Genetic Associations of ACOX2 Gene with Milk Yield and Composition Traits in Chinese Holstein Cows
Hui Cao, Zhe Wang, Lingna Xu, Bo Han, Dongxiao Sun

TL;DR
This study shows that the ACOX2 gene influences milk production and composition in Chinese Holstein cows, offering new genetic markers for breeding.
Contribution
The study identifies specific SNPs in the ACOX2 gene linked to milk traits and suggests their use for genomic selection in dairy cattle.
Findings
Five SNPs in the ACOX2 gene significantly affect milk yield, fat, and protein in Chinese Holstein cows.
The SNP rs109066086 may regulate ACOX2 transcription by creating binding sites for transcription factors.
Haplotype blocks of SNPs are associated with 305-day milk, fat, and protein yields.
Abstract
This study investigated the genetic effects of the acyl-CoA oxidase 2 (ACOX2) gene on milk production traits in Chinese Holstein cows. We identified five single nucleotide polymorphisms (SNPs) that significantly influenced 305-day milk, fat, and protein yields. In addition, we found that rs109066086 may regulate the transcriptional activity of the ACOX2 gene. Overall, our research highlights the genetic effects of the ACOX2 gene on milk yield and composition traits in dairy cattle and provides valuable molecular markers for genomic selection. In our previous studies on the liver proteome of Holstein cows, the acyl-CoA oxidase 2 (ACOX2) gene was identified as a promising candidate for milk traits, being involved in the processes of fatty acid metabolism and bile acid formation. Herein, we evaluated its genetic effects on milk production traits in 922 Chinese Holstein cows. By sequencing…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
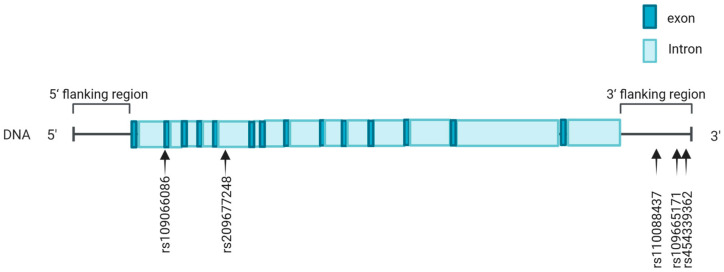 Figure 1
Figure 1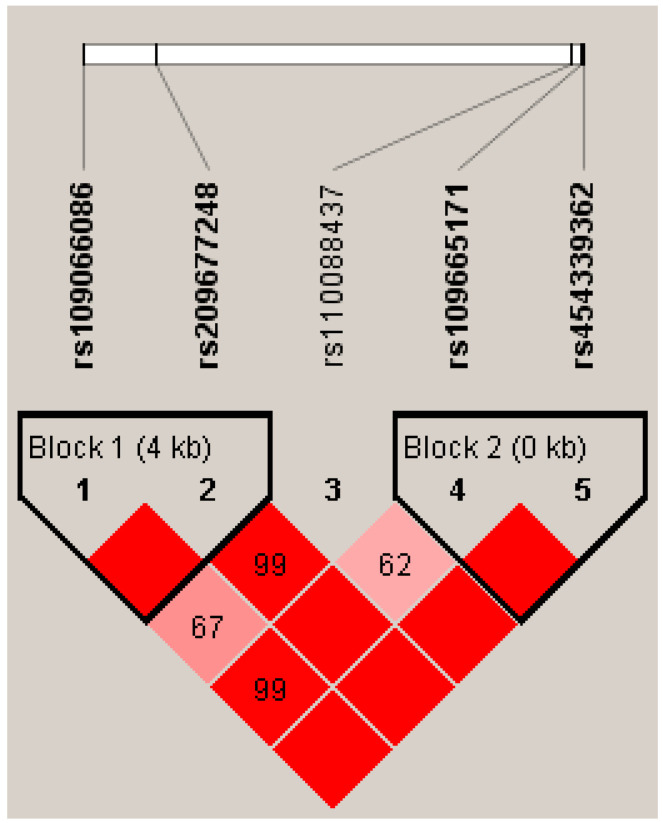 Figure 2
Figure 2- —National Key R&D Program of China
- —S&T Program of Hebei
- —Science and Technology Program of Inner Mongolia Autonomous Region
- —Program for Changjiang Scholars and Innovation Research Team in University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenetic and phenotypic traits in livestock · Reproductive Physiology in Livestock · Meat and Animal Product Quality
1. Introduction
Milk is an essential source of nutrients in the human diet, being rich in carbohydrates, protein, vitamins, and minerals [1]. Numerous studies have highlighted the biological functions of milk, such as its ability to lower blood pressure and enhance immune function, which has led to a higher demand for high-quality milk [1,2,3]. Improving both milk yield and quality has always been the primary goal of dairy cattle breeding.
