A knock-in translational reporter for NLP-29 reveals AMP secretion to the apical extracellular matrices following epidermal damage in Caenorhabditis elegans
Nathalie Pujol, Henrik Bringmann

TL;DR
Scientists created a tool to track a specific antimicrobial peptide in worms, showing it is released after skin damage.
Contribution
A knock-in translational reporter for NLP-29 enables visualization of AMP secretion in C. elegans.
Findings
NLP-29 is secreted into the cuticle after genetic or physical damage in C. elegans.
The NLP-29::mKate2 reporter allows visualization of AMP trafficking and secretion.
This tool will help dissect the roles of NLP-29 in immunity and signaling.
Abstract
Antimicrobial peptides (AMPs) are small proteins produced and secreted as part of the innate immune response to infection and wounding. They target pathogens and can also function as signalling molecules, for example, promoting sleep in response to injury in C. elegans . A transcriptional reporter transgene for nlp-29 has been pivotal in studying AMP gene expression and regulation, but to understand AMPs antimicrobial and signalling roles, protein expression and trafficking needs to be monitored. We have now created a knock-in translational reporter allele for nlp-29 , with NLP-29 fused to mKate2, that enables visualisation of this secreted AMP. Using the NLP-29::mKate2 reporter, we demonstrate that NLP-29 is secreted into the cuticle upon genetic or physical cuticle damage. NLP-29::mKate2 will therefore be a valuable tool for visualising the secretion of this peptide in C.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
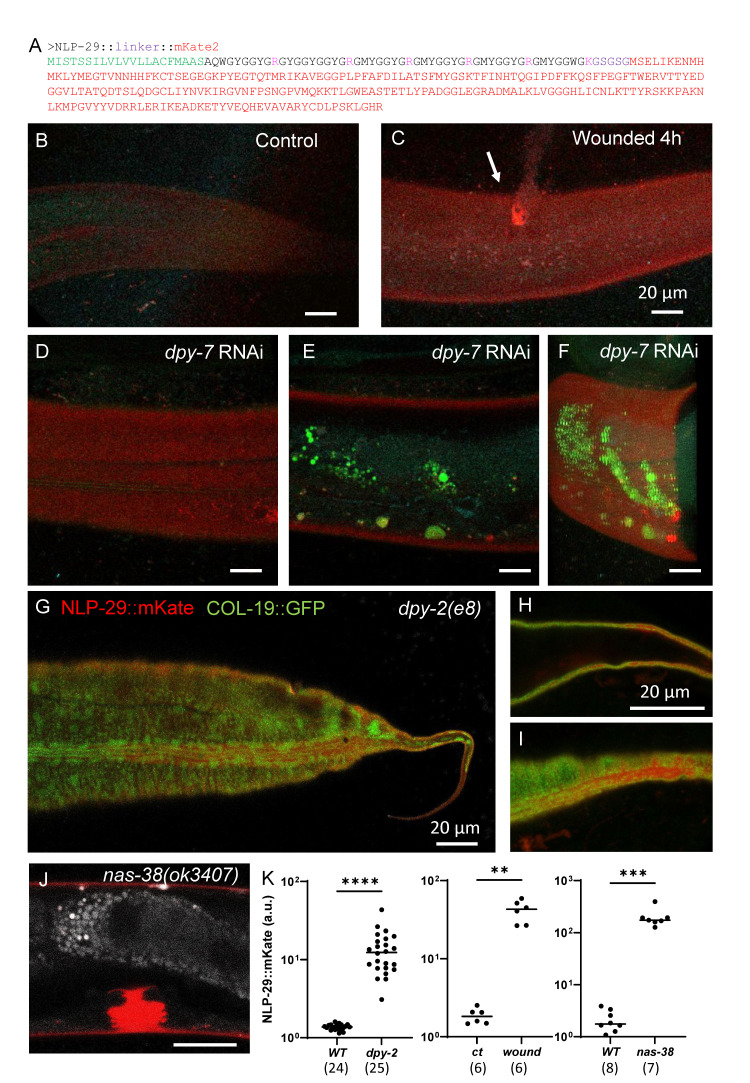 Figure 1
Figure 1- —French National Research Agency
- —Deutsche Forschungsgemeinschaft (Germany)https://ror.org/018mejw64
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
Topicsbioluminescence and chemiluminescence research
Description
The skin serves as the outermost defence against pathogens. Across species, damage or infection of the skin triggers an innate immune response in the underlying epidermis, involving a cascade of signalling molecules, leading to the production of antimicrobial peptides (AMPs) (Hanson and Lemaitre, 2020; Lai and Gallo, 2009; Martineau et al., 2021). These AMPs act against pathogens by disrupting membrane structures (Lai and Gallo, 2009), and also play roles in signalling, such as promoting sleep in response to injury (Sinner et al., 2020; Toda et al., 2019) and neuron degeneration (E et al., 2018).
