Improvements of Paraquat Treatment in Liquid Media for Behavior and Neurodegenerative Tests
Axel J. Ufarry Alvarado, Malik A. Zaidi Pons, Jayvier Plaza Hernández, Ceidy Torres Ortiz

TL;DR
This study shows that using Paraquat in liquid media improves testing for motor and neurodegenerative effects in worms linked to ALS.
Contribution
A more cost-effective and rapid method for behavior and neurodegenerative assays in worms using Paraquat in liquid media.
Findings
2.5 mM Paraquat in liquid M9 buffer increased locomotion defects and neurodegeneration in C. elegans.
Paraquat treatment in liquid media is more rapid and reproducible than solid NGM media for these assays.
Abstract
Amyotrophic Lateral Sclerosis (ALS) is a disease characterized by misfolded and aggregated proteins that have toxic effects on motor neurons. The missense mutation, G85R, of the sod-1 gene associated with ALS displays locomotor impairments in Caenorhadbitis elegans ( C. elegans ). We treated the sod-1 (G85R) strain with 0 and 2.5 mM Paraquat treatments in a liquid M9 buffer for 4 hours and in solid NGM media for 18 hours. In both methodologies, the locomotion defects and neurodegeneration were significantly increased with 2.5 mM Paraquat. Our work provides evidence of methodology that is more cost effective, rapid and reproducible to perform behavior and neurodegenerative assay in worms.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
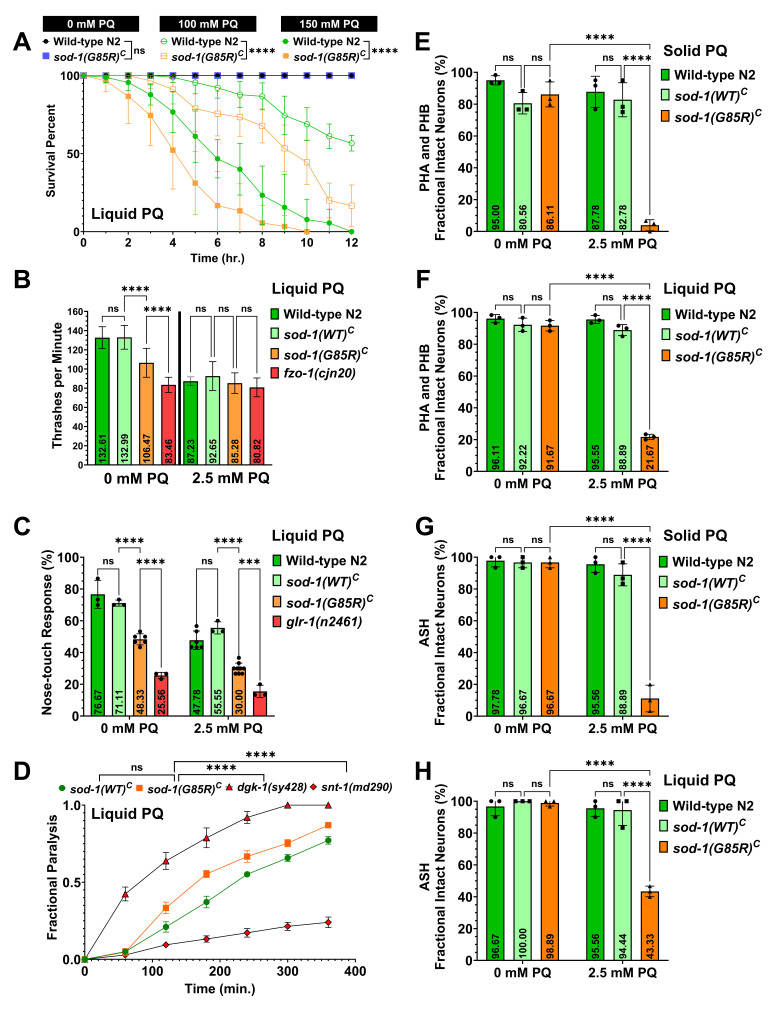 Figure 1
Figure 1|
|
|
|
|
|
|
CGC |
|
|
|
CGC |
|
|
|
Dr. Hart Lab, Bio Med Center Brown University |
|
|
|
CGC |
|
|
|
CGC |
|
|
|
CGC |
|
|
|
CGC |
|
|
|
Dr. Colon Ramos Lab in Yale University |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsParaquat toxicity studies and treatments
Description
Amyotrophic Lateral Sclerosis (ALS) is a motor neuron disease characterized by the loss of cholinergic and glutaminergic neurons. Although the cause of ALS is unknown, the Sod1 gene was one of the first genes related to the disease (Rosen et al., 1993; Nguyen, 2024). The Sod1 gene codifies for Cu/Zn superoxide dismutase 1 ( SOD-1 protein, which is an antioxidant enzyme catalyzing the conversion of superoxide ions (O 2 ^-^ ) to hydrogen peroxide (H 2 O 2 ). Hundreds of missense mutations have been identified in the SOD1 gene that are associated with ALS (Mathis et al., 2019; Huai & Zhang, 2019 and Abel et al., 2012). Some SOD1 mutations have a loss and toxic gain of function that contributes to ALS progression (Qualls et al., 2013; Sahin et al., 2017; Baskoylu et al ., 2018).
