Genetic Diversity of Sodefrin-Variant Pheromones and Pheromone Responsiveness in Subspecies of the Japanese Sword-Tail Newt Cynops ensicauda
Tomoaki Nakada, Fumiyo Toyoda, Atsushi Tominaga, Koji Mochida, Makoto Yokosuka, Sakae Kikuyama

TL;DR
This study explores the genetic diversity of pheromones in two subspecies of the Japanese sword-tail newt and how these pheromones influence mating behavior.
Contribution
The study identifies a novel pheromone, caudarin, and reveals genetic and behavioral differences in pheromone use between newt subspecies.
Findings
The [Gln8]sodefrin precursor mRNA is widely expressed in the abdominal glands of both C. e. ensicauda and C. e. popei.
Caudarin attracts conspecific females, and silefrin is additionally expressed in C. e. popei males.
Imorin attracts C. e. ensicauda males but not C. e. popei males, indicating subspecies-specific pheromone responsiveness.
Abstract
Cynops newts inhabiting the Japanese archipelago have been classified into two species, namely, Cynops pyrrhogaster and C. ensicauda, with the latter being divided into two subspecies, namely, C. ensicauda ensicauda and C. e. popei. Experiments were conducted to determine whether mRNA expression of a sodefrin-like female attractant pheromone precursor is differently expressed in the abdominal glands of these subspecies. The results indicate that the precursor mRNA encoding [Gln8]sodefrin is widely detected in the abdominal glands of both subspecies. Preference tests demonstrated that [Gln8]sodefrin attracts conspecific females. This novel pheromone was named caudarin. Modification of the pheromone molecules was discussed in consideration of the geographical and chronological location of the newts’ habitat. Experiments were conducted to examine whether the abdominal glands of Cynops…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1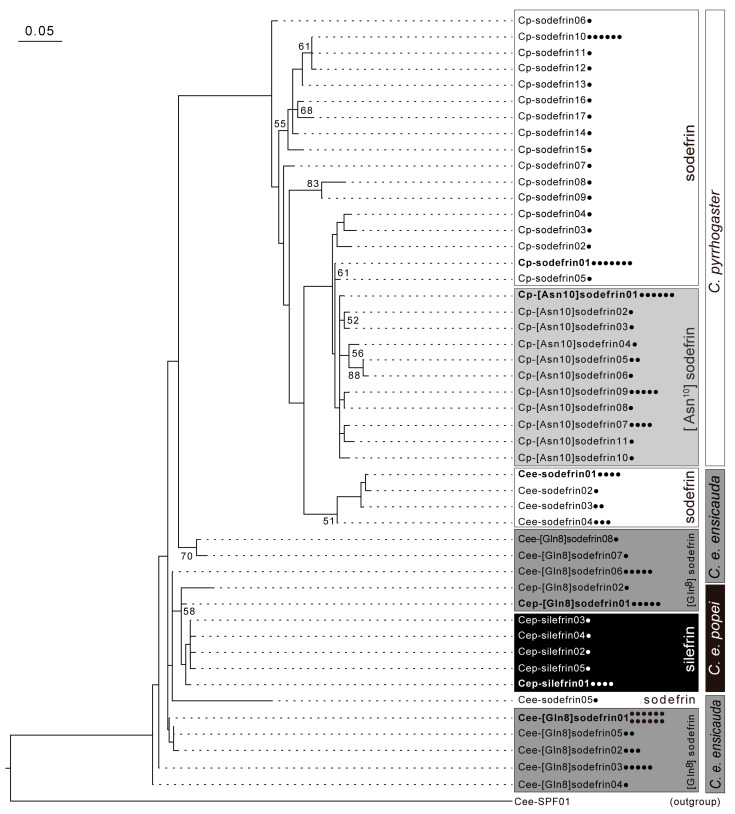 Figure 2
Figure 2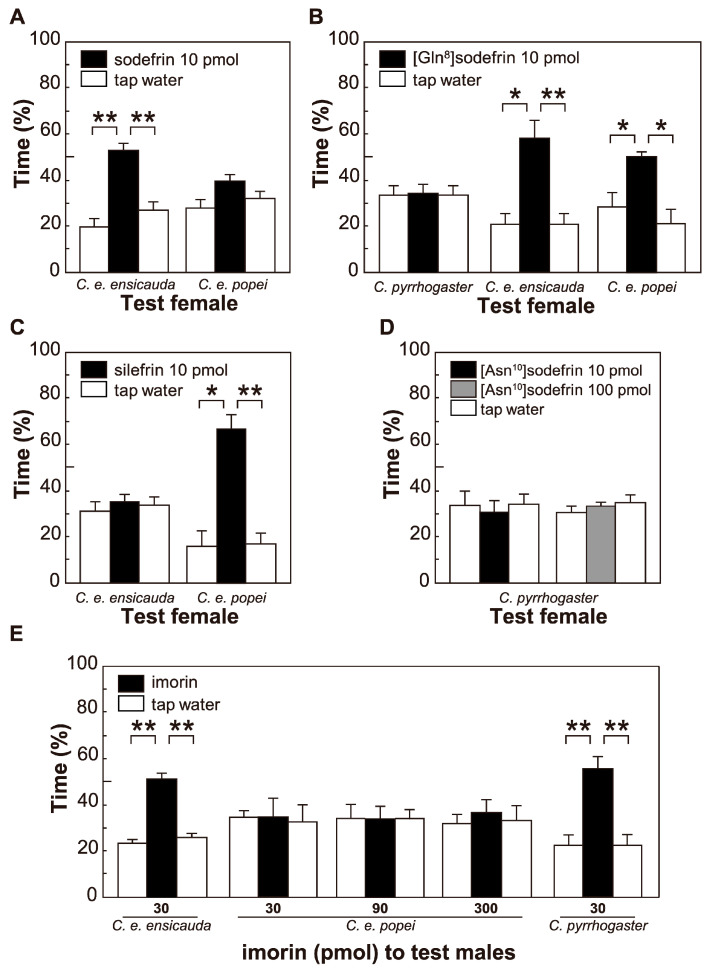 Figure 3
Figure 3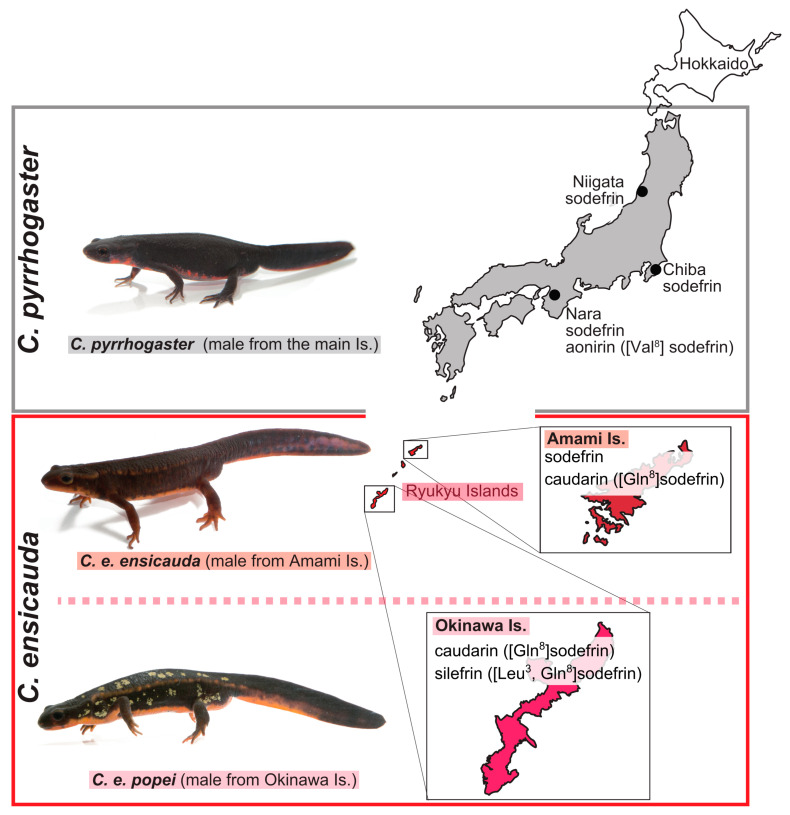 Figure 4