C. elegans * is an important model for AMP research due to its lack of an adaptive immune system, its amenability to genetic manipulation, and its ability to be wounded or infected by various pathogens (Couillault et al., 2004; Martineau et al., 2021; Pujol et al., 2008a; Pujol et al., 2008b; Taffoni et al., 2020). Innate immune responses of the epidermis involve the production of two main classes of AMPs: 1) neuropeptide-like peptides (NLP) AMPs, regulated by PMK-1 p38 MAP kinase signalling (Pujol et al., 2008a), and 2) caenacin (CNC) AMPs, regulated by the TGF-β homolog DBL-1 (Martineau et al., 2021; Zugasti and Ewbank, 2009). The mature peptides are basic and rich in glycine and aromatic amino acids and harbour a conserved motif QWGYG just C-terminal to the predicted signal sequence cleavage site (Couillault et al., 2004). Transcriptional reporters based on multi-copy arrays have been instrumental for visualising AMP expression and identification of the signalling pathways leading to their regulation, like * frIs7 (nlp-29p::GFP) * (Pujol et al., 2008a), and * cnc-2 p::GFP * (Zugasti and Ewbank, 2009), as reviewed in (Martineau et al., 2021). While very few antimicrobial proteins have been directly tagged, including LYS-1 (Mallo et al.,2002), SPP-3 and SPP-12 (Hoeckendorf el al., 2012a&b) *, * and ASP-3 and ASP-4 (Wong et al., 2007), AMPs have not yet been visualised in * C. elegans * , and it is not known, for example, whether they are secreted apically or in a basolateral manner.
To investigate AMP secretion, we designed and created knock-in translational fusion alleles of * cnc-2 * and * nlp-29 * with mKate2 . We used a linker sequence (GSGSG) to attach mKate2 to the C-terminus of either AMP gene. We codon-optimized the linker and mKate2 sequence for optimal expression and inserted two synthetic introns into the mKate2 coding sequence (Redemann et al., 2011) ( Figure 1A ; methods). This optimised construct was then integrated into the endogenous * cnc-2 * and * nlp-29 * loci using CRISPR-Cas9 to create * cnc-2 ::mKate2 * and * nlp-29 ::mKate2 * , respectively. The CNC-2 and NLP-29 preproteins contain a cleavable N-terminal signal sequence but lack the dibasic residues that act as sites of internal proteolytic cleavage (Couillault et al., 2004; Nathoo et al., 2001). This suggests that the mature peptides are not further processed and that the AMP would remain connected to the mKate2 sensor after secretion, which should allow for visualising the AMPs in vivo .
We imaged mKate2 fluorescence in adult worms under baseline conditions and after cuticle damage, which triggers innate immune responses involving the upregulation of * cnc-2 * and * nlp-29 * expression (Dodd et al., 2018; Meng et al., 2020; Pujol et al., 2008a). For damaging the cuticle, we used either a glass needle (Pujol et al., 2008a; Xu and Chisholm, 2011) or the inactivation of furrow collagens (Aggad et al., 2023; Dodd et al., 2018; Pujol et al., 2008b; Sundaram and Pujol, 2024; Taffoni et al., 2020). We did not detect fluorescence from * cnc-2 ::mKate2 * under these conditions, potentially because the endogenous level of expression is too low, which is in line with the results obtained with the transcriptional reporter (Zugasti and Ewbank, 2009). * nlp-29 ::mKate2 * also showed no detectable fluorescence under baseline conditions. This is consistent with low level of expression observed in several transcriptomic studies (Couillault et al., 2004; Dodd et al., 2018; Hendriks et al., 2014). A clear signal in the cuticle was, however, observed after epidermal wounding in the wild type ( Figure 1 C) and upon inactivation of the furrow collagen * dpy-7 * using RNAi ( Figure 1 E-K). We confirmed that the NLP-29::mKate2 signal localized to the cuticle using a cuticle collagen marker, COL-19 ::GFP. In the furrow collagen * dpy-2 ( e8 ) * mutant, both signals co-localised ( Figure 1 I-K). We confirmed this localisation in a gain-of-function mutant of * nas-38 * ( * nas-38 ( ok3407 ), * Figure 1J ), which represents another condition in which * nlp-29 * is known to be up-regulated (Sinner et al., 2020). Interestingly, NLP-29 is also found to be secreted in the vulva lumen during the L4 stage in * nas-38 * mutants ( Figure 1J ) or furrow-less mutants. The quantification of the reporter signal in the cuticle of all three conditions — furrow-less * dpy-2 * mutants, gain-of-function * nas-38 * mutants, and wounded wild-type worms — shows a significant increase in the NLP-29::mKate2 fluorescence signal ( Figure 1K ). The localisation of NLP-29::mKate2 in the apical extracellular matrices aligns with its antimicrobial function.
Given the known signalling function of AMPs such as NLP-29 , one would predict that some of the protein should be released basolaterally into the pseudocoelom to then diffuse and act on the nervous system (E et al., 2018; Sinner et al., 2020). We did not, however, observe NLP-29::mKate2 in the pseudocoelom or coelomocytes. This lack of detection of NLP-29::mKate2 internally could reflect a low level basolateral secretion, and/or its dilution or degradation in the pseudocoelom. It should also be noted that any detection in coelomocytes is complicated by their intrinsic red autofluorescence. We cannot, therefore, rule out potential basolateral secretion that might be important for NLP-29 's signalling to neurons (E et al., 2018; Sinner et al., 2020). Our observations suggest that NLP-29 's antimicrobial functions that rely on direct action on pathogen membranes could require higher peptide concentrations compared to signalling functions, which are amplified via G protein-coupled receptors (Sinner et al., 2020; Zugasti et al., 2014). Hence, more sensitive detection methods with brighter fluorophores, or immunohistochemistry, might be needed to visualise the putative internal secretion of NLP-29 or other AMPs in the future.
This NLP-29::mKate2 fusion protein is, to our knowledge, the first reporter allowing an AMP's localisation to be monitored in * C. elegans * and will be a valuable tool for future studies.
Methods
Nematode strains
All * C. elegans * strains were maintained on nematode growth medium (NGM) and fed with E. coli OP50 , as described (Stiernagle, 2006). The TP12 * kaIs12 [ COL-19 ::GFP] * (Thein et al., 2003) and * dpy-2 ( e8 ) * (Brenner, 1974) strains were obtained from the CGC. The IG2117 * nlp-29 ( syb1965 )[NLP-29::linker::mKate2]) V; kaIs12 [ COL-19 ::GFP], * IG2115 * dpy-2 ( e8 ) II; nlp-29 ( syb1965 )[NLP-29::linker::mKate2]) V; kaIs12 [ COL-19 ::GFP] * and IG2000 * nas-38 ( ok3407 ) X; nlp-29 ( syb1965 )[NLP-29::linker::mKate2]) V * were obtained by conventional crosses.
** Generation of reporter knock-in alleles for * cnc-2 * and * nlp-29 * **
We designed the linker and mKate2 sequence in silico, and the corresponding DNA was synthesized by a commercial provider. Using this synthetic DNA as a template, the endogenous loci of * cnc-2 * and * nlp-29 * were edited using CRISPR-Cas9 by a commercial service provider (Sunybiotech) to introduce the gene edits. Synonymous mutations were also introduced during the gene editing process to prevent recutting by the nuclease (synonymous changes for * nlp-29 * are labelled in bold in the sequence below). The allele for * cnc-2 * was constructed using the identical linker::mKate2 sequence, with the sequence GGAATGCT C ATGGGCAAG mutated to GGAATGCT G ATGGGCAAG (a silent mutation) to remove the PAM site. The editing resulted in two strains, both of which were superficially normal and homozygous viable.
PHX1965 * nlp-29 ( syb1965 [::linker::mKate2]) * with the following sequence (the linker is underlined, the synonymous mutation is labelled in bold )
ATGATTTCAACCTCTTCAATTCTTGTTCTTGTCGTCCTTCTCGCCTGCTTCATGGCTGCCAGCGCACAATGGGGATATGGAGGATATGGAAgtgagtttttttgtgctttttgacttatctcaaaaaaagtagatcattcacacatattttcattttcagGAGGATATGGAGGATATGGTGGATACGGACGAGGAATGTATGGAGGCTATGGACGCGGAATGTATGGTGGATATGGACGTGGAATGTACGGAGGATACGGACGCGGAATGTATGGAGG T TGGGGAAAG GGATCCGGATCCGGA ATGTCCGAGCTCATCAAGGAGAACATGCACATGAAGCTCTACATGGAGGGAACCGTCAACAACCACCACTTCAAGTGCACCTCCGAGGGAGAGGGAAAGCCATACGAGGGAACCCAAACCATGCGTATCAAGgtaagtttaaacatatatatactaactaaccctgattatttaaattttcagGCCGTCGAGGGAGGACCACTCCCATTCGCCTTCGACATCCTCGCCACCTCCTTCATGTACGGATCCAAGACCTTCATCAACCACACCCAAGGAATCCCAGACTTCTTCAAGCAATCCTTCCCAGAGGGATTCACCTGGGAGCGTGTCACCACCTACGAGGACGGAGGAGTCCTCACCGCCACCCAAGACACCTCCCTCCAAGACGGATGCCTCATCTACAACGTCAAGATCCGTGGAGTCAACTTCCCATCCAACGGACCAGTCATGCAAAAGAAGACCCTCGGATGGGAGGCCTCCACCGAGACCCTCTACCCAGCCGACGGAGGACTCGAGGGACGTGCCGACATGGCCCTCAAGCTCGTCGGAGGAGGACACCTCATCTGCAACCTCAAGgtaagtttaaacatgattttactaactaactaatctgatttaaattttcagACCACCTACCGTTCCAAGAAGCCAGCCAAGAACCTCAAGATGCCAGGAGTCTACTACGTCGACCGTCGTCTCGAGCGTATCAAGGAGGCCGACAAGGAGACCTACGTCGAGCAACACGAGGTCGCCGTCGCCCGTTACTGCGACCTCCCATCCAAGCTCGGACACCGTTAAatacatggataaccatctattaataatttgaaaatctcatttcgttatgtaacaatgcgttgtacatatcctgatttctcacttttttcttgaataaaaacttgcataat
PHX1939 * cnc-2 ( syb1939 [::linker::mKate2]) * .
The alleles were Sanger sequenced, confirming that the intended edits had been successfully introduced.
The strain PHX1965 is available at the * Caenorhabditis * Genetics Center (CGC). PHX1939 is available upon request.
Confocal microscopy
Worms were mounted on a 2 % agarose pad, in a drop of 1 mM levamisole in 50 mM NaCl. Images were acquired during the following 60 min, using a Zeiss LSM880 confocal laser scanning microscope with a Plan-Apochromat Oil DIC M27 40×/1.4 objective and the acquisition software Zen. Pinhole size was set to 1 AU. Samples were illuminated with 488 nm (GFP) and 561 nm (mKate2) with constant laser power, with 4 lines accumulation and 750 gain settings. Spectral imaging combined with linear unmixing was used to separate the autofluorescence (Aggad et al., 2023). Quantification of the NLP-29::mKate2 red signal was performed using Fiji. The worm cuticle was automatically segmented based on the COL-19 ::GFP green signal using a Gaussian blur filter with a radius of 2 pixels and a triangle threshold to convert the image to binary and create a mask. The resulting mean intensity in the define region of interest (ROI) for each condition was analysed with the GraphPad Prism 10.3 software. Statistical differences between groups were determined by the unpaired nonparametric Mann-Whitney test.
RNA interference
RNAi bacterial clones were obtained from the Ahringer library (Kamath et al., 2003) and verified by sequencing. RNAi bacteria were seeded on NGM plates supplemented with 100 g/ml ampicillin and 1 mM Isopropyl-β-D-thiogalactopyranoside (IPTG). Worms were transferred onto RNAi plates as L1 larvae and cultured at 25 °C until the young adult stage. In all our experiments, we used * sta-1 * as a control, as we have shown that it does not affect the development nor any stress or innate response in the epidermis (Aggad et al., 2023).
Wounding
Needle wounding was performed as previously described (Pujol et al., 2008a; Taffoni et al., 2000) with a standard microinjection needle under a dissecting microscope by pricking the worm's posterior body or tail on agar plates; worms were analysed after 4 to 8 h.
Reagents
PCR and sequencing primers used for * syb1965 * where:
HB20-seq-s:TGTTCTTGTCGTCCTTCTCG
HB20-seq-a:CCATGTCTCAGTTGCCTTA
PCR and sequencing primers used for * syb1939 * where:
HB19-seq-s: CGTCATCATTTGGTTCGTCA
HB19-seq-a: TCCTTTGGTCTCGAAATGAC
HB19-mid-s: CTGGGAGCGTGTCACCACCTA
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Aggad Dina Brouilly Nicolas Omi Shizue Essmann Clara Luise Dehapiot Benoit Savage-Dunn Cathy Richard Fabrice Cazevieille Chantal Politi Kristin A Hall David H Pujol Remy Pujol Nathalie 2023313 Meisosomes, folded membrane microdomains between the apical extracellular matrix and epidermise Life 122050-084X 10.7554/elife.75906 PMC 1001068936913486 · doi ↗ · pubmed ↗
- 2Brenner S 197451 THE GENETICS OF CAENORHABDITIS ELEGANS Genetics 7711943-2631719410.1093/genetics/77.1.714366476 PMC 1213120 · doi ↗ · pubmed ↗
- 3Dodd William Tang Lanlan Lone Jean-Christophe Wimberly Keon Wu Cheng-Wei Consalvo Claudia Wright Joni E Pujol Nathalie Choe Keith P 201841 A Damage Sensor Associated with the Cuticle Coordinates Three Core Environmental Stress Responses in Caenorhabditis elegans Genetics 20841943-26311467148210.1534/genetics.118.30082729487136 PMC 5887142 · doi ↗ · pubmed ↗
- 4E Lezi Zhou Ting Koh Sehwon Chuang Marian Sharma Ruchira Pujol Nathalie Chisholm Andrew D. Eroglu Cagla Matsunami Hiroaki Yan Dong 201811 An Antimicrobial Peptide and Its Neuronal Receptor Regulate Dendrite Degeneration in Aging and Infection Neuron 9710896-6273125138.e 510.1016/j.neuron.2017.12.00129301098 PMC 5757245 · doi ↗ · pubmed ↗
- 5Hanson Mark Austin Lemaitre Bruno 202021 New insights on Drosophila antimicrobial peptide function in host defense and beyond Current Opinion in Immunology 620952-7915223010.1016/j.coi.2019.11.00831835066 · doi ↗ · pubmed ↗
- 6Hendriks Gert-Jan Gaidatzis Dimos Aeschimann Florian Großhans Helge 201421 Extensive Oscillatory Gene Expression during C. elegans Larval Development Molecular Cell 5331097-276538039210.1016/j.molcel.2013.12.01324440504 · doi ↗ · pubmed ↗
- 7Hoeckendorf Aylin Leippe Matthias 201291 SPP-3, a saposin-like protein of Caenorhabditis elegans, displays antimicrobial and pore-forming activity and is located in the intestine and in one head neuron Developmental & Comparative Immunology 3810145-305X 18118610.1016/j.dci.2012.05.00722677064 · doi ↗ · pubmed ↗
- 8Hoeckendorf Aylin Stanisak Mareike Leippe Matthias 2012627 The saposin-like protein SPP-12 is an antimicrobial polypeptide in the pharyngeal neurons of Caenorhabditis elegans and participates in defence against a natural bacterial pathogen Biochemical Journal 44520264-602120521210.1042/bj 2011210222519640 · doi ↗ · pubmed ↗