |
| 378 | C | Y | Special case | Hydrophobic | |
| 381 | T | K | Polar uncharged | Positive | |
|
| 9 | H | Y | Positive | Hydrophobic |
| 79 | T | I | Polar uncharged | Hydrophobic | |
| 81 | M | T | Hydrophobic | Polar uncharged | |
| 160 | V | T | Hydrophobic | Polar uncharged | |
| 171 | A | T | Hydrophobic | Polar uncharged | |
| 173 | T | A | Polar uncharged | Hydrophobic | |
| 189 | T | A | Polar uncharged | Hydrophobic | |
| 260 | Q | E | Polar uncharged | Negative | |
| 263 | T | P | Polar uncharged | Special case | |
| 312 | I | T | Hydrophobic | Polar uncharged | |
| 323 | C | Y | Special case | Hydrophobic | |
|
| 5 | T | A | Polar uncharged | Hydrophobic |
| 56 | T | A | Polar uncharged | Hydrophobic | |
| 65 | T | A | Polar uncharged | Hydrophobic | |
| 140 | V | T | Hydrophobic | Polar uncharged | |
| 185 | A | S | Hydrophobic | Polar uncharged | |
| 229 | T | M | Polar uncharged | Hydrophobic | |
| 299 | H | Y | Positive | Hydrophobic | |
|
| 7 | T | M | Polar uncharged | Hydrophobic |
| 19 | I | T | Hydrophobic | Polar uncharged | |
| 64 | L | P | Hydrophobic | Special case | |
| 77 | T | P | Polar uncharged | Special case | |
|
| 8 | T | M | Polar uncharged | Hydrophobic |
| 40 | H | Q | Positive | Polar uncharged | |
| 57 | C | G | Special case | Special case | |
| 63 | S | A | Polar uncharged | Hydrophobic | |
| 112 | A | T | Hydrophobic | Polar uncharged | |
| 170 | H | Q | Positive | Polar uncharged | |
| 171 | I | T | Hydrophobic | Polar uncharged | |
| 195 | L | S | Hydrophobic | Polar uncharged | |
| 201 | M | T | Hydrophobic | Polar uncharged | |
|
| 11 | S | A | Polar uncharged | Hydrophobic |
| 16 | C | S | Special case | Polar uncharged | |
| 43 | T | A | Polar uncharged | Hydrophobic | |
|
| 16 | A | T | Hydrophobic | Polar uncharged |
| 61 | S | M | Polar uncharged | Hydrophobic | |
| 71 | I | T | Hydrophobic | Polar uncharged | |
| 332 | T | M | Polar uncharged | Hydrophobic | |
| 350 | A | N | Hydrophobic | Polar uncharged | |
| 382 | M | T | Hydrophobic | Polar uncharged | |
| 407 | A | T | Hydrophobic | Polar uncharged | |
| 438 | M | T | Hydrophobic | Polar uncharged | |
| 597 | T | I | Polar uncharged | Hydrophobic | |
|
| 36 | G | A | Special case | Hydrophobic |
| 50 | P | S | Special case | Polar uncharged | |
| 53 | A | S | Hydrophobic | Polar uncharged | |
| 73 | C | S | Special case | Polar uncharged | |
| 78 | L | P | Hydrophobic | Special case | |
| 107 | E | G | Negative | Special case | |
| 126 | V | S | Hydrophobic | Polar uncharged | |
| 140 | W | R | Hydrophobic | Positive | |
| Gene | Position | OWV | NWV | Side chain property | |
|---|---|---|---|---|---|
| OWV | NWV | ||||
|
| 31 | F | I | Hydrophobic | Hydrophobic |
| 83 | I | I | Hydrophobic | Hydrophobic | |
| 139 | I | I | Hydrophobic | Hydrophobic | |
|
| 24 | I | I | Hydrophobic | Hydrophobic |
| 50 | S | P | Polar uncharged | Special case | |
|
| 468 | M | M | Hydrophobic | Hydrophobic |
|
| 70 | I | I | Hydrophobic | Hydrophobic |
|
| 15 | I | I | Hydrophobic | Hydrophobic |
| 26 | P | S | Special case | Polar uncharged | |
| 47 | L | L | Hydrophobic | Hydrophobic | |
| 213 | I | V | Hydrophobic | Hydrophobic | |
| 220 | P | P | Special case | Special case | |
| 307 | F | F | Hydrophobic | Hydrophobic | |
| 310 | K | K | Positive | Positive | |
| 321 | L | L | Hydrophobic | Hydrophobic | |
| 370 | T | L | Polar uncharged | Hydrophobic | |
|
| 15 | S | S | Polar uncharged | Polar uncharged |
| 179 | L | L | Hydrophobic | Hydrophobic | |
|
| 19 | I | I | Hydrophobic | Hydrophobic |
| 22 | S | S | Polar uncharged | Polar uncharged | |
| 122 | S | S | Polar uncharged | Polar uncharged | |
| 325 | T | T | Polar uncharged | Polar uncharged | |
| 327 | I | T | Hydrophobic | Polar uncharged | |
| 335 | I | L | Hydrophobic | Hydrophobic | |
|
| 1 | I | I | Hydrophobic | Hydrophobic |
| 108 | T | N | Hydrophobic | Polar uncharged | |
|