Figure 4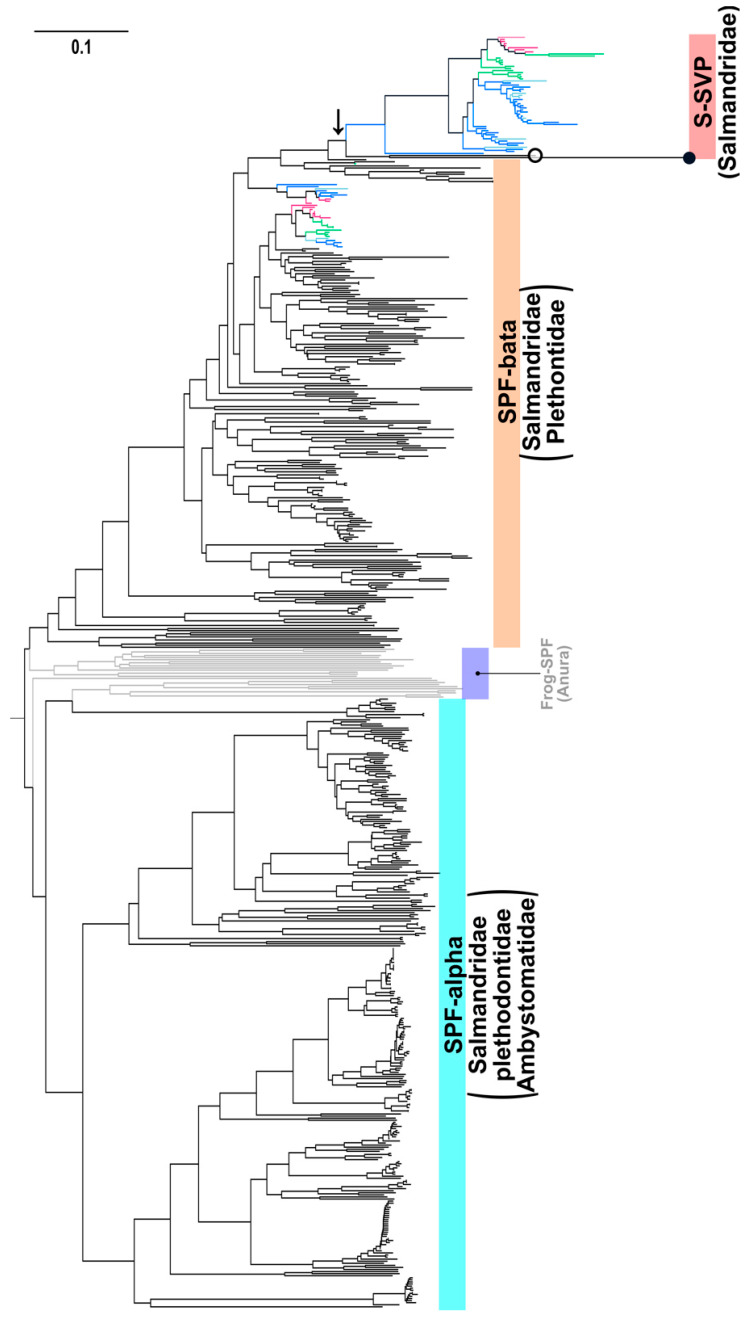 Figure 5
Figure 5 Figure 6
Figure 6- —JSPS
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInsect and Arachnid Ecology and Behavior · Plant and animal studies · Neurobiology and Insect Physiology Research
1. Introduction
In urodeles, sex pheromones play an important role by allowing both sexes to find a partner and maintain reproductive interactions, proceeding into courtship behavior that ends in successful fertilization. Accordingly, it is understood that structural changes of pheromones toward more effective molecules affect subsequent reproductive success and may even cause reproductive isolation and speciation [1]. Since sodefrin (Ser-Ile-Pro-Ser-Lys-Asp-Ala-Leu-Leu-Lys) emerged as the first pheromone to be identified in amphibians and the first peptide pheromone in vertebrates [2], the existence of several peptidic [3,4,5] and proteinaceous [6] pheromones in urodeles and even in an anuran species [7] has been elucidated. Sodefrin is generated in the abdominal gland of the male red-bellied newt Cynops pyrrhogaster, which inhabits the Shikoku and Kyushu islands as well as the main island of Japan. It is delivered through the male’s tail-fanning behavior, exerting a potent female-attracting activity toward sexually mature conspecific females at the initial stage of courtship behavior [2], and elicits a neural response in the sensory cells of the female vomeronasal organ [8,9,10]. Conversely, males of the congeneric newt C. ensicauda popei inhabiting Okinawa island were revealed to possess the pheromone silefrin, whose sequence differs from that of sodefrin by two amino acid residues. The effect of both pheromones appears to be species-specific. Sodefrin attracts C. pyrrhogaster females exclusively, whereas silefrin attracts only C. ensicauda females [3,11]. The primary structures of the precursor proteins of sodefrin and silefrin have been predicted from the nucleotide sequence of cloned cDNAs [11].
