Comprehensive Assessment of Initial Adaptation of Extended-Spectrum β-Lactamase–Positive ST131 Escherichia coli to Carbapenem Exposure
William C Shropshire, Xinhao Song, Jordan Bremer, Seokju Seo, Susana Rodriguez, Selvalakshmi Selvaraj Anand, An Q Dinh, Micah M Bhatti, Anna Konovalova, Cesar A Arias, Awdhesh Kalia, Yousif Shamoo, Samuel A Shelburne

TL;DR
This study explores how a high-risk strain of Escherichia coli adapts to carbapenem antibiotics, revealing that gene amplification is a key initial step toward resistance.
Contribution
The study identifies IS26-mediated gene amplification as a conserved initial adaptation mechanism in ESBL-positive ST131 E. coli under carbapenem exposure.
Findings
Initial carbapenem exposure leads to ESBL gene amplification via IS26-mediated transposons in ST131 C2/H30Rx isolates.
Increased ESBL gene copy number correlates strongly with higher mutation frequencies in ESBL-positive ST131 strains.
Stable porin gene mutations occur only after prolonged carbapenem exposure, suggesting a sequential adaptation process.
Abstract
It remains unclear how high-risk Escherichia coli lineages, like sequence type (ST) 131, initially adapt to carbapenem exposure in their progression to carbapenem resistance. Carbapenem mutation frequency was measured in multiple subclades of extended-spectrum β-lactamase (ESBL)–positive ST131 clinical isolates using a fluctuation assay followed by whole genome sequencing (WGS) characterization. Genomic, transcriptomic, and porin analyses of the ST131 C2/H30Rx isolate MB1860, under prolonged, increasing carbapenem exposure was performed using 2 experimental evolutionary platforms to measure fast versus slow adaptation. All 13 ESBL-positive ST131 strains selected from a diverse (n = 184) ST131 bacteremia cohort had detectable ertapenem (ETP) mutational frequencies, with a positive correlation between initial ESBL gene copy number and mutation frequency (r = 0.87, P < 1e-5). WGS…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
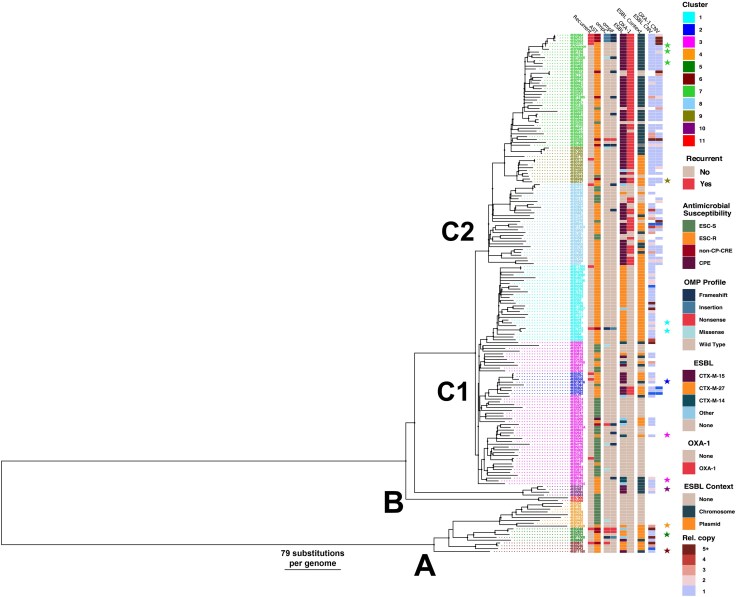 Figure 1
Figure 1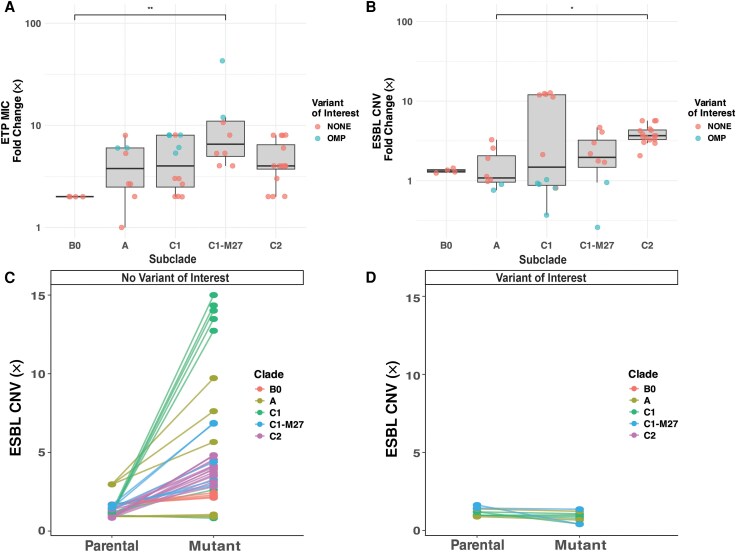 Figure 2
Figure 2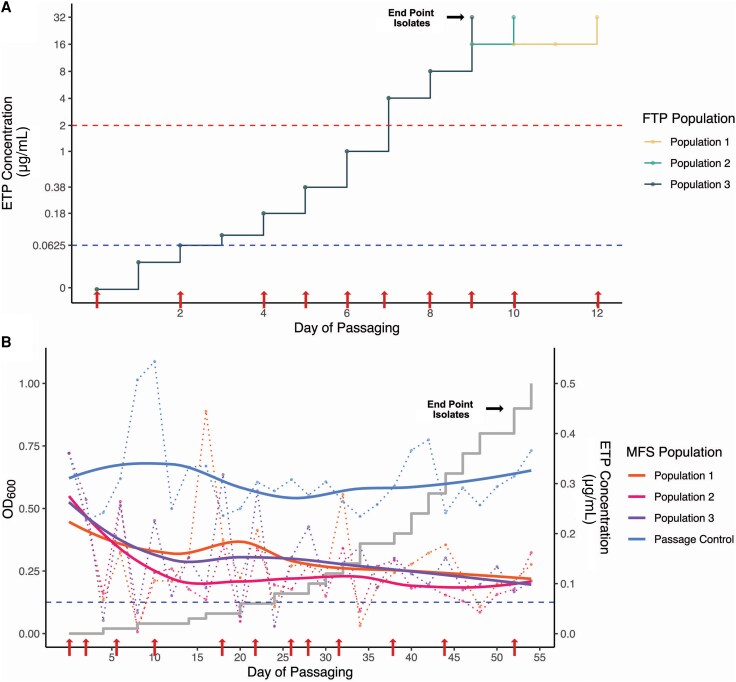 Figure 3
Figure 3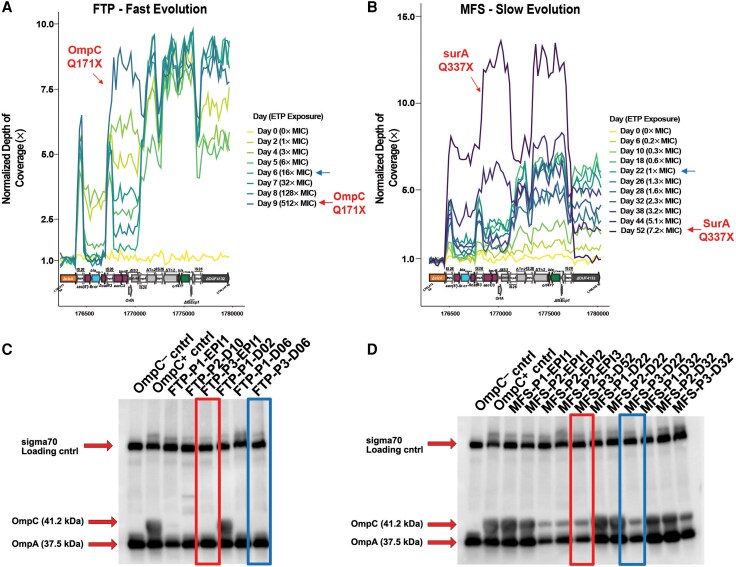 Figure 4
Figure 4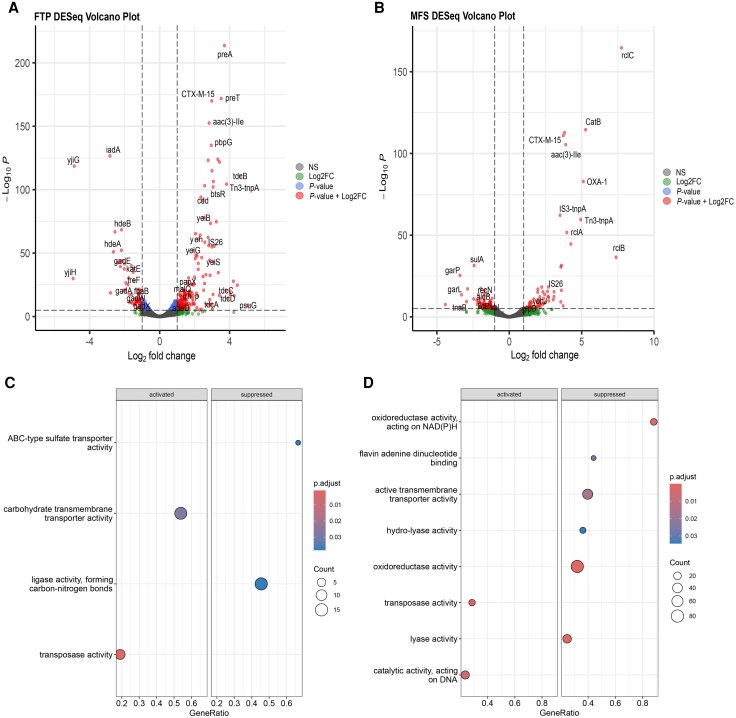 Figure 5
Figure 5- —National Institute of Allergy and Infectious Diseases10.13039/100000060
- —National Institutes of Health10.13039/100000002
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Antibiotics Pharmacokinetics and Efficacy · Antibiotic Use and Resistance
The increased worldwide prevalence of extended-spectrum β-lactamase (ESBL)–producing Escherichia coli (ESBL-Ec) represents a serious public health challenge, significantly contributing to the global burden of antimicrobial resistance (AMR)–related mortality [1]. A recent population-based United States (US) study indicated that from 2012 to 2017 there was a 53.3% increase in ESBL-producing Enterobacterales infections, with 87% of that increase due to ESBL-Ec infections [2]. Carbapenems are recommended for treating complicated ESBL-Ec infections [3], yet carbapenem resistance can still develop within these pathogens that lack carbapenem-degrading carbapenemases, driven by increased ESBL enzyme activity and altered outer membrane permeability that reduces intracellular carbapenem levels [4–6]. While these non-carbapenemase-producing, carbapenem-resistant E coli (non-CP-CREc) mechanisms are becoming more well understood, there remains limited knowledge of how ESBL-Ec strains initially adapt to carbapenem exposures prior to acquiring clinically defined carbapenem resistance.
Perhaps the most successful multidrug-resistant E coli lineage of the 21st century is the extraintestinal pathogenic sequence type (ST) known as ST131 [7–9]. Given this lineage's global dissemination and wide range of observed AMR phenotypes, ST131 has been considered an excellent model for examining AMR evolutionary trajectories [10–12]. Although there is significant spatiotemporal heterogeneity in population structure of carbapenem-resistant E coli [13, 14], a recent population-based US surveillance study of carbapenem-resistant Enterobacterales found that ST131 was the leading cause of carbapenem-resistant E coli infections with the vast majority having non-CP-CREc mechanisms [15].
Given the substantial prevalence of ST131 causing non-CP-CREc infections, we sought to perform a comprehensive assessment of ESBL-positive ST131 isolates’ initial adaptation to carbapenems using a multidisciplinary approach. Our study on ST131 bacteremia strains, particularly C2/H30Rx subclades, reveals that insertion sequence (IS)–mediated ESBL gene amplification is a key driver of initial carbapenem resistance. This response across multiple evolutionary models highlights a potential target for reducing resistance progression.
MATERIALS AND METHODS
Data Collection
ST131 E coli bacteremia events were abstracted from our REDCap database and cross-referenced with MicroBank isolates saved at −80°C in 20% glycerol stocks. Whole genome sequencing (WGS) results of E coli bacteremia isolates identified as ST131 from previous studies were included in the study [4, 5, 16–18].
Antimicrobial Susceptibility Testing
Clinical isolates were tested for their minimum inhibitory concentrations (MICs) to ertapenem (ETP) and meropenem (MEM) using both gradient strip Etest as well as broth microdilution following Clinical and Laboratory Standards Institute M100 (2022) guidelines [19].
Modified Luria Delbruck Assay
A modified fluctuation assay was performed for the purposes of measuring mutation frequency rate in our samples as previously described [20, 21]. Detail on assay and mutation frequency calculation is included in the Supplementary Material. For samples that had detectable positive growth, up to 4 mutants were randomly selected for ETP MIC determination using broth microdilution as previously described above and stored at −80°C for future genomic characterization as described below.
Microfluidic System and Flask Transfer Experimental Evolutionary Platform Protocols
The C2H30Rx strain MB1860 [4] was passaged using 2 distinct experimental evolutionary platforms. The novel microfluidic system (MFS) methodology has been published previously [22]. Further detail on the novel MFS system and the flask transfer protocol (FTP) methodology are included in the Supplementary Material. Daily populations as well as end point isolates (EPIs) collected from colonies streaked out from populations with last ETP exposure were collected for further characterization, described below.
DNA Sequencing
Mutants collected from our modified fluctuation assay of ST131 subclades (n = 52) and daily MB1860 population isolates collected from experimental evolution platforms (n = 64), respectively, were collected for Illumina short-read sequencing. Samples underwent genomic DNA extraction and Illumina short-read, paired-end sequencing DNA library prep as previously described [5]. A total of 4 samples (MB6054 [n = 4]; MB10029 [n = 1]) from the modified fluctuation assay were censored due to paired-end read contamination, leaving a total of 48 samples available for mutant analysis.
RNA Sequencing
Daily MB1860 population isolates from both experimental evolution platforms (ie, FTP and MFS) were selected for RNA sequencing (RNA-Seq). In particular, we selected daily populations that were exposed to approximately 1 × ETP MIC = 0.0625 μg/mL (ie, FTP population 1, day 2 isolates [FTP-P1-D2] and MFS population 1, day 26 [MFS-P1-D26] isolates; we collectively refer to these isolates as “experiment”), as well as passaged isolates collected from the same founder population and same day with no ETP exposure from each respective population (ie, FTP-P1-D2 and MFS-P1-D26 without ETP exposure). Three technical replicates from each group were grown to mid-log phase (optical density at 600 nm [OD_600_] = 0.5 ± 0.05) with the experimental isolates grown in Luria-Bertani broth supplemented with 0.03 μg/mL ETP whereas control isolates were grown without ETP exposure. Further detail on RNA-Seq methodology is included in the Supplementary Material.
Western Blot Analysis
Immunoblot analysis of outer membrane porins has been described previously [23]. In brief, samples are grown to mid-log phase and then standardized by OD_600_. Samples are then boiled and subjected to electrophoresis on sodium dodecyl sulfate polyacrylamide gel supplemented with 4M urea. Anti-rabbit antibodies targeting OmpC/OmpF/OmpA have been previously validated and reported [24, 25].
Comparative Genomics
A total of 183 ST131 E coli paired-end short-reads that were sequenced from previous molecular epidemiology projects [4, 5, 16–18] were aligned to MB1860 (RefSeq number NZ_CP049085.2) using the Snippy-v4.6.0 variant call pipeline (T. Seemann, https://github.com/tseemann/snippy). Core genome alignment file was used to infer a recombination-free maximum likelihood phylogenetic tree using UFBoot2 (1000X) and ModelFinder with IQTree-2-v2.0.3 [26] and Gubbins-v3.2.1 [27]. Further comparative genomics analysis detail can be found in the Supplementary Material.
Differential Expression Analysis
MB1860-passaged isolates were selected in the MFS and FTP system with and without ETP selective pressure (passage control, 0 µg/mL ETP; experiment, 0.0625 µg/mL ETP). A full description of differential expression and downstream RNA-seq analysis can be found in the Supplementary Material.
Statistical Analysis
All statistical analysis was performed in R version 4.3.2 software as previously described [5, 17, 18].
Supplementary Material
jiae587_Supplementary_Data
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Murray CJ , Ikuta KS, Sharara F, et al Global burden of bacterial antimicrobial resistance in 2019: a systematic analysis. Lancet 2022; 399:629–55.35065702 10.1016/S 0140-6736(21)02724-0PMC 8841637 · doi ↗ · pubmed ↗
- 2Jernigan JA , Hatfield KM, Wolford H, et al Multidrug-resistant bacterial infections in U.S. hospitalized patients, 2012–2017. N Engl J Med 2020; 382:1309–19.32242356 10.1056/NEJ Moa 1914433 PMC 10961699 · doi ↗ · pubmed ↗
- 3Tamma PD , Aitken SL, Bonomo RA, Mathers AJ, Van Duin D, Clancy CJ. Infectious Diseases Society of America guidance on the treatment of extended-spectrum β-lactamase producing Enterobacterales (ESBL-E), carbapenem-resistant Enterobacterales (CRE), and Pseudomonas aeruginosa with difficult-to-treat resistance (DTR-P. aeruginosa). Clin Infect Dis 2021; 72:e 169–83.33106864 10.1093/cid/ciaa 1478 · doi ↗ · pubmed ↗
- 4Shropshire WC , Aitken SL, Pifer R, et al IS 26-mediated amplification of bla OXA-1 and bla CTX-M-15 with concurrent outer membrane porin disruption associated with de novo carbapenem resistance in a recurrent bacteraemia cohort. J Antimicrob Chemother 2021; 76:385–95.33164081 10.1093/jac/dkaa 447PMC 7816169 · doi ↗ · pubmed ↗
- 5Shropshire WC , Konovalova A, Mc Daneld P, et al Systematic analysis of mobile genetic elements mediating beta-lactamase gene amplification in noncarbapenemase-producing carbapenem-resistant Enterobacterales bloodstream infections. Msystems 2022; 7:e 00476-22.36036505 10.1128/msystems.00476-22PMC 9601100 · doi ↗ · pubmed ↗
- 6Tangden T , Adler M, Cars O, Sandegren L, Lowdin E. Frequent emergence of porin-deficient subpopulations with reduced carbapenem susceptibility in ESBL-producing Escherichia coli during exposure to ertapenem in an in vitro pharmacokinetic model. J Antimicrob Chemother 2013; 68:1319–26.23478794 10.1093/jac/dkt 044 · doi ↗ · pubmed ↗
- 7Pitout JDD , Devinney R. Escherichia coli ST 131: a multidrug-resistant clone primed for global domination. F 1000 Res 2017; 6:195.10.12688/f 1000 research.10609.1PMC 533360228344773 · doi ↗ · pubmed ↗
- 8Johnson JR , Johnston B, Clabots C, Kuskowski MA, Castanheira M. Escherichia coli sequence type ST 131 as the major cause of serious multidrug-resistant E. coli infections in the United States. Clin Infect Dis 2010; 51:286–94.20572763 10.1086/653932 · doi ↗ · pubmed ↗