Before the advent of genomic selection (GS), progeny testing was the primary method used in dairy cattle breeding. Although this method is highly accurate, it has a long cycle, typically 5 to 6 years [4]. GS has reduced the dairy breeding cycle to approximately two years, improved breeding accuracy, and reduced breeding costs by 92% [5]. Studies have shown that incorporating functional gene information into genomic-selection chips can improve the accuracy of genomic estimation of breeding values [6,7,8]. However, milk yield and composition traits are controlled by polygenes with minor effects [5,9]. So far, only diacylglycerol O-acyltransferase 1 (DGAT1), growth hormone receptor (GHR), and ATP-binding cassette subfamily G member 2 (ABCG2) genes have been confirmed as causal genes for milk production traits [10,11,12,13]. The development of sequencing technologies and multi-omics approaches has made it easier to identify functional genes associated with milk production traits in dairy cattle.
In our previous studies, proteomic analyses revealed that the expression of the ACOX2 was significantly higher in the livers of Holstein cows during the peak of lactation compared to the dry period (fold change ≥ 1.5, p = 6.75 × 10^−3^) [14]. The ACOX2 gene in bovines is located on chromosome 22 and has 14 exons with a gene length of 30,884 bp. It is involved in fatty acids biosynthesis by oxidizing long-chain fatty acids to short-chain fatty acids in the peroxisome and catalyzing the conversion of cholesterol to bile acids (BAs) by oxidizing CoA ester intermediates of BAs. ACOX2 deficiency leads to the accumulation of BAs and intermediates [15]. BAs can act as signaling molecules to regulate lipid, glucose, and energy metabolisms, and they promote the absorption of fat and fat-soluble vitamins from food [16,17]. Previous studies have found that increasing ACOX2 levels helps eliminate Staphylococcus aureus in bovine mammary cells [18]. In addition, ACOX2 is also associated with the metabolism of subcutaneous fat in beef cattle [19]. Of note, the ACOX2 gene is located 3.14 cM from the QTL (ID: 2487) for milk protein percentage in dairy cattle [20]. Moreover, ACOX2 is close to the SNP ARS-BFGL-BAC-33314, which is associated with milk fat and protein traits, with 1.8 Mb, and the SNP ARS-BFGL-NGS-61008, which is associated with the milk protein percentage, with 0.2 Mb [21]. The kyoto encyclopedia of genes and genomes (KEGG) pathway analysis (https://www.kegg.jp/kegg/pathway.html, accessed on 12 January 2025) also indicated that this gene is involved in primary bile acid biosynthesis, metabolic pathways, and the PPAR signaling pathway. Therefore, the ACOX2 gene is regarded as a promising candidate for milk traits.
Therefore, the purpose of this study was to systematically evaluate whether the ACOX2 gene has significant genetic effects on milk yield and composition traits, thereby providing valuable genetic markers for genomic selection programs in dairy cattle.
2. Materials and Methods
2.1. Animals and Phenotypic Data Collection
The experimental population comprised 922 Chinese Holstein cows from 44 sire families. These cows were collected from the 21 dairy farms of Beijing Sunlon Livestock Development Co., Ltd. (Beijing, China) and were raised under uniform feeding conditions; their pedigree information and phenotypic data (including 305-day milk yield, milk fat yield, milk fat percentage, milk protein yield, and milk protein percentage) were provided by the Beijing Dairy Cattle Center (Beijing, China). Descriptive statistics of the phenotypic values for milk production traits across the two lactations are presented in Table S1.