The sodefrin precursor protein comprises 189 amino acid residues, and the sodefrin sequence (residues 177–186) is located close to the COOH-terminus of the protein [11]. The existence of enzymes that cleave the Arg^176^-Ser^177^ and Lys^186^-Ile^187^ bonds to generate sodefrin from the precursor was demonstrated in the abdominal glands of sexually mature C. pyrrhogaster males [12]. In addition to the precursors of sodefrin and silefrin, the presence of two types of sodefrin precursor-like proteins named sodefrin precursor-like factors-alpha and beta (SPF-α and -β) has been predicted from the nucleotide sequences of mRNAs expressed in the exocrine glands of several salamanders [13,14,15,16,17,18,19]. To date, multiple SPF isoforms were found to be expressed in each species, and more than 500 SPF mRNAs have been submitted to GenBank, not only for salamanders but also for frogs [20] and even a shark (sequence accession numbers are listed in Table S1). It is notable that SPFs secreted by the mental gland in the terrestrially courting lungless salamanders (Plethodontidae) themselves function as courtship pheromones [14]. More recently, the cloacal gland in the aquatically courting newt Lissotriton helveticus was also shown to secrete the SPF pheromone [21].
It was previously demonstrated that genes encoding the precursor protein of the peptide pheromones and SPFs that are devoid of peptide pheromone sequences belong to the same multigene family, and that sodefrin precursors have a higher sequence similarity with the SPF-β subgroup. Thus, the β subgroup has been regarded as the genetic origin of the peptide pheromone precursor proteins [22,23]. In fact, sodefrin precursor transcripts were revealed to contain an extra 62-base insertion as compared with SPF transcripts; this insertion causes a frameshift in protein translation, generating the sodefrin sequence and enzymatic cleavage sites for sodefrin [22]. Multiple genes encoding sodefrin or silefrin precursors and SPFs are known to be expressed in the abdominal gland of an individual Cynops newt as well [4,22,23,24].
Different morphological characteristics between C. ensicauda populations inhabiting both the Amami and Okinawa islands have been noticed. Accordingly, Inger (1947) divided them into two subspecies, namely, C. e. ensicauda and C. e. popei, respectively [25]. Recently, on the basis of analyses of mitochondrial and nuclear DNA sequences, it was confirmed that C. ensicauda species should be divided into two genetically distinct subspecies [26,27].
We previously demonstrated that the abdominal gland of male C. e. popei generates silefrin but not sodefrin [3,11]. In this study, we conducted experiments to see whether the abdominal gland of another subspecies, C. e. ensicauda, expresses sodefrin-like pheromone precursor proteins that differ from the precursors for silefrin, and if so, whether C. e. ensicauda females respond to other Cynops pheromones such as silefrin and sodefrin in addition to the putative C. e. ensicauda pheromone. Experiments were also conducted to see whether the males of the C. ensicauda subspecies would respond to a male-attracting pheromone, imorin, which is known to be delivered from the oviducal epithelium of sexually mature C. pyrrhogaster females [5]. A phylogenetic tree of the sodefrin precursor and sodefrin-variant precursor proteins was constructed, incorporating the data obtained in the present experiments in order to understand the evolutionary relationships of the pheromone molecules among C pyrrhogaster, C. e. popei, and C. e. ensicauda.
2. Materials and Methods
2.1. Animals
To obtain the abdominal glands for the analysis of transcripts encoding sodefrin or its variant precursors or SPF proteins, eight sexually mature male C. e. ensicauda and C. e. popei were collected during the breeding season (from January to March) in the suburbs of Amami and Okinawa islands, respectively. Similarly, eight sexually mature male C. pyrrhogaster newts captured during the breeding season (from March to April) in Chiba of the main island of Japan were used for transcript analysis. In addition, eight female C. e. ensicauda and C. e. popei obtained in February and eight female C. pyrrhogaster collected in Chiba and Nara of the main island in April and May were kept for one month or more in a laboratory vivarium and were used for testing the biological activity of the pheromones or pheromone candidate. Prior to the test, they were treated with 1 IU ovine prolactin (Sigma, St. Louis, MO, USA) and 15 IU human chorionic gonadotropin (Asuka Pharmaceutical Co., Tokyo, Japan) every 2 days for 14 days to bring them to a reproductive state, ensuring their responsiveness to the pheromonal substances, according to the previously described methods [28]. All animal experiments were conducted in accordance with the approved protocols and the Guidelines for the Care and Use of Laboratory Animals of the Steering Committees for Animal Experimentation at Nippon Veterinary and Life Science University and Nara Medical University, Japan.