| 43 | L | L | Hydrophobic | Hydrophobic |
| 90 | A | T | Hydrophobic | Polar uncharged | |
| 183 | H | P | Positive | Special case | |
| 263 | T | T | Polar uncharged | Polar uncharged | |
| 357 | T | T | Polar uncharged | Polar uncharged | |
| 394 | I | I | Hydrophobic | Hydrophobic | |
| 418 | T | T | Polar uncharged | Polar uncharged | |
|
| 73 | T | T | Polar uncharged | Polar uncharged |
|
| 30 | T | T | Polar uncharged | Polar uncharged |
| 74 | M | T | Hydrophobic | Polar uncharged | |
| 291 | T | T | Polar uncharged | Polar uncharged | |
| 404 | Y | Y | Hydrophobic | Hydrophobic | |
| 434 | E | G | Negative | Special case | |
| 600 | I | I | Hydrophobic | Hydrophobic | |
|
| 3 | A | T | Hydrophobic | Polar uncharged |
| 142 | A | A | Hydrophobic | Hydrophobic | |
| 166 | M | L | Hydrophobic | Hydrophobic | |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenetic diversity and population structure · Genomics and Phylogenetic Studies · Evolution and Paleontology Studies
Introduction
The Asian king vulture (Sarcogypscalvus Scopoli, 1786), also known as the red-headed vulture, belongs to the Old World vulture group (tribe Gypini) within the order Accipitriformes and the family Accipitridae. Classified as Critically Endangered by the International Union for Conservation of Nature (IUCN) Red List of Threatened Species, S.calvus is also listed under Appendix II of the Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES) (BirdLife International 2021; CITES 2024). Additionally, the species is protected under Thailand’s Wild Animal Conservation and Protection Act, B.E. 2562 (2019) (FAOLEX Database 2019). Gypini are distributed across Africa, Asia, and Europe, while New World vultures (Cathartidae) inhabit the Americas. Currently, the global population of S.calvus in the wild is estimated to be between 2,500 and 9,999 individuals, while in Thailand, the species is possibly extinct in the wild and only seven individuals remain in captivity (BirdLife International 2021; Buthasane et al. 2024).
The mitogenome (mitochondrial genome) is a valuable tool for investigating phylogenetic relationships, molecular identification, and adaptive evolution (De Panis et al. 2021; Kong et al. 2024). Mitogenomes of four species of Gypini have been reported, i.e., those of the Eurasian griffon (Gypsfulvus Hablizl, 1783), cape vulture (Gypscoprotheres Forster, 1798), cinereous vulture (Aegypiusmonachus Linnaeus, 1766) and Himalayan griffon (Gypshimalayensis Hume, 1869) (Li et al. 2015; Mereu et al. 2017; Jiang et al. 2019; Adawaren et al. 2020). For Cathartidae, seven mitogenomes from six species have been documented, i.e. the Andean condor (Vulturgryphus Linnaeus, 1758), king vulture (Sarcoramphuspapa Linnaeus, 1758), California condor (Gymnogypscalifornianus Shaw, 1797), lesser yellow-headed vulture (Cathartesburrovianus Cassin, 1845), turkey vulture (Cathartesaura Linnaeus, 1758) and black vulture (Coragypsatratus Bechstein, 1793) (Slack et al. 2007; De Panis et al. 2021; Urantówka et al. 2021). In addition, 11 mitogenomes from other species in the family Accipitridae have been reported, including those of the golden eagle (Aquilachrysaetos Linnaeus, 1758), common buzzard (Buteobuteo Linnaeus, 1758) and black kite (Milvusmigrans Boddaert, 1783) (Haring et al. 2001; Jeon et al. 2018; Mead et al. 2021). The genome of S.calvus has recently been published (Buthasane et al. 2024). However, no mitogenomic data are currently available for S.calvus, and its mitochondrial features remain understudied. This study aimed to elucidate the complete mitogenome of S.calvus and provide a comprehensive analysis of its structure, phylogenetic position, and the divergence time from other vultures. This research offers valuable insights into the mitochondrial profiles, evolutionary relationships, and population genetics of S.calvus in relation to other Gypini, Cathartidae and related species.
Materials and methods
Whole blood samples were obtained from a female S.calvus, approximately 25 years old, at Nakhon Ratchasima Zoo, the Zoological Park Organization of Thailand (ZPOT). Sampling was conducted in compliance with the ethical guidelines under the Chulalongkorn University Animal Care and Use Committee (CU-ACUC), Thailand (approval number 2131005). Total DNA was extracted from the whole blood sample using the Wizard HMW DNA Extraction Kit (Promega, Madison, WI, USA). The DNA concentration was determined using a NanoDrop One Microvolume UV-Vis Spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA).
The mitochondrial genome was sequenced using the short-read MGISEQ-2000 platform (MGI Tech, Shenzhen, China) and assembled with NOVOPlasty v. 3.8.2 (Dierckxsens et al. 2017). Annotation was carried out using the MITOS WebServer (Bernt et al. 2013). Protein-coding, rRNA and tRNA genes were further identified using the NCBI Basic Local Alignment Search Tool (BLAST) (Altschul et al. 1990). The circular structure of the mitogenome was visualized with OrganellarGenomeDRAW (OGDRAW) v. 1.3.122 (Greiner et al. 2019). Analyses of nucleotide and amino acid composition were conducted using MEGA X (Kumar et al. 2018). Simple sequence repeats (SSRs) of 1–6 bp in length were identified using the Microsatellite identification tool (Beier et al. 2017).
Mitogenomes from 39 bird species, representing the orders Accipitriformes (Old World vultures, hawks, eagles, and kites), Cathartiformes (New World vultures), Falconiformes (falcons), Strigiformes (owls), Anseriformes (ducks and relatives) and Galliformes (chickens and relatives), were used for phylogeny reconstruction, comparative codon usage analyses (Table 1) and divergence time estimation. These mitogenomes, along with the newly sequenced mitogenome of S.calvus, were analyzed. Multiple sequence alignments of 13 conserved protein-coding genes (PCGs)−cytochrome B (CYTB), NADH dehydrogenase subunits 1 (ND1), ND2, ND3, ND4, ND4L, ND5, ND6, cytochrome c oxidase subunits 1 (COX1), COX2, COX3, ATP synthase F0 subunit 6 (ATP6) and ATP8−were performed using PRANK v170427. The best-fit model, mtVer+I+R4, was selected using ModelFinder, and maximum likelihood phylogenies were constructed using IQ-TREE v. 2.2.0.3 with 1,000 ultrafast bootstrap replications (Nguyen LT et al. 2014; Hoang et al. 2017). The phylogenetic tree was visualized with Figtree v. 1.4.4 (Rambaut 2018). Several species of Anseriformes and Galliformes were used as outgroups. Divergence times were estimated using MCMCTree in the PAML 4.9j package, with the Hessian matrix computed via CODEML and a burn-in of 2,000 iterations. Fossil calibration times were obtained from the TimeTree database (Kumar et al. 2022). Genetic distance analyses were conducted using MEGA X and visualized using ggplot2 and ggtree in R (Wickham 2016; Yu et al. 2017; Kumar et al. 2018). The nomenclature for higher taxa follows Gregory et al. (2024).
Relative synonymous codon usage (RSCU) values for S.calvus mitochondrial protein-coding genes were calculated using MEGA X (Kumar et al. 2018). RSCU values reflect codon bias, with values greater than one indicating positive codon bias, values less than one indicating a negative codon bias, and values equal to one indicating random codon usage (Wong et al. 2010).
Amino acid sequences from 13 mitochondrial protein-coding genes in the mitogenomes of Gypini (Aegypiusmonachus, Gypscoprotheres, Gypsfulvus, Gypshimalayensis, S.calvus) and Cathartidae (Cathartesaura, Cathartesburrovianus, Coragypsatratus, Gymnogypscalifornianus, Sarcoramphuspapa, Vulturgryphus) were aligned using the Unipro UGENE Multiple Alignment Editor (Okonechnikov et al. 2012).
Results
The complete mitogenome of S.calvus was determined to be 17,750 base pairs (bp) in length and was assigned GenBank accession number OR896160. The circular structure of the mitogenome of S.calvus is illustrated in Fig. 1. This mitogenome contained 13 PCGs, 22 transfer RNA genes (tRNAs), two ribosomal RNA genes and two putative control regions (CRs), also referred to as D-loop regions (Table 2). The nucleotide composition was characterized by 54.1% adenine and thymine (AT) and 45.9% guanine and cytosine (GC).
Circular mitogenome map of Asian king vulture. The complex I (NADH dehydrogenase), complex IV (cytochrome c oxidase), ATP synthase, ribosomal RNAs, transfer RNAs, cytochrome b and control region (D-loop) are annotated. Genes located outside the circle are transcribed in a clockwise direction, whilefig. genes inside are transcribed counterclockwise. The inner ring shadow denotes the GC content of the genome.
The protein-coding regions spanned 11,407 bp, accounting for 64.26% of the length of the complete mitogenome of S.calvus. All PCGs, except for ND6, were transcribed on the plus strand. The predominant start codon for most PCGs was ATG, except for COX1, which utilized GTG as the start codon (Table 2). A detailed overview of the RSCU and codon distribution in the protein-coding genes of the mitogenome of S.calvus is provided. The codons CUA (L), CCU (P), and CUC (L) exhibited the highest frequency of occurrence (Fig. 2). A total of 1,257 SSRs were identified in the mitogenome, comprising 314 (24.98%) mono-, 529 (42.08%) di-, 301 (23.95%) tri-, 78 (6.21%) tetra-, 24 (1.91%) penta- and 11 (0.88%) hexanucleotide repeats. The ND5 gene contained the highest number of SSRs with 135 repeats (Table 3).
The relative synonymous codon usage (RSCU) and codon distribution of the Asian king vulture mitogenome. The different colors in the column chart symbolize distinct codon families associated with the amino acids listed below. Consistent coloring is applied to maintain representation uniformity across the same codon families. Bar chart showed-relative synonymous codon usage in all protein-coding genes of the mitogenome of S.calvus.
The mitogenome of S.calvus was aligned with 39 previously published mitogenomes of bird species from the orders Accipitriformes, Cathartiformes, Falconiformes, Strigiformes, Anseriformes and Galliformes. Maximum likelihood phylogenies are illustrated in Fig. 3. The mitogenome of S.calvus was part of a clade formed by the tribe Gypini (Gypsfulvus, Gypscoprotheres, Gypshimalayensis and Aegypiusmonachus). Gypini formed a sister group with the serpent-eagles of the tribe Circaetini (Spilornischeela Latham, 1790 and Circaetuspectoralis Smith, 1829). Gypini and Circaetini formed the sister-group of a clade comprising the subfamilies Accipitrinae and Aquilinae. The subfamily Aquilinae included the species Spizaetustyrannus Wied, 1820 and Aquilachrysaetos, whereas the subfamily Accipitrinae included the tribe Accipitrini (Accipitervirgatus Temminck, 1822, Accipitergentilis and Circusmelanoleucos Forster, 1795) and the tribe Buteonini (Milvusmigrans, Haliasturindus Boddaert, 1783, Haliaeetusalbicilla and Buteobuteo) (Fig. 3A). The estimated evolutionary divergence time between S.calvus and its sister taxa, based on mitochondrial data, was approximately 22.2 million years ago (Mya) (95% highest posterior density (HPD): 2.8–43.8 Mya) (Fig. 3B). Fossil calibration constraints were applied to several groups, including Accipitriformes and Falconiformes, as well as Accipitriformes and Galliformes, among others. The estimated divergence times for these groups ranged from 6.2 to 101.5 Mya. Genetic distance analysis showed that the genetic distance between S.calvus and other Gypini ranged from 4.02% to 5.17%, while the distance between S.calvus and Cathartidae ranged from 10.90% to 12.26% (Suppl. material 1: table S1).
Maximum-likelihood phylogenetic trees based on amino acid alignments of 13 conserved protein-coding genes from the mitochondrial genomes of Sarcogypscalvus and 39 other bird species, with species from the orders Galliformes and Anseriformes used as outgroups A phylogram indicating bootstrap support values based on 1,000 replicates at each node B time calibrated phylogeny with the estimated minimum and maximum divergence times (millions of years ago, Mya) are indicated at each node. Sarcogypscalvus is in red.
A total of 138 conserved amino acid substitution sites were observed between Old World vulture (Gypini) and New World vulture (Cathartidae) groups. The largest number of substitutions was found in the ND5 gene (23 sites), whereas the smallest number occurred in ATP6 (3 sites) (Suppl. material 1: table S2). Among these, 67 sites displayed substitutions between different amino acid chemical groups, with ND1 showing the highest number of such substitutions (11 sites). Special case amino acid substitutions were found in ATP8 (1 site), COX1 (1 site), COX2 (1 site), CYTB (2 sites), ND1 (2 sites), ND3 (2 sites), ND4 (1 site), ND4L (1 site) and ND6 (5 sites). Among these, Pro was the most frequently substituted amino acid, with 7 substitutions across ATP8, CYTB, ND1, ND3 and ND6, followed by Cys (5 sites) and Gly (4 sites) (Table 4). In the Old World vulture clade, 43 amino acid substitution sites were identified between S.calvus and other Gypini. The largest number of substitutions was found in CYTB (9 sites), while the smallest number occurred in COX1, COX2 and ND4L (1 site each) (Table 5). Unique amino acid chemical groups were found at 15 sites in S.calvus, with the largest number located in ND5 (5 sites). Pro was the most frequently substituted amino acid in this group (3 sites in ATP8, CYTB and ND4), with Gly ranking second (1 site in ND5) (Table 5).
Discussion
The present study has, for the first time, characterized the complete mitogenome of S.calvus and compared it with 39 other avian mitogenomes. The mitogenome of S.calvus included 13 PCGs, 22 tRNA genes, two rRNA genes and two putative CR regions, consistent with the mitogenomes of the Gypshimalayensis and Aepygiuscinereus (Li et al. 2015; Jiang et al. 2019). The total length of the PCG region of S.calvus was 11,407 base pairs (bp), which fell within the range observed in other members of Accipitriformes (11,377–11,920 bp). We observed that mitogenomes are subject to weaker translational selection compared to nuclear genomes (dos Reis et al. 2004). Regarding the GTG initiation codon of COX1, a previous study has reported the utilization of GTG as a start codon in the ribosomal protein L16 (rpl16) gene in some plant mitochondria (Bock et al. 1994). Additionally, RNA editing could also be a factor, as it has been observed in chicken mitochondria (Yokobori and Pääbo 1997). In the context of RSCU values, we identified AGG and AGA as the preferred stop codons in the mitogenome of S.calvus, with the RSCU values of 1.7 and 1.52, respectively. AGG has been identified as a stop codon for the NADH dehydrogenase 1 (ND1) and cytochrome c oxidase subunit 1(COX1) mitochondrial genes in the cinereous vulture and Himalayan griffon (Li et al. 2015; Jiang et al. 2019). Similarly, AGA has been shown to function as a stop codon for the NADH dehydrogenase subunit 3 (NADH3) and NADH dehydrogenase subunit 5 (NADH5) mitochondrial genes in the ostrich (Härlid et al. 1997). In contrast, UAA and UAG stop codons exhibited a negative bias, with RSCU values of 0.64 and 0.14, respectively. Additionally, we detected a negative bias against guanine at the third codon position across all 13 PCGs, consistent with findings in the cinereous vulture and Himalayan griffon (Li et al. 2015; Jiang et al. 2019).
Our results corroborate the position of S.calvus within the Old World vulture clade (Gypini), consistent with previous studies (Seibold and Helbig 1995; Lerner and Mindell 2005; Mindell et al. 2018; Khatri et al. 2019; Catanach et al. 2024). We estimated that S.calvus diverged from its sister clade (Gyps and Aegypius) approximately 22 Mya, while the divergence between Gypini and Cathartidae was estimated to have occurred around 74.1 Mya. This estimate closely aligns with previous findings (De Panis et al. 2021). Our analysis of amino acid substitutions, particularly those involving different chemical groups, suggests that these changes could potentially influence protein structure and function (Teng et al. 2010). Substitutions involving Cys, Pro, and Gly are particularly significant due to their unique roles in protein structure and stability. In this study, we observed transitions from Cys, which forms disulfide bonds critical for protein stability, to hydrophobic residues, potentially affecting protein folding and stability (Zavodszky et al. 2001; Alvares et al. 2013). Additionally, we detected changes involving Pro, known to restrict backbone flexibility, and Gly, which contributes to protein folding due to its small size, and may disrupt protein dynamics (Wilman et al. 2014; Senthil et al. 2019). We also noticed substitutions from hydrophobic to polar uncharged residues, such as Ser and Thr, across mitochondrial genes. These residues enhance hydrogen bonding and stability in aqueous environments, although transitions between similar residues (e.g., Ser to Thr) likely have minimal structural impact (Saeki et al. 2013). The observed amino acid changes may reflect functional adaptations and divergence within Gypini, with implications for mitochondrial function and the conservation of S.calvus. Future studies should explore these findings using structural modeling to better understand their impact.
Conclusions
Our study documents the characteristics of the complete mitogenome of S.calvus. Phylogenetic analysis corroborated its evolutionary relationships within Accipitriformes. S.calvus was most closely related to a clade formed by Aegypiusmonachus and species of Gyps. Additionally, we identified conserved amino acid changes between Gypini and Cathartidae, as well as unique amino acid substitutions specific to the S.calvus. These findings enhance our understanding of the evolutionary history and functional genomics of this critically endangered species.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Adawaren EO Du Plessis M Suleman E Kindler D Oosthuizen AO Mukandiwa L Naidoo V (2020) The complete mitochondrial genome of Gypscoprotheres (Aves, Accipitridae, Accipitriformes): phylogenetic analysis of mitogenome among raptors. Peer J 8: e 10034. 10.7717/peerj.10034 PMC 766654333240589 · doi ↗ · pubmed ↗
- 2Altschul SF Gish W Miller W Myers EW Lipman DJ (1990) Basic local alignment search tool. Journal of Molecular Biology 215: 403−410. 10.1016/S 0022-2836(05)80360-22231712 · doi ↗ · pubmed ↗
- 3Alvares RDA Tulumello DV Macdonald PM Deber CM Prosser RS (2013) Effects of a polar amino acid substitution on helix formation and aggregate size along the detergent-induced peptide folding pathway. Biochimica et Biophysica Acta 1828: 373−381. 10.1016/j.bbamem.2012.09.02423031573 · doi ↗ · pubmed ↗
- 4Beier S Thiel TMünch T Scholz U Mascher M (2017) MISA-web: a web server for microsatellite prediction. Bioinformatics 33: 2583−2585. 10.1093/bioinformatics/btx 198PMC 587070128398459 · doi ↗ · pubmed ↗
- 5Bernt M Donath AJühling F Externbrink F Florentz C Fritzsch GPütz J Middendorf M Stadler PF (2013) MITOS: improved de novo metazoan mitochondrial genome annotation. Molecular Phylogenetics and Evolution 69: 313−319. 10.1016/j.ympev.2012.08.02322982435 · doi ↗ · pubmed ↗
- 6Bird Life International (2021) Sarcogypscalvus. The IUCN Red List of Threatened Species 2021: e.T 22695254 A 205031246. http://datazone.birdlife.org/species/factsheet/red-headed-vulture-sarcogyps-calvus [accessed on 30 June 2024]
- 7Bock H Brennicke A Schuster W (1994) Rps 3 and rpl 16 genes do not overlap in Oenothera mitochondria: GTG as a potential translation initiation codon in plant mitochondria? Plant Molecular Biology 24: 811–818. 10.1007/BF 000298638193306 · doi ↗ · pubmed ↗
- 8Buthasane W Shotelersuk V Chetruengchai W Srichomthong C Assawapitaksakul A Tangphatsornruang S Pootakham W Sonthirod C Tongsima S Wangkumhang P Wilantho A Thongphakdee A Sanannu S Poksawat C Nipanunt T Kasorndorkbua C Koepfli K-P Pukazhenthi BS Suriyaphol P Wongsurawat T Jenjaroenpun P Suriyaphol G (2024) Comprehensive genome assembly reveals genetic diversity and carcass consumption insights in critically endangered Asian king vultures. Scientific Reports 14: 9455. 10.1038/s 41598-024-59990-9PMC 1104345038658744 · doi ↗ · pubmed ↗