2.2. Genomic DNA Extraction
Genomic DNA was extracted from frozen semen of 44 Holstein bulls using the optimized high-salt method and from whole blood samples of 922 daughters using the TIANamp Blood DNA Kit (Tiangen, Beijing, China). The quantity and quality of the extracted DNA was assessed using a NanoDrop 2000 Spectrophotometer (Thermo Scientific, Hudson, NH, USA) and 2% agarose gel electrophoresis, respectively.
2.3. SNP Identification and Genotyping
Using Primer 3.0 (https://primer3.ut.ee/, accessed on 20 May 2024), 21 pairs of primers were designed (Table S2) to amplify the entire coding region and 2000 bp of the 5′ and 3′ flanking regions for the ACOX2 gene (Gene ID: 514969) and were synthesized at the Beijing Genomics Institute (BGI, Beijing, China). Genomic DNA from 44 bulls was diluted to a final concentration of 50 ng/μL. The 44 DNA samples were randomly divided into two DNA pools with 22 samples each. We performed the PCR for the amplifications (Table S2). PCR products were detected via 2% agarose gel electrophoresis, and the qualified products were sequenced at Beijing Qingke Xinye Biotechnology Co., Ltd. (Beijing, China). Chromas software 2.6 was used to check the sequence diagram and compare it with the bovine reference genome sequence (ARS-UCD1.2) to identify polymorphic sites. Genotypes of individual SNPs of 922 cows were identified using the Genotyping by Target Sequencing (GBTS) method at Shijiazhuang MolBreeding Biotechnologu Co., Ltd. (Shijiazhuang, China).
2.4. Linkage Disequilibrium (LD) Estimation
We estimated the linkage disequilibrium (LD) between the identified SNPs using Haploview 4.2 (Broad Institute of MIT and Harvard, Cambridge, MA, USA). The magnitude of LD is represented by the D’ value, and haplotypes with frequencies greater than 0.05 were retained.
2.5. Association Analyses
We performed association analyses between the SNPs and/or haplotypes and five milk traits using the mixed procedure of SAS 9.4 software (SAS Institute Inc., Cary, NC, USA) with the following animal model:
where y represents the phenotypic value of each trait for each cow; μ is the population mean; HYS includes the fixed effects of farm (1–21: 21 farms), calving year (1–4: 2012–2015), and season (1: April–May; 2: June–August; 3: September–November; 4: December–March); is the month of calving as a covariant, where b is its regression coefficient; is the effect of the genotype or haplotype combination; a is the individual random additive genetic effect, distributed as , with the additive genetic variance δ_a^2; and e is the random residual, distributed as , with the identity matrix I and the residual error variance .
Also, we calculated the additive effect (a), dominant effect (d), and substitution effect (α) using the formulas:
where AA, BB, and AB are the least squares means of each trait for the respective genotypes, p is the frequency of allele A, and q is the frequency of allele B.
The genetic effect of an SNP on a particular trait can be expressed by the proportion of phenotypic variation it explains. The formula for calculating the phenotypic variance explained by an SNP is as follows:
where p and q are the allele frequencies for allele A and allele B, α is the substitution effect, and is the phenotypic variance of the target trait [22].
2.6. Prediction of the Changes in Transcription Factor Binding Sites
We used JASPAR software (http://jaspar.genereg.net/, accessed on 12 February 2025) to predict the changes in the transcription factor binding sites (TFBSs) of the SNP rs109066086, which is in the 5′ UTR of the ACOX2 gene. We inputted the sequence of each allele and its 10 bp flanking sequences separately to predict whether the transcription factor binding sites were altered (relative score ≥ 0.90).
3. Results
3.1. SNP Identification in ACOX2 Gene
We identified a total of five SNPs in the ACOX2 gene (Figure 1), including one, rs109066086, in the 5′ UTR; rs209677248 in intron 5; and three, rs110088437, rs109665171, and rs454339362, in the 3′ flanking region. The genotypic and allelic frequencies of all the identified SNPs are summarized in Table 1.