2.2. Sequence Determination of cDNA Encoding Sodefrin or Its Variant Precursors or SPF Proteins
Total RNA was individually extracted from the abdominal glands of C. e. ensicauda, C. e. popei, and C. pyrrhogaster using ISOGEN (Nippon Gene, Tokyo, Japan) and reverse-transcribed with reverse transcriptase (SuperscriptIII: Invitrogen, Carlsbad, CA, USA). Amplification by PCR of the partial cDNAs, covering the whole open reading frame of sodefrin or its variant peptide precursors, was performed with 50 pmol of each synthetic degenerate primer (sense: 5′-ACCCTAYTCCTTACTCTCCTAGCA-3′; antisense: 5′-RTCCCCGCCATGTGGAAAAT-3′) with 0.5 units Pfu-X polymerase (Greiner Bio-One, Tokyo, Japan) per 20 μL reaction solution. Amplified DNA fragments were treated with Ex Taq DNA polymerase (Takara Bio, Shiga, Japan) in the presence of dATP to add A-overhang then ligated into a pT7-blue T vector (Novagen, Madison, WI, USA), and the plasmids were used to transform JM109 competent cells (Takara Bio). Eight clones from different colonies obtained individually were subjected to sequence analysis. Nucleotide sequences were analyzed by an ABI PRISM 3100-Avant Genetic Analyzer (Applied Biosystems, Foster City, CA, USA) or a commercial sequencing service (Eurofins Genomics, Tokyo, Japan).
2.3. Phylogenetic Tree Reconstruction
A total of 192 nucleotide sequences encoding sodefrin or sodefrin-variant peptide precursor proteins, obtained from 24 individuals consisting of 8 of each of the C. e. ensicauda, C. e. popei, and C. pyrrhogaster newts, were translated into amino acid sequences using the program Serial Cloner [version 2.6.1; Serial Basics; http://serialbasics.free.fr/Serial_Cloner.html (accessed on 17 February 2025)]. Nucleotide and predicted amino acid sequences were aligned using the multiple sequence alignment programs MAFFT [version 7.450; https://mafft.cbrc.jp/alignment/software/ (accessed on 17 February 2025)] and ClustalX (version 2.1 [29]), respectively, with default parameters, and minor refinements were made manually. Nucleotide sequences were then aligned, and the transcripts were named using a “species (species and subspecies from which the transcript was cloned)–protein (a protein encoded by the transcript)–isoform (serial number of the protein isoform)–transcript (serial number of the transcript)” manner. For example, “transcript 1 that encodes sodefrin precursor isoform 2 derived from C. e. ensicauda” was designated as “Cee-sodefrin02-1”. These transcripts were deposited in GenBank (12 transcripts for Cee-[Gln^8^]sodefrin: LC861867-LC861878; 8 transcripts for Cee-sodefrin: LC861879-LC861886; 10 transcripts for Cee-SPF: LC861887-LC861896; 2 transcripts for Cep-[Gln^8^]sodefrin: LC861897-LC861898; 5 transcripts for Cep-silefrin: LC 861899-LC861903; 13 transcripts for Cee-SPF: LC861904-LC861916; 14 transcripts for Cp-[Asn^10^]sodefrin: LC861917-LC861930; 19 transcripts for Cp-sodefrin: LC861931-LC861949; and 11 transcripts for Cp-SPF: LC861950-LC861960).
Together with the 192 amino acid sequences obtained in this work, 534 sequences of sodefrin precursor, sodefrin-variant precursor, or SPF proteins were downloaded from GenBank (the list of proteins used in the tree is shown in Table S1) and also used in tree reconstruction. Phylogenetic trees were generated by applying the maximum-likelihood and the neighbor-joining algorithms to the aligned sequences, using MEGAX software (version 10.2.6 [30]) with 1000 bootstrap replications, with corrections for gaps or multiple substitutions with corrections for misalignments.
2.4. Test for Biological Activity of Sodefrin, Sodefrin-Variant Peptides, and Imorin
Test substances, such as sodefrin, silefrin, and [Gln^8^]sodefrin, were synthesized by Eurofins Genomics (Tokyo, Japan). Determination of the female-attracting activity of these peptides was performed according to previously described methods [28]. Briefly, a plastic container (diameter 37 cm) was filled with 3000 mL of tap water, and a female newt was kept in a smaller cylinder of stainless steel mesh (15 cm in diameter) that had been placed in the center of the container. The container was divided into three sectors. A sponge block (5.6 cm × 7.3 cm × 3.4 cm) was gently placed into each sector. One block contained the test substance dissolved in 100 mL water. It has previously been confirmed that the minimum effective amount of sodefrin to attract conspecific females is 10 pmol when sponge blocks adsorbed with the pheromone are placed in a container filled with 3000 mL of water [2]. Consequently, the same amount of synthetic sodefrin-like peptide was used to evaluate the female-attracting activity. For the male attractant tripeptide, 30 pmol imorin was used because its molecular weight is about one third that of the decapeptide pheromones. In cases where this amount proved ineffective, the test substance was increased to 3–10 times the initial amount. The other two contained tap water (blank). Thirty seconds after the introduction of the sponge blocks, the inner cylinder was removed. The position of the snout of the test animal was observed, and the time spent by the snout in each sector was video-recorded for 10 min. In each series of tests, eight females were used. The time spent by the snout of the test animals in each sector was analyzed statistically by Friedman’s two-way analysis of variance, followed by the Wilcoxon matched-pairs signed-rank test.
The male-attracting peptide imorin was synthesized by Peptide Institute Inc. (Osaka, Japan). Its activity in the males of Cynops species and subspecies was determined according to previously described methods [5].
