Epigenetic modulation of thyroid cancer metastasis and glycolysis through circSSU72-mediated ubiquitination of gamma-catenin and beta-catenin signaling
Zeyu Zhang, Duntao Su, Xiangyuan Qiu, Lei Yao, Fada Xia, Xinying Li

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
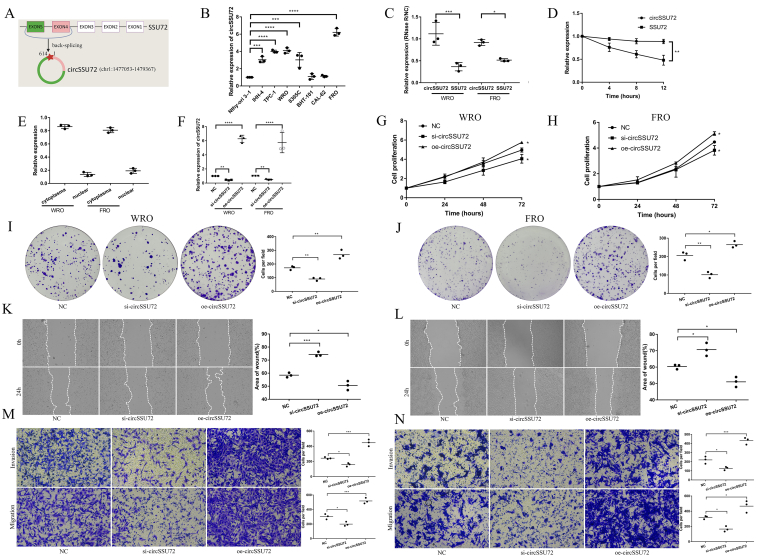 Figure 1
Figure 1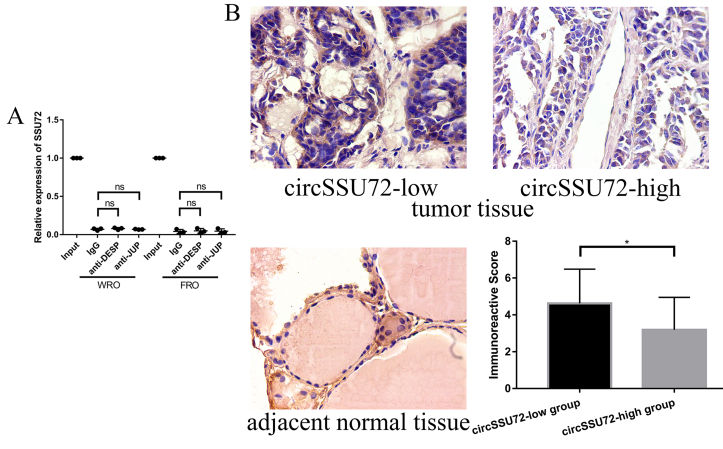 Figure 2
Figure 2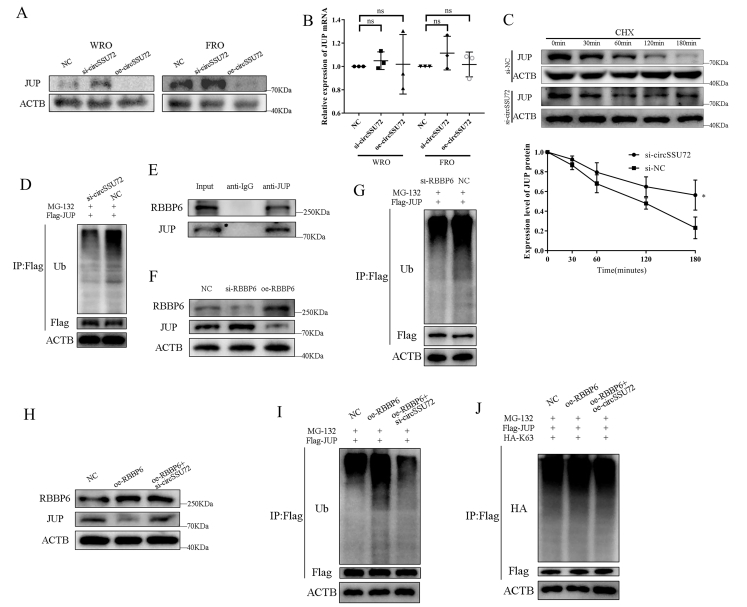 Figure 3
Figure 3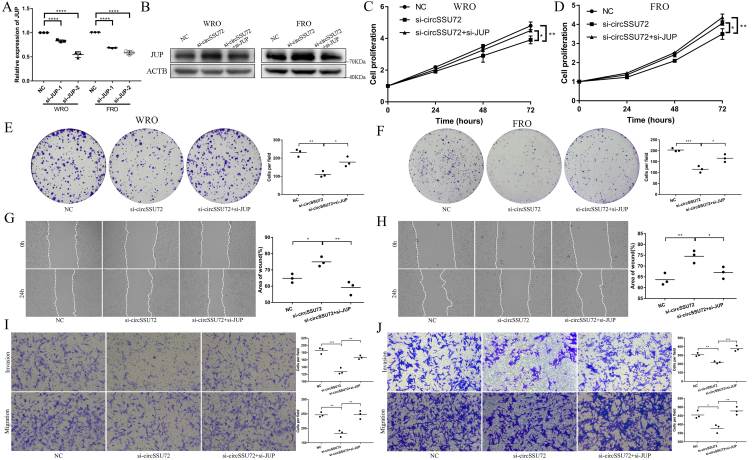 Figure 4
Figure 4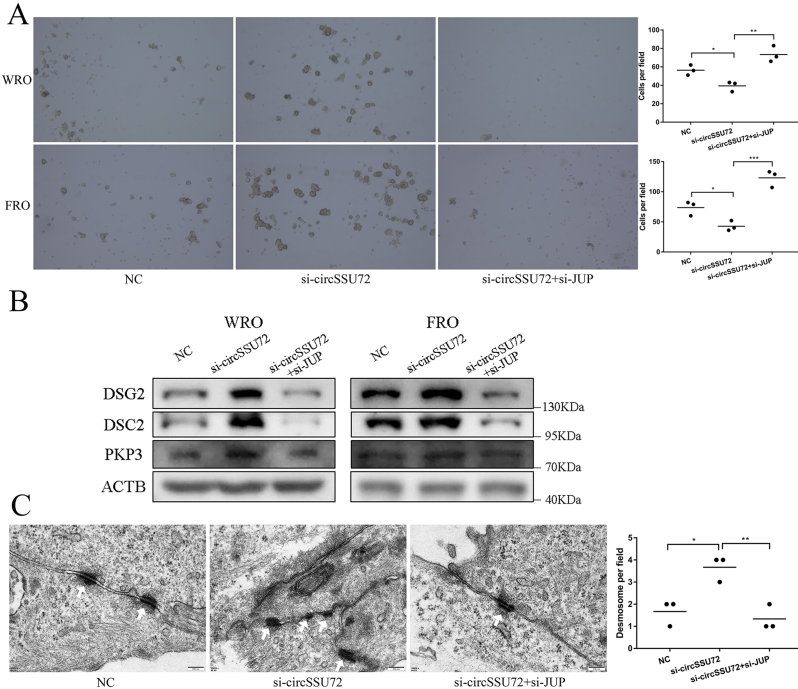 Figure 5
Figure 5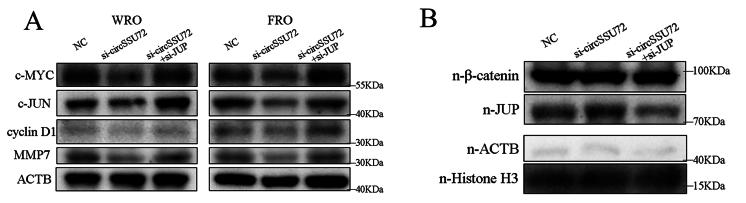 Figure 6
Figure 6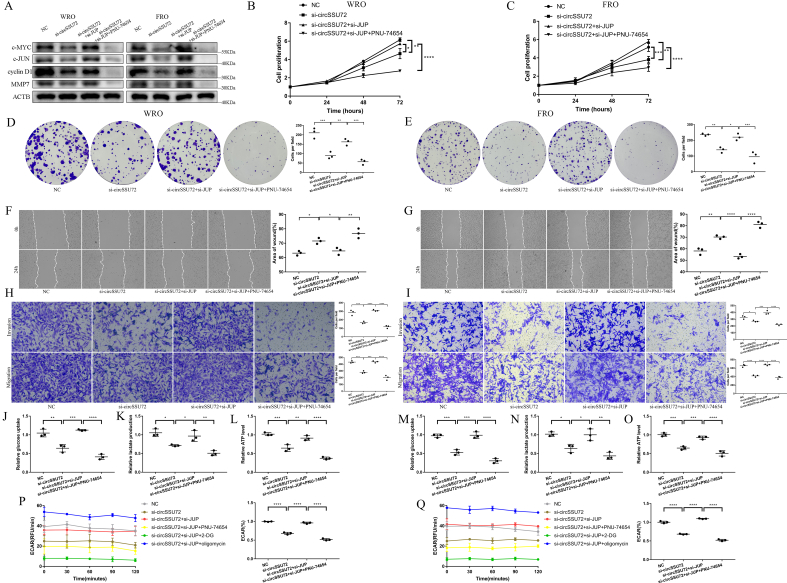 Figure 7
Figure 7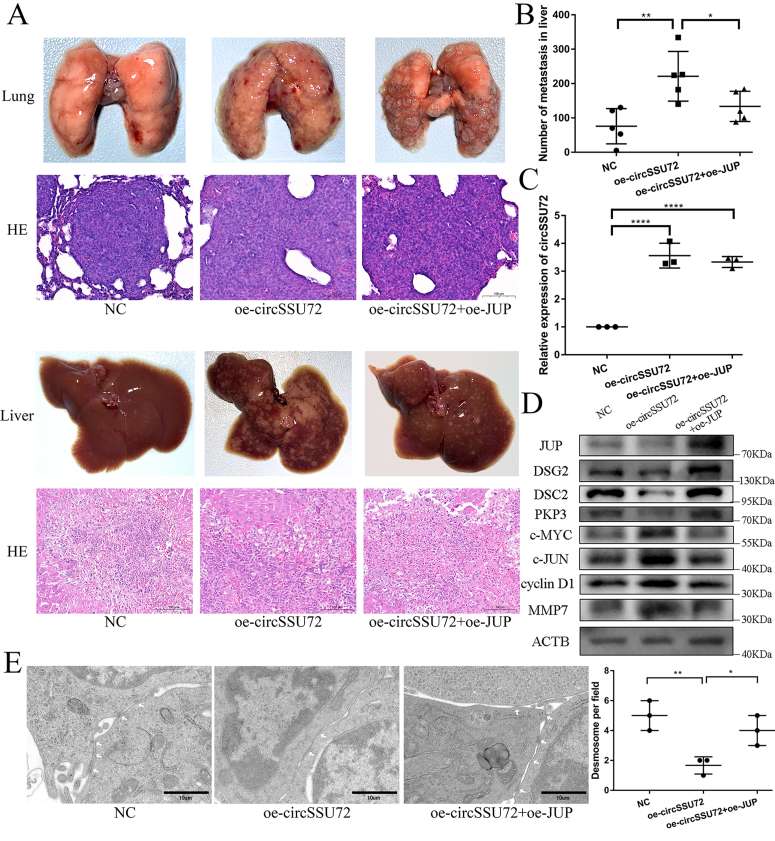 Figure 8
Figure 8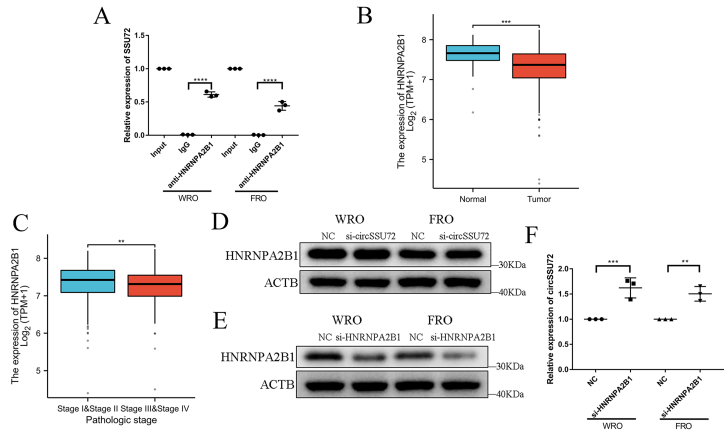 Figure 9
Figure 9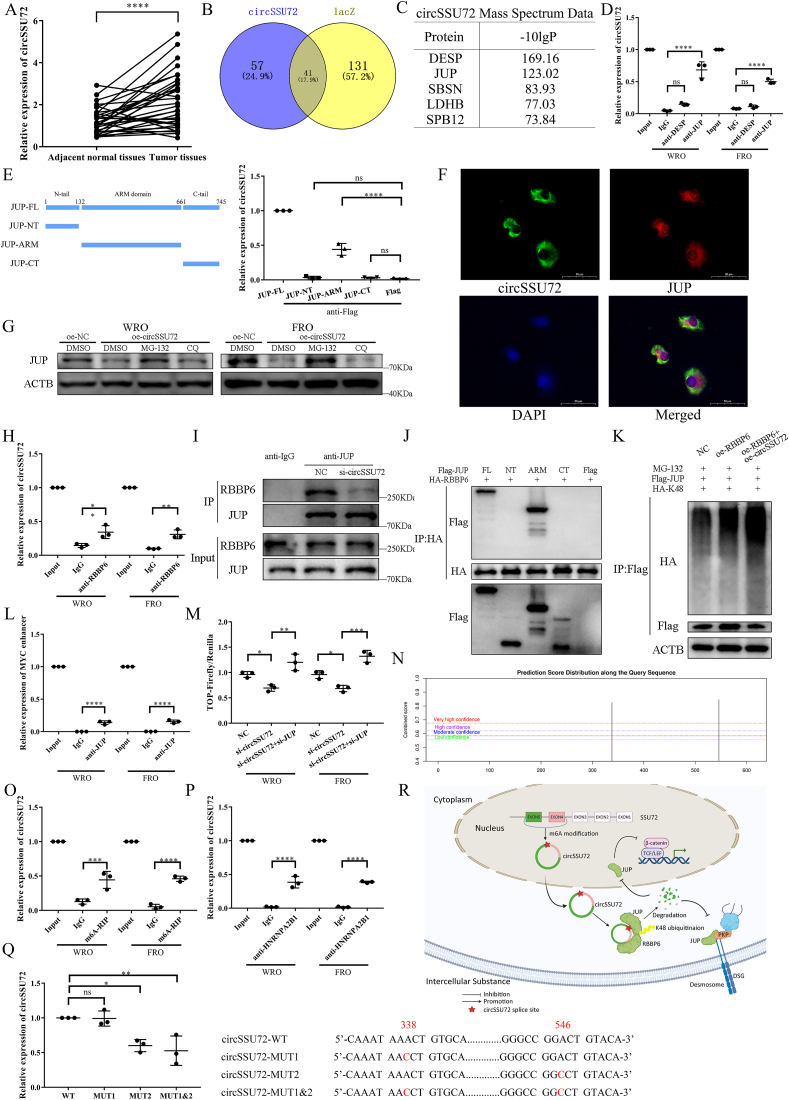 Figure 10
Figure 10Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCircular RNAs in diseases · Histone Deacetylase Inhibitors Research · Cancer-related gene regulation
The incidence of thyroid cancer (TC) has continuously risen worldwide in the past three decades. While most TC patients have a good prognosis, 60% of them still suffer from lymph node metastasis, which is significantly associated with patient prognosis.1 Circular RNAs (circRNAs) are a large class of noncoding RNAs that function as tumor suppressors or tumor promoters in multiple human cancers, including TC.2 In the previous study, we found one circRNA (circSSU72) was significantly up-regulated in both tissues and cell lines of papillary thyroid carcinoma.3 However, the biological role and associated mechanisms of circSSU72 in TC, especially in the field of metastasis, have not been well elucidated.
In the present study, our study demonstrated that circSSU72 could bind to the armadillo (ARM) domain of junction plakoglobin (JUP, also known as gamma-catenin) and promote K48-specific ubiquitination by enhancing the binding between JUP and retinoblastoma-binding protein 6 (RBBP6), influencing desmosomes and the beta-catenin pathway and thus promoting TC metastasis and glycolysis. m^6^A modification at position 546 might involve the up-regulated expression of circSSU72. These findings highlighted the circSSU72/JUP axis as a potential diagnostic marker and a therapeutic target in TC. This study was approved by the Institutional Ethics Committee of Xiangya Hospital, Central South University (No. 201908226), and all subjects signed the written consent.
circSSU72 (hsa_circ_0009294) was derived from two exons of the SSU72 gene, containing 614 nucleotides (Fig. S1A). First, we reconfirmed circSSU72 level was significantly up-regulated in TC (Fig. 1A), which was consistent with TC cell lines (Fig. S1B), and associated with worse tumor characteristics (Table S1). Compared with linear SSU72 mRNA, circSSU72 showed significant insensitivity to RNase R and actinomycin D (Fig. S1C, D). Nuclear and cytoplasmic RNA separation (Fig. S1E) showed that circSSU72 was mainly localized in the cytoplasm. After the effective knockdown and overexpression of circSSU72 (Fig. S1F), the CCK-8 assays, colony formation assays, wound healing assays, and Transwell assays showed that circSSU72 could significantly promote TC cell proliferation, migration, and invasion (Fig. S1G–N).Figure 1N6-methyladenosine-mediated circSSU72 promotes metastasis and glycolysis in thyroid cancer by facilitating RBBP6-induced ubiquitination of JUP. (A) The expression of circSSU72 in thyroid cancer tissues (n = 32). (B, C) Venn diagram and proteins identified with the chromatin isolation by RNA purification and mass spectrometry. (D) The interaction between JUP and circSSU72 was reconfirmed by RNA immunoprecipitation assays (n = 3). (E) RNA immunoprecipitation assay showed that circSSU72 interacted with the ARM domain of JUP in HEK293T cells (n = 3). (F) Fluorescence in situ hybridization and immunofluorescence assays showed circSSU72 and JUP were co-localized in the cytoplasm. (G) The circSSU72-induced degradation of JUP could be rescued by 15 μM MG-132 (a proteasome inhibitor) but not by 20 μM chloroquine (a lysosome inhibitor). (H) The interaction between RBBP6 and circSSU72 was reconfirmed by RNA immunoprecipitation assay (n = 3). (I) Knockdown of circSSU72 reduced the binding between RBBP6 and JUP. (J) RBBP6 interacted with the ARM domain of JUP in HEK293T cells. (K) circSSU72 affected K48-specific ubiquitination of JUP in HEK293T cells. (L) Chromatin immunoprecipitation assays showed that JUP could bind to the MYC enhancer (a typical binding site for the TCF/LEF transcription factor). (M) The TOP-luciferase assay showed that the circSSU72/JUP axis could influence the transcription of TCF/LEF-DNA complexes. (N) The m^6^A modification site of circSSU72 was predicted by the SRAMP database. (O) Methylated RNA immunoprecipitation showed that circSSU72 was associated with m^6^A modification (n = 3). (P) RNA immunoprecipitation reconfirmed the interaction between circSSU72 and HNRNPA2B1 (n = 3). (Q) RNA immunoprecipitation of HNRNPA2B1 in HEK293T cells showed that position 546 (GGACT) was an essential motif in m^6^A modification of circSSU72 (n = 3). (R) Graphical illustration of the regulatory mechanisms of the circSSU72/JUP axis in thyroid cancer. One-way analysis of variance (ANOVA) and Welch's ANOVA were applied for quantitative analyses. ∗p < 0.05; ∗∗p < 0.01; ∗∗∗p < 0.001; ∗∗∗∗p < 0.0001.Figure 1
Subsequently, we identified 57 proteins that specifically bound to circSSU72 with chromatin isolation by RNA purification and mass spectrometry (Fig. 1B). The two most significant proteins (DESP and JUP) were chosen for further validation, while only the binding between circSSU72 and JUP was reconfirmed (Fig. 1C, D). Meanwhile, there was no significant binding between the linear SSU72 gene and the two proteins (Fig. S2A). Further RNA immunoprecipitation (RIP) assays in circSSU72-overexpressing HEK293T cells showed that circSSU72 interacted with the ARM domain of JUP (Fig. 1E). JUP was down-regulated in TC tissues compared with adjacent normal thyroid tissues, and TC tissues in the circSSU72-high group showed a significantly lower expression of JUP (n = 32; Fig. S2B), while the low JUP level was associated with lymph node metastasis (Table S2). After the knockdown or overexpression of circSSU72 in TC cells, the protein level of JUP changed accordingly, while the mRNA level of JUP did not (Fig. S3A, B), indicating that the regulation of JUP by circSSU72 might occur at the posttranslational level. The fluorescence in situ hybridization and immunofluorescence assays showed circSSU72 and JUP were co-localized in the cytoplasm (Fig. 1F).
We then examined JUP protein stability with cycloheximide, showing that circSSU72 significantly destabilized the JUP protein (Fig. S3C). Moreover, MG-132 partially reversed the regulatory effect of circSSU72 on JUP, while chloroquine did not (Fig. 1G). Meanwhile, the knockdown of circSSU72 could inhibit the ubiquitination of JUP (Fig. S3D). These results indicated that the regulatory effect of circSSU72 on JUP might involve the ubiquitin-proteasome system. Among 57 circSSU72-specific binding proteins, we noticed an E3 ubiquitin-protein ligase, RBBP6. Thus, we assumed that circSSU72 might play a role in the binding between RBBP6 and JUP. RIP assay confirmed the binding between circSSU72 and RBBP6 (Fig. 1H), while co-immunoprecipitation assays showed that RBBP6 could bind to JUP (Fig. S3E). Meanwhile, the knockdown of RBBP6 increased the expression of JUP by inhibiting JUP ubiquitination (Fig. S3F, G). Further, co-immunoprecipitation assays showed that the knockdown of circSSU72 could reduce the binding between RBBP6 and JUP (Fig. 1I). Consistently, the knockdown of circSSU72 rescued the RBBP6-induced ubiquitination of JUP (Fig. S3H, I). Further investigations showed that RBBP6 interacted with the ARM domain of JUP (Fig. 1J). To investigate the specific ubiquitination type of JUP, K48 and K63 ubiquitin were transfected with Flag-JUP in HEK293T cells. The results showed that circSSU72 affected the K48-specific ubiquitination of JUP (Fig. 1K; Fig. S3J). These results suggested that circSSU72 promoted the binding between RBBP6 and the ARM domain of JUP, thus affecting the K48-specific ubiquitination of JUP.
After the knockdown of JUP (Fig. S4A, B), CCK-8 assays, colony formation assays, wound healing assays, and Transwell assays revealed that the knockdown of JUP could significantly rescue the effects of circSSU72 knockdown on TC cell proliferation, migration, and invasion (Figs. S4C–J). JUP is reported to play an essential role in desmosomes.4 Thus, we investigated the role of the circSSU72/JUP axis in desmosomes. A hanging-drop assay, desmosome-related protein level, and transmission electron microscopy showed that regulation of circSSU72 significantly affected the cell–cell adhesion and desmosomes, while regulation of JUP significantly rescued these effects (Fig. S5A–C), indicating a role of circSSU72/JUP axis in desmosome structure of TC cells.
Besides, we also found the circSSU72/JUP axis functioned via the beta-catenin signaling pathway (Fig. S6A). Interestingly, while the expression of nuclear JUP was regulated accordingly, the expression of nuclear beta-catenin remained unchanged (Fig. S6B). Subsequent chromatin immunoprecipitation assays showed that JUP could bind to the MYC enhancer, a typical binding site for the TCF/LEF transcription factor (Fig. 1L), indicating binding between JUP and TCF/LEF-DNA complexes. Moreover, the TOP-luciferase assay showed that the circSSU72/JUP axis could influence the transcription of TCF/LEF-DNA complexes (Fig. 1M). PNU-74654, a known beta-catenin signaling pathway inhibitor, was applied to confirm the effect of the circSSU72/JUP axis on the pathway (Fig. S7A). The CCK-8 assays, colony formation assays, wound healing assays, and Transwell assays (Figs. S7B–I) revealed that the effects of the circSSU72/JUP axis on TC cell proliferation, migration, and invasion could be inhibited by PNU-74654. In the pathway analysis of circSSU72-specific binding proteins, we noticed the involvement of glucose metabolism (Supplement Material 1), in which the Wnt/beta-catenin signaling pathway is reported to be involved.5 The knockdown of circSSU72 led to a significantly decreased level of glucose uptake, lactate production, ATP level, and extracellular acidification rate, while the knockdown of JUP could significantly rescue these effects (Fig. S7J–Q).
To validate the effect of the circSSU72/JUP axis on TC metastasis, we established metastatic models in vivo. The results showed that circSSU72 could significantly promote TC cell metastasis, which was reversed by JUP (Fig. S8A–C). Moreover, the Western blot results and transmission electron microscopy confirmed the effects of the circSSU72/JUP axis on desmosomes and the beta-catenin signaling pathway (Fig. S8D, E).
An additional endeavor was made to investigate the reasons for the dysregulated expression of circSSU72 in TC. In all 98 circSSU72-binding proteins, we noticed HNRNPA2B1, an m^6^A reader, indicating that m^6^A modification might be involved. The SRAMP database predicted that there were two potential sites of m^6^A modification in circSSU72 with very high confidence (Fig. 1N). Methylated RIP assays confirmed that circSSU72 was involved in m^6^A modification (Fig. 1O). Further, RIP assays showed that there was binding between circSSU72 and HNRNPA2B1 (Fig. 1P). Interestingly, we also found binding between linear SSU72 mRNA and HNRNPA2B1 (Fig. S9A). Data on thyroid cancer from the TCGA database showed that HNRNPA2B1 was significantly down-regulated and associated with a better tumor stage (Fig. S9B, C). The knockdown of circSSU72 showed no effect on the HNRNPA2B1 protein level (Fig. S9D). However, the knockdown of HNRNPA2B1 significantly increased the circSSU72 level (Fig. S9E, F). To investigate the specific m^6^A motif, we created three mutant plasmids according to the predicted m^6^A site. RIP assays showed that position 546 (GGACT) was an essential motif (Fig. 1Q). These results indicated that m^6^A modification involved the up-regulated level of circSSU72 in TC (Fig. 1R).
Ethics declaration
This study was approved by the Institutional Ethics Committee of Xiangya Hospital, Central South University (No. 201908226), and all subjects signed the written consent. This study strictly complied with the Declaration of Helsinki and its later amendments or comparable ethical standards.
Funding
This work was supported by the 10.13039/501100001809National Natural Science Foundation of China (No. 82073262).
CRediT authorship contribution statement
Zeyu Zhang: Conceptualization, Data curation, Formal analysis, Methodology, Project administration, Visualization, Writing – original draft. Duntao Su: Formal analysis, Validation, Writing – review & editing. Xiangyuan Qiu: Formal analysis, Validation, Writing – original draft. Lei Yao: Methodology, Writing – review & editing. Fada Xia: Investigation, Methodology, Software, Writing – original draft. Xinying Li: Conceptualization, Funding acquisition, Supervision, Writing – review & editing.
Data availability
The datasets supporting the conclusions of this article are included within the article and its additional files.
Conflict of interests
The authors declared no competing interests.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Kim D.H.Kim G.J.Kim S.W.Hwang S.H.Predictive value of ipsilateral central lymph node metastasis for contralateral central lymph node metastasis in patients with thyroid cancer: systematic review and meta-analysis Head Neck 43102021317731843412479110.1002/hed.26787 · doi ↗ · pubmed ↗
- 2Su M.Xiao Y.Ma J.Circular RN As in cancer: emerging functions in hallmarks, stemness, resistance and roles as potential biomarkers Mol Cancer 1812019903099990910.1186/s 12943-019-1002-6PMC 6471953 · doi ↗ · pubmed ↗
- 3Zhang Z.Xia F.Yao L.Jiang B.Li X.circ SSU 72 promotes cell proliferation, migration and invasion of papillary thyroid carcinoma cells by targeting mi R-451a/S 1PR 2 axis Front Cell Dev Biol 10202281702810.3389/fcell.2022.817028 PMC 896713135372340 · doi ↗ · pubmed ↗
- 4Aktary Z.Alaee M.Pasdar M.Beyond cell-cell adhesion: plakoglobin and the regulation of tumorigenesis and metastasis Oncotarget 819201732270322912841675910.18632/oncotarget.15650 PMC 5458283 · doi ↗ · pubmed ↗
- 5Vallée A.Lecarpentier Y.Vallée J.N.The key role of the WNT/β-catenin pathway in metabolic reprogramming in cancers under normoxic conditions Cancers 1321202155573477171810.3390/cancers 13215557 PMC 8582658 · doi ↗ · pubmed ↗