C. elegans * has served as a model for ALS, with * sod-1 * mutations conferring sensitivity to oxidative stress including Paraquat treatment (Osborne et al., 2021; Wang et al., 2009). A previous study by Baskoylu et al. (2018) reported that * C. elegans * single copy/knock-in model for G85R was associated with cholinergic and glutamatergic neurodegeneration after oxidative stress. In the current study, we asked if similar results could be observed when experiments are conducted in liquid media (M9 buffer) instead of solid media (nematode growth medium, NGM).
To this extent, we used the * sod-1 (G85R) ^C ^ * strain, as well as the * sod-1 (WT) ^C ^ * and N2 wild type strains. The * sod-1 (WT) ^C ^ * strain contains all the silent codon changes needed for CRISPR/Cas9 genome editing of the * sod-1 (G85R) ^C^ * strain (Baskoylu et al., 2018). First, we tested the nematode survival rate under PQ-induced oxidative stress. The * sod-1 (G85R) ^C ^ * strain and N2 wild type were exposed to 0, 100 and 150 mM PQ for 12 hours. Results showed (Fig A) , that at the highest PQ concentration, the * sod-1 (G85R) ^C^ * mutants survival rate had decreased survival compared to wild type. The liquid media by itself did not have an effect on survival. Then we tested if the locomotion alterations associated with cholinergic and glutamatergic defects in * sod-1 (G85R) ^C^ * strain could be replicated in liquid media. The nose touch avoidance assay was used to evaluate the response of glutamatergic neurons such as ASH (Davis et al., 2022) and the thrashing assay to evaluate locomotion mediated by cholinergic motor neurons and muscle cells. The results showed a decrease in thrashing in the absence of PQ in the * sod-1 (G85R) ^C^ * strain when compared to controls strains, and a defective nose touch avoidance response with or without PQ-induced oxidative stress (Fig B & C) . Even though the * sod-1 (G85R) ^C^ * strain showed locomotion defects, no changes in synaptic transmission of cholinergic neurons were observed (Fig D) . Nevertheless, the aldicarb resistance assay showed accelerated paralysis in * sod-1 (G85R) ^C ^ * strain when compared to wild type, with a critical point at 180 minutes, indicating defective neuromuscular signaling. Since defects on nose touch avoidance response assay may be associated with glutamatergic neurodegeneration, we compared the uptake of a lipophilic fluorescent dye under PQ treatment in solid vs. liquid media. The * sod-1 (G85R) ^C^ * strain showed a decrease in the percentage of intact ASH, PHA and PHB neurons in both media (Fig E-H) . However, these effects were observed earlier during PQ exposure in liquid media (4 hr, Fig F & H ) compared to solid media (18 hr, Fig E & G ).
Overall, we were able to consistently reproduce the results observed in the * sod-1 (G85R) ^C^ * single copy/knock-in C. elegans model developed by Baskoylu et al. (2018) regardless of whether treatment is done on solid or liquid media. Therefore, we conclude that it is possible to conduct studies of neurodegeneration and behavior in * C. elegans * using liquid media, which allows it to be conducted in less time and at lower cost.
Methods
**Worm maintenance: ** The Nematode Growth Medium (NGM) [17 g agar, 3 g NaCl, 2.5 g peptone, 1 ml 1M CaCl 2 , 1 ml 5 mg/ml cholesterol in ethanol, 1 ml 1 M MgSO 4 , and 25 ml of 1M KPO 4 ] plates were seeded * with Escherichia coli * OP50 and incubated to 37°C overnight and stored at room temperature in boxes or used for cultivating the worms. The worms were transferred every 3-4 days to seeded plates with * Escherichia coli * OP50 . All worm strains were maintained in an incubator at 20°C.
Worm synchronization: Young adult worms were transferred to NGM plates and incubated at 20°C until the next day. After removing the young adult worms, the eggs remain on the plate incubated for four days at 20°C to let the worms reach day 1 of adulthood. For all the experiments the worms were synchronized to the first day of adulthood.
**Paraquat treatment in liquid and solid media: ** The worms were transferred to a well (24-well plate) or microtubes with M9 buffer. Then, the treatments were added to obtain a final concentration Paraquat in M9 buffer (refers to PQ Liquid) for 4 hours. After incubation worms were transferred to solid plates with E.coli OP50 for 30 minutes before experimental assay. For solid PQ, worms were transferred to NGM plates seeded with E. coli OP50 with 0 mM and 2.5 mM Paraquat (refers to PQ solid) for 18 hours. After incubation worms were transferred to solid plates only with E.coli OP50 for 30 minutes before experimental assay. All experiments were conducted blinded for genotype and treatment.
**Survival test: ** The worms were transferred to a well (24-well plate) with 100 or 150 mM Paraquat treatment diluted in M9 buffer (refers to PQ liquid). The worm was counted dead if no movement occurred after touching the worms 3 times in the head and tail consecutively with a platinum wire. The worms were evaluated each hour for 12 hours. The survival percentage was calculated and graphed with the standard deviation of the three independent experiments (N=3).
**Nose Touch Avoidance Assay: ** The evaluation consists of placing a single brush bristle perpendicular to the front of a worm moving forward, then waiting for a nose collision. If the worm responds with reverse locomotion, it is classified as non-defective. The nose touch avoidance percentage was calculated of the total non-defective worms divided by the total number of worms evaluated multiplied by 100. The nose touch avoidance percentage was calculated and graphed with the standard deviation of the three independent experiments (N=3).
**Thrashing assay: ** The worms were transferred to a well with 50 µL of M9 buffer in a 96-well plate. The worms were left for 30 seconds to acclimatize in the well. A three-minute video of the worms swimming was taken with a NIKON SMZ745T stereoscope with Kopa Software. The evaluation consists of processing the video in slow motion and counting the number of thrashes (C-shape) per minute. An average of three minutes was calculated for each worm. The number of thrashes was calculated and graphed with the standard deviation of the three independent experiments (N=3).
**Aldicarb Resistance Assay: ** The worms were transferred to a microtube with M9 buffer for 4 hours at 20°C. The worms were transferred to the NGM plates with 1 mM aldicarb seeded with E. coli OP50 and evaluated for worm paralysis each hour for 6 hours. Paralysis was defined as the absence of movement and pumping for 5 seconds after touching the worm head and tail 3 times consecutively. The fractional paralysis was calculated and graphed with the standard deviation of the three independent experiments (N=3).
DiI staining assay: After treatment, the worms were washed and incubated with ('DiI'; DiIC 18 (3)) D282 for 2 hours. All worms were transferred to NGM plates seeded with E. coli OP50 for 30 min. Up to 15-20 worms placed in microscope slides were immobilized with 10 mM levamisole. The ASH in the head and the PHA and PHB in the tail were visualized at 575-625 nm and scored. The percentage of fractional intact neurons was calculated and analyzed in the three independent experiments (N=3).
Statistical analysis
Quantitative data for the statistical analysis is organized and processed using the Microsoft Office 2011 Excel software package (Microsoft Corporation, Redmond, USA) and analyzed using GraphPad Prism version 10.0.0 for Windows (GraphPad Software, Boston, Massachusetts, USA). The standard deviation is used to present the maximum and minimum values with the meaning in graphs. The value of p < 0.05 is considered statistically significant. For nose touch avoidance and neurodegeneration assays a Two-way ANOVA was used for statistical analysis. For thrashing assay, a One-way ANOVA was used for statistical analysis. For survival and aldicarb resistance assays a log rank test is used to compare distribution of time of curves and determine significant differences.
Reagents
Worm and bacteria strains
The following strains were obtained from the Caenorhabditis Genetics Center (CGC) funded by NIH Office of Research Infrastructure Programs (P40 OD010440)
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abel O Powell JF Andersen PM Al-Chalabi A 2012716 AL So D: A user-friendly online bioinformatics tool for amyotrophic lateral sclerosis genetics.Hum Mutat 3391059-77941345135110.1002/humu.2215722753137 · doi ↗ · pubmed ↗
- 2Baskoylu SN Yersak J O'Hern P Grosser S Simon J Kim S Schuch K Dimitriadi M Yanagi KS Lins J Hart AC 2018108 Single copy/knock-in models of ALS SOD 1 in C. elegans suggest loss and gain of function have different contributions to cholinergic and glutamatergic neurodegeneration.P Lo S Genet 14101553-7390 e 1007682 e 100768210.1371/journal.pgen.100768230296255 PMC 6200258 · doi ↗ · pubmed ↗
- 3Davis P Zarowiecki M Arnaboldi V Becerra A Cain S Chan J Chen WJ Cho J da Veiga Beltrame E Diamantakis S Gao S Grigoriadis D Grove CA Harris TW Kishore R Le T Lee RYN Luypaert M Müller HM Nakamura C Nuin P Paulini M Quinton-Tulloch M Raciti D Rodgers FH Russell M Schindelman G Singh A Stickland T Van Auken K Wang Q Williams G Wright AJ Yook K Berriman M Howe KL Schedl T Stein L Sternberg PW 202244 Worm Base in 2022-data, processes, and tools for analyzing Caenorhabditis elegans.Genetics 22040016-673110.1093/genetics/iyac 00335134929 PMC 89 · doi ↗ · pubmed ↗
- 4Huai J Zhang Z 2019521 Structural Properties and Interaction Partners of Familial ALS-Associated SOD 1 Mutants.Front Neurol 101664-229552752710.3389/fneur.2019.0052731164862 PMC 6536575 · doi ↗ · pubmed ↗
- 5Mathis S Goizet C Soulages A Vallat JM Masson GL 2019221 Genetics of amyotrophic lateral sclerosis: A review.J Neurol Sci 3990022-510X 21722610.1016/j.jns.2019.02.03030870681 · doi ↗ · pubmed ↗
- 6Nguyen Lien 2024521 Updates on Disease Mechanisms and Therapeutics for Amyotrophic Lateral Sclerosis Cells 13112073-440988888810.3390/cells 1311088838891021 PMC 11172142 · doi ↗ · pubmed ↗
- 7Osborne JF Yanagi KS Hart AC 2021115 Genetic interactions in a C. elegans sod-1 ALS model: glutamatergic neuron degeneration.Micro Publ Biol 202110.17912/micropub.biology.00033833474528 PMC 7812382 · doi ↗ · pubmed ↗
- 8Qualls DA Prudencio M Roberts BL Crosby K Brown H Borchelt DR 20131217 Features of wild-type human SOD 1 limit interactions with misfolded aggregates of mouse G 86R Sod 1.Mol Neurodegener 8464610.1186/1750-1326-8-4624341866 PMC 3881023 · doi ↗ · pubmed ↗