3. Results
3.1. Analysis of Transcripts Encoding Sodefrin Precursor, Sodefrin-Variant Precursor, and SPF Proteins
After removal of the primer sequences, 128 nucleotide sequences encoding sodefrin precursor, sodefrin-variant precursor, or SPF proteins that were obtained from mRNAs expressed in the abdominal gland of each of the eight individual C. e. ensicauda, C. e. popei, and C. pyrrhogaster newts were determined. The encoded proteins were categorized according to the predicted amino acid sequence similarity (Table A1 and Table A2, simplified as Table 1). A total of 41 clones consisting of 20 types of transcripts encoding 13 isoforms of sodefrin precursor or sodefrin-variant precursor proteins from C. e. ensicauda, as well as 14 clones consisting of 7 types of transcripts encoding 7 isoforms of sodefrin-variant precursor proteins from C. e. popei, were identified. In C. e. ensicauda, 11 clones (8 types of transcripts) encoding 5 isoforms of sodefrin precursor, 30 clones (12 types of transcripts) encoding 8 isoforms of sodefrin-variant precursor ([Gln^8^]sodefrin precursor), and 23 clones (10 types of transcripts) encoding 10 isoforms of SPF proteins were detected; no clones encoding silefrin ([Leu^3^, Gln^8^]sodefrin) precursor were found. In C. e. popei, 6 clones consisting of 2 types of transcripts that were encoded with 2 isoforms of [Gln^8^]sodefrin precursor, and 8 clones consisting of 5 types of transcripts that were encoded with 5 isoforms of silefrin precursor were recognized, whereas no clones encoding sodefrin were detected in this subspecies. However, 50 clones (13 types of transcripts) encoding 10 isoforms of SPF proteins were detected.
Similarly, 64 nucleotide sequences encoding sodefrin precursor, sodefrin-variant precursor, or SPF proteins that were obtained from mRNAs expressed in the abdominal glands of eight individual C. pyrrhogaster were analyzed. As a result, 28 clones consisting of 19 types of transcripts encoding 17 isoforms of sodefrin precursor, 24 clones (16 types of transcripts) encoding 11 isoforms of sodefrin-variant precursor ([Asn^10^]sodefrin), and 12 clones (11 types of transcripts) encoding 11 types of SPF proteins were identified. However, neither clones encoding [Gln^8^]sodefrin nor those encoding silefrin were detected in this species.
In C. e. ensicauda, the most frequently encoded peptides were [Gln^8^]sodefrin (30 clones encoding 8 precursor isoforms) and silefrin in C. e. popei (8 clones encoding 5 precursor isoforms). In C. pyrrhogaster, along with sodefrin precursor proteins (28 clones encoding 17 precursor isoforms), proteins containing the sequence [Asn^10^]sodefrin (24 clones encoding 11 isoforms) were recognized. In the abdominal glands of both subspecies of C. ensicauda, a total of 28 SPF proteins were predicted from 36 transcripts derived from 99 clones (Table 1). The most frequently observed isoform in C. e. ensicauda was a certain type of SPF protein, designated as SPF isoform 10 (14 clones), and another type of SPF protein, designated as SPF isoform 3 (31 clones), was most common in C. e. popei. In C. pyrrhogaster, 11 types of SPF isoforms were predicted from the transcripts derived from 12 clones. The aforementioned data are summarized in Table 1; details are shown in Table A1. Structural differences in representative examples of precursor proteins of sodefrin and sodefrin-variant peptides most frequently observed in each taxon are shown in Figure 1. Among these peptides, [Gln^8^]sodefrin is a previously unknown sodefrin-variant peptide, but sodefrin, [Asn^10^]sodefrin, and silefrin all had homologous genes encoding their precursor sequences in GenBank. BLAST (version: 2.16) searches for known sequences showing the highest sequence similarity to each precursor protein at the amino acid sequence level revealed that sodefrin precursor (CAB53093.1) has 100% sequence similarity to the Cp-sodefrin precursor 01 and 99.5% sequence similarity to the Cp-[Asn^10^]sodefrin precursor 01; [Asn^10^]sodefrin precursor (AMO51436.1) has 94.7% sequence similarity to the Cee-Sodefrin precursor 01; and silefrin precursor (CAB53094.1) has 94.3% sequence similarity to the Cee-[Gln^8^]sodefrin precursor 01, 95.3% sequence similarity to Cep-[Gln^8^]sodefrin precursor 01, and 96.9% sequence similarity to Cep-silefrin precursor 01.
Several common amino acid substitutions were found among the predicted amino acid sequences from C. ensicauda (black-shadowed residues in Figure 1). However, many amino acid substitutions were shared among the [Gln^8^]sodefrin and silefrin precursor isoforms, including the insertion of three amino acid residues (Ser-Ala-Lys) at positions 124–126 (Figure 1). The sodefrin precursor protein isoform predicted from the mRNA expressed in C. pyrrhogaster (Cp-sodefrin01) showed 99.5% sequence similarity to the Cp-[Asn^10^]sodefrin 01 isoform, which is also expressed in C. pyrrhogaster, and showed 93.1% sequence similarity to the Cee-sodefrin01 isoform expressed in C. e. ensicauda. The majority of clones encoding precursor proteins of these peptides contained a predicted open reading frame of 567 bp, which encoded a precursor protein of 189 amino acid residues including a region containing hydrophobic amino acids that was presumed to be a signal peptide on the N-terminus, in addition to the sodefrin or [Asn^10^]sodefrin sequence near the C-terminus region. Similarly, the [Gln^8^]sodefrin precursor protein isoform predicted from the mRNA expressed in C. e. popei (Cep-[Gln^8^]sodefrin01) showed 98.4% sequence similarity to the Cep-silefrin01 isoform also expressed in C. e. popei, and 97.9% sequence similarity to the Cee-[Gln^8^]sodefrin01 isoform expressed in C. e. ensicauda. The majority of clones encoding precursor proteins of these peptides contained a predicted open reading frame of 576 bp. The predicted amino acid sequences homologous to sodefrin decapeptide were expected to be flanked by monobasic amino acid cleavage sites, arginine and lysine (Figure 1), for generating [Gln^8^]sodefrin or silefrin peptides from the precursor.
3.2. Phylogenetic Analysis of the Sodefrin Precursor-Related Proteins
The relationships of the sodefrin or sodefrin-variant precursor proteins predicted from the transcripts obtained from C. e. ensicauda, C. e. popei, and C. pyrrhogaster were inferred through phylogenetic trees constructed both with and without the 534 known sequences obtained from Genbank (Table S1). The reconstructed tree from analysis of all obtained protein sequences clearly indicated that sodefrin precursor-like proteins consisted of two types of SPF group (SPF-α and -β) in addition to a sodefrin and sodefrin-variant precursor (S-SVP) subgroup that belongs to the SPF-β subfamily. All of the proteins predicted from the transcripts obtained here from the Cynops newts were classified as members of the S-SVP or SPF group in the tree, and all the predicted SPF proteins obtained in this study were revealed to belong to the SPF-β subfamily (Figure A1). The molecular phylogenetic relationships of SPFs and S-SVPs obtained here were analyzed separately from the existing dataset (Figure 2 and Figure A2). In C. ensicauda, 20 S-SVP and 20 SPF isoforms were predicted from the 128 transcripts, and S-SVP isoforms were divided into three types of protein that were estimated to generate different peptides, such as sodefrin, silefrin, and [Gln^8^]sodefrin. Although 6 of 20 S-SVP isoforms and 7 of 20 SPF isoforms were found to be intra-subspecifically common, no isoforms were found to be shared with another subspecies (Figure 2 and Figure A2). In C. pyrrhogaster, 28 S-SVP isoforms and 11 SPF isoforms were predicted from the 64 transcripts, and S-SVPs were divided into two types of protein that were estimated to generate different peptides (sodefrin and [Asn^10^]sodefrin) (Table 1; Figure 2). Although 5 of the 28 S-SVP isoforms and 1 of the 11 SPF isoforms were found to be intraspecifically common, no isoforms were found to be shared with C. ensicauda.
3.3. Evaluation of the Pheromonal Activity of Sodefrin, Sodefrin Variants and Imorin
Behavioral responses to sodefrin, silefrin, [Gln^8^]sodefrin, and [Asn^10^]sodefrin of female newts belonging to C. e. ensicauda, C. e. popei, and C. pyrrhogaster are shown in Figure 3. When sodefrin was applied to C. ensicauda females, C. e. ensicauda females were attracted by sodefrin, whereas C. e. popei females were not (Figure 3A). When [Gln^8^]sodefrin was tested with the females of the two subspecies, females belonging to both C. ensicauda subspecies were attracted by it, whereas C. pyrrhogaster females were not (Figure 3B). When silefrin was tested with females of the two subspecies of C. ensicauda, C. e. popei females were attracted by the peptide, whereas C. e. ensicauda females were not (Figure 3C). It was also confirmed that C. pyrrhogaster females were attracted by neither 10 pmol nor 100 pmol [Asn^10^]sodefrin (Figure 3D).
Subsequently, sexually mature C. pyrrhogaster, C. e. ensicauda, and C. e. popei males were tested with imorin for its male-attracting activity. As shown in Figure 3E, both C. pyrrhogaster and C. e. ensicauda males responded positively to 30 pmol imorin. However, C. e. popei males were not attracted, even with 300 pmol imorin.
3.4. Obtaining and Organizing the Amino Acid Sequences of Known Sodefrin or Sodefrin-Variant Precursor and SPF Proteins
Five hundred and thirty-four amino acid sequences of sodefrin precursor and sodefrin-variant precursor proteins were downloaded from GenBank (Table S1). Ten transcripts that had been identified from three species of Salamandridae newts are available in the GenBank database. Three of the ten transcripts (GenBank accession numbers KM463792.1, KU213617.1, and KU213619.1) were registered as SPF transcripts, although their sequences differed from the other SPF precursor transcripts by the presence of a 62-base insertion causing a frameshift to generate the sodefrin-variant peptide sequence ([Asn^10^]sodefrin: Ser-Ile-Pro-Ser-Lys-Asp-Ala-Leu-Leu-Asn), of which cDNA clones were previously isolated from the mRNAs expressed in the abdominal gland of C. pyrrhogaster [24]. Because the tridecapeptide COOH-terminally extended with [Asn^10^]sodefrin-ISA peptide (Ser-Ile-Pro-Ser-Lys-Asp-Ala-Leu-Leu-Asn-Ile-Ser-Ala) was isolated from the abdominal glands of the newts in the previous study, we categorized these three transcripts into the sodefrin-variant peptide group rather than into the SPF, the tentative proteinaceous pheromone group [13]. In addition to the [Asn^10^]sodefrin transcripts, two transcripts lacking the 62-base insertion (AF446080.1 from Triturus carnifex and KJ402351.1 from Lissotriton helveticus) were also classified as sodefrin variant transcripts because of a frameshift that generated sequences homologous to sodefrin (Cys-Thr-Pro-Ser-Lys-Asp-Ala-Leu-Leu-Glu and His-Ile-Ala-Arg-Lys-Asp-Ala-Leu-Pro-Lys, respectively). As shown in Table S1, 524 amphibian SPF transcripts are available in GenBank. In urodeles, 500 SPF transcripts have been registered and identified mainly in Salamandroidea (300, 13, and 187 transcripts from 10 salamandrid newts, 1 ambystomatid salamander (Ambystoma mexicanum), and 30 plethodontid salamanders, respectively.). In anurans, 24 SPF transcripts have been registered in the database (18 transcripts from four hylid frogs, 2 transcripts from two nyctibatrachid frogs, 1 transcript from one pipid frog, and 3 transcripts from one ranid frog).
4. Discussion
In the present study, we performed analyses of transcripts encoding sodefrin precursor, sodefrin-variant precursor, and SPF proteins using the abdominal glands of male C. pyrrhogaster and two C. ensicauda subspecies, C. e. ensicauda and C e. popei. Our results revealed that the abdominal glands of both C. ensicauda subspecies possess a novel sodefrin-variant pheromone, [Gln^8^]sodefrin. Because this decapeptide exerts considerable female-attracting activity, this pheromone was designated caudarin (cauda- from the species name “ensicauda”, and -rin from sodefrin).
When the presence of the peptide pheromone silefrin was reported in the abdominal gland of C. e. popei from Okinawa, there was no clear explanation for the origin of this “sodefrin-like” pheromone [11]. Information concerning sodefrin-variant peptides in the abdominal gland of C. e. ensicauda obtained in the present study enables the explanation that silefrin diverged from caudarin as a result of a genetic mutation in a duplicated caudarin gene encoding the third amino acid residue, leucine instead of proline. It was also suggested that the precursor proteins of the novel pheromone candidate caudarin ([Gln^8^]sodefrin) and silefrin ([Leu^3^, Gln^8^]sodefrin) share a common protein ancestor that appeared more recently compared with that of S-SVP isoforms, and that predicted sodefrin precursor proteins expressed in C. e. ensicauda have a closer phylogenetic relationship with those expressed in C. pyrrhogaster as compared with any other precursors expressed in C. e. ensicauda. Similarly, we postulate that the caudarin gene originated from the sodefrin gene, and that the sodefrin gene disappeared from the abdominal gland of C. e. popei during the course of allopatric separation. As shown in the molecular phylogenetic analysis in Figure 3, the precursors of Cee-sodefrin have a closer relationship in sequence similarity with the precursors of Cp-sodefrin than those of the S-SVPs found in C. e. ensicauda. This supports the assumption that the ancestral sodefrin gene common to the Cp-sodefrin and Cee-sodefrin genes existed prior to the divergence of C. pyrrhogaster and C. ensicauda—the earliest divergence analyzed in this study—and may be the root gene for several of the S-SVP genes that arose from the gene duplications and were found in the subsequently emerging taxa. Similarly, the presence of the ancestral caudarin gene common to the Cee-caudarin and Cep-caudarin genes in the common ancestor of both subspecies prior to the divergence of the two C. ensicauda subspecies supports the assumption that the silefrin gene may have originated from the ancestral caudarin gene.
As shown in Table 1, the presence of [Asn^10^]sodefrin transcripts was recognized in seven out of eight specimens of C. pyrrhogaster. According to our previous work [24], we could not find [Asn^10^]sodefrin (SIPSKDALLN) as a peptide form, but [Asn^10^]sodefrin C-terminally extended with three amino acid residues (SIPSKDALLNISA) was detected among the fractions chromatographically separated from the abdominal gland extract. A preference test of [Asn^10^]sodefrin-ISA revealed that it had no female-attracting activity [24,27]. In the present experiment, [Asn^10^]sodefrin did not exhibit pheromonal activity (Figure 3D). Thus, we conclude that in C. pyrrhogaster, only sodefrin serves as a courtship pheromone, except for the case of aonirin, which is specifically involved in spousal communication in newts from the Nara region [3] (see Figure 4), and the [Asn^10^]sodefrin gene disappeared from C. ensicauda.
Additionally, two transcripts encoding sodefrin and sodefrin variants, which are active in terms of attracting conspecific females, were recognized in the abdominal gland of C. ensicauda subspecies (Table 1). Although both precursor protein mRNAs (i.e., sodefrin and caudarin in C. e. ensicauda; caudarin and silefrin in C. e. popei) were confirmed to be expressed, few specimens were found to possess both transcripts. We presume that once speciation of C. e. ensicauda and C. e. popei is complete, they will possess exclusively either caudarin or silefrin, provided that females exhibit stronger preferences for either. It is notable that pheromonal peptides, such as sodefrin, caudarin, and silefrin, that were identified in the males of the Cynops species and subspecies specifically attract their own groups’ female partners, and females are not attracted to sex pheromones that are not possessed by the males in their population. This means that receptors for each pheromone were newly equipped within the females’ vomeronasal organs during speciation and subspeciation, losing the receptors for pheromones that were once possessed but then lost by the male partner.
Imorin is a male-attracting pheromone identified in the oviducal epithelium of C. pyrrhogaster [5]. Although it has been confirmed that this tripeptide pheromone is effective in attracting sexually mature conspecific males, our results show that it attracts C. e. ensicauda males but not those of C. e. popei (Figure 3E); that is, the species or subspecies for which imorin was found to be effective were those for which sodefrin was effective. We thus speculate that the common ancestor of C. pyrrhogaster and C. ensicauda maintained spousal communication via sodefrin (male pheromone) and imorin (female pheromone), which have been shown to be used conservatively in the two species even today. It remains to be investigated, however, whether C. e. popei females utilize a male-attracting pheromone comparable to imorin.
As stated by Houck et al. [14], the existence of precursor proteins devoid of pheromonal peptide sequences generated as a result of gene duplication has been documented. Furthermore, the extract of the mental gland of the terrestrially courting salamanders (Plethodontidae) has been shown to exert pheromonal activity. These novel proteins were designated sodefrin precursor-like factors (SPFs) due to their sequence similarity with the sodefrin precursor protein. A comparison of the transcripts of the SPF family proteins and the sodefrin precursor protein revealed that the latter contains an additional 62-base insertion. This insertion is hypothesized to have induced a frame shift, resulting in the formation of the sodefrin peptide sequence and the subsequent emergence of enzymatic cleavage sites [22]. Nevertheless, the presence of transcripts encoding precursors with sodefrin-homologous sequences has been documented in Triturus carnifex and Lissotriton helveticus, the only taxon confirmed to utilize decapeptide pheromones is the genus Cynops so far; and the emergence of S-SVPs is estimated to have occurred after 34.3–26.5 MYA, coinciding with the divergence of the Asian newts that include this genus [22,31].
To date, the significance of SPF as a sex pheromone in Cynops newts is not well understood, but SPF transcripts have been cloned relatively frequently, especially in C. ensicauda (Table 1). Molecular phylogenetic analysis of the SPF proteins observed in this study identified a particular group of homologous SPF genes across the subspecies (the clade enclosed by the gray square in Figure A2). Because SPF may act as a sex pheromone in other species [14], these inter-subspecifically conserved SPFs expressed in C. ensicauda might also exhibit some pheromonal activity during courtship behavior.
According to previous analyses [26,31], genetic isolation between C. pyrrhogaster and C. ensicauda is estimated to have been established 13.75–12.40 million years ago (MYA), followed by the divergence between C. e. ensicauda and C. e. popei at 5.21 MYA. Therefore, the origin of reproductive communication by sodefrin and imorin, which is common to both species, must have occurred before the C. pyrrhogaster/C. ensicauda divergence. The origin of caudarin, which is common to the two subspecies of C. ensicauda but absent in C. pyrrhogaster, can be dated to 8.54–7.19 MYA, i.e., between the divergence of the two species and the divergence of the C. ensicauda subspecies. Similarly, silefrin, which is expressed only in C. e. popei males, is assumed to have acquired its function as a sex pheromone after the divergence of the two C. ensicauda subspecies (5.21 MYA). Focusing on the distribution of conserved amino acid residues among the sequences of precursor proteins of sodefrin, caudarin, and silefrin (Figure 1), it is obvious that the genes for caudarin and silefrin precursors were derived from those for sodefrin and caudarin precursors, respectively. Thus, accompanying the generation of caudarin and silefrin genes, in addition to the loss of [Asn^10^]sodefrin and sodefrin genes, speciation and subspeciation occurred successively. It is notable, as has been pointed out previously [26,31], that time periods of 12.40 and 5.21 million years were required for speciation and subspeciation, respectively.
A sex pheromone such as sodefrin is one of the critical factors for reproductive success and should be under high selective pressure for resistance to loss-of-function mutations. Conversely, the acquisition of a novel pheromone that is preferred by mating partners over an existing pheromone used by reproductive competitors would be an evolutionarily adaptive event. The multiplexing and diversity of male sex pheromone genes in newts demonstrated in this study may provide a solution to this conflict by not only conservatively maintaining the existing sex pheromone system (e.g., sodefrin in C. e. ensicauda) but also by exploring the co-evolution of the partner’s response system to acquire a new and more effective pheromonal communication system (e.g., silefrin in C. e. popei). In fact, such multiplexing has been observed in other vertebrates (e.g., main urinary proteins and exocrine gland-secreting peptides in rodents [32,33]), as well as in amphibians (e.g., SPFs, plethodontid modulating factors [13,34], and plethodontid receptivity factors [6,35]).
5. Conclusions
In conclusion, the present experiment provided evidence for the two Cynops subspecies, C. ensicauda ensicauda and C. e. popei, inhabiting the Amami and Okinawa Islands in isolation, having come to possess different courtship pheromones, namely, caudarin and silefrin, accompanying the loss of the [Asn^10^]sodefrin precursor gene and the acquisition of the caudarin precursor gene in the course of speciation, and similarly, the loss of the sodefrin precursor gene along with the acquisition of the silefrin precursor gene in the course of subspeciation (Figure 4).
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Arnold S.J. Reagan N.L. Verrell P.A. Reproductive isolation and speciation in plethodontid salamanders Herpetologica 199349216228
- 2Kikuyama S. Toyoda F. Ohmiya Y. Matsuda K. Tanaka S. Hayashi H. Sodefrin: A female-attracting peptide pheromone in newt cloacal glands Science 19952671643164510.1126/science.78864527886452 · doi ↗ · pubmed ↗
- 3Yamamoto K. Kawai Y. Hayashi T. Ohe Y. Hayashi H. Toyoda F. Kawahara G. Iwata T. Kikuyama S. Silefrin, a sodefrin-like pheromone in the abdominal gland of the sword-tailed newt, Cynops ensicauda FEBS Lett.200047226727010.1016/S 0014-5793(00)01455-110788624 · doi ↗ · pubmed ↗
- 4Nakada T. Toyoda F. Iwata T. Yamamoto K. Conlon J.M. Kato T. Kikuyama S. Isolation, characterization and bioactivity of a region-specific pheromone, [Val 8]sodefrin from the newt Cynops pyrrhogaster Peptides 20072877478010.1016/j.peptides.2006.12.00617275139 · doi ↗ · pubmed ↗
- 5Nakada T. Toyoda F. Matsuda K. Nakakura T. Hasunuma I. Yamamoto K. Onoue S. Yokosuka M. Kikuyama S. Imorin: A sexual attractiveness pheromone in female red-bellied newts (Cynops pyrrhogaster)Sci. Rep.201774133410.1038/srep 4133428120945 PMC 5264602 · doi ↗ · pubmed ↗
- 6Rollmann S.M. Houck L.D. Feldhoff R.C. Proteinaceous pheromone affecting female receptivity in a terrestrial salamander Science 19992851907190910.1126/science.285.5435.190710489368 · doi ↗ · pubmed ↗
- 7Wabnitz P.A. Bowie J.H. Tyler M.J. Wallace J.C. Smith B.P. Aquatic sex pheromone from a male tree frog Nature 199940144444510.1038/4672410519546 · doi ↗ · pubmed ↗
- 8Toyoda F. Kikuyama S. Hormonal influence on the olfactory response to a female-attracting pheromone, sodefrin, in the newt, Cynops pyrrhogaster Comp. Biochem. Physiol. Part B Biochem. Mol. Biol.200012623924510.1016/S 0305-0491(00)00202-910874171 · doi ↗ · pubmed ↗