3.2. Genetic Associations Between SNPs and Milk Yield and Composition Traits
By performing phenotype–genotype association analysis between the five SNPs of ACOX2 and five milk traits (Table 2), we found that the SNP rs109066086 was significantly associated with the 305-day milk yield, milk fat yield, and milk protein yield in both the first and second lactation (p ≤ 4.03 × 10^−2^); rs109665171 was significantly associated with milk protein yield (p = 1.65 × 10^−2^); and rs454339362 was significantly associated with the 305-day milk yield, milk fat percentage, and milk protein yield in the first lactation (p ≤ 9.30 × 10^−3^). All five SNPs—rs109066086, rs209677248, rs110088437, rs109665171, and rs454339362—were significantly associated with the 305-day milk yield (p ≤ 8.00 × 10^−3^), milk fat yield (p ≤ 5.50 × 10^−3^), and milk protein yield (p ≤ 5.00 × 10^−4^) in the second lactation. Additionally, rs209677248 and rs109665171 were associated with the milk protein percentage (p = 3.37 × 10^−2^) and milk fat percentage (p = 1.17 × 10^−2^) in the second lactation, respectively. Additive, dominant, and allelic substitution effects of the five SNPs in the ACOX2 gene were also calculated (Supplementary Table S3). We also calculated the phenotypic variance ratios (PVRs) of five SNPs for milk production traits and found that rs209677248 had higher PVRs for the 305-day milk, protein, and fat yields and milk protein percentage in the second lactation, with values of 2.94%, 2.58%, 1.83%, and 4.90%, respectively (Supplementary Table S4).
3.3. Genetic Associations Between Haplotypes and Milk Yield and Composition Traits
Using Haploview 4.2, we identified two haplotype blocks formed by four SNPs (D’ = 1; Figure 2). Block 1 included rs109066086 and rs209677248, in which the frequencies of H1 (CC), H2 (TC), and H3 (CG) were 0.47, 0.29, and 0.24, respectively. Block 2 included rs109665171 and rs454339362, in which the frequencies of H1 (TT), H2 (CC), and H3 (CT) were 0.33, 0.10, and 0.57, respectively. Block 1 showed significant associations with the 305-day milk, fat, and protein yields in both lactations (p ≤ 8.60 × 10^−3^; Table 3). Block 2 showed significant associations with the milk and protein yields in the first lactation (p ≤ 1.03 × 10^−2^; Table 3) and associations with the milk, fat, and protein yields and milk fat percentage in the second lactation (p ≤ 9.10 × 10^−4^; Table 3).
3.4. Transcription Factor Binding Site Changes Caused by SNPs
With JASPAR software, the allele T of the SNP rs109066086 in the 5′ UTR of the ACOX2 gene was predicted to create a binding site for the transcription factors NR2C2 (RS = 0.91) and TFAP4 (RS = 0.90) (Table 4), implying its potential regulatory role in ACOX2 expression.
4. Discussion
Based on our previous analysis of the liver proteomes across dry and lactation stages in Holstein cows, the ACOX2 gene was identified as a promising candidate for milk production traits. In this study, our results provided the first evidence that the ACOX2 gene indeed has significant genetic influences on milk yield and composition traits in dairy cattle.
In human breast cancer, the expression of the ACOX2 gene was significantly decreased (log2 ^(fold change)^ = −2.21, p = 6.34 × 10^−14^) [18]. Previous research has shown that the upregulation of ACOX2 can increase the production of peroxisomes and reactive oxygen species (ROS), thereby facilitating the elimination of Staphylococcus aureus in bovine mammary epithelial cells [23]. The ACOX2 gene has been associated with physiological changes in subcutaneous and visceral fat in beef cattle [19]. A previous study has shown that the expression of ACOX2 is downregulated in the jejunums of broilers suffering from necrotic enteritis, and this alteration may lead to disturbances in lipid metabolism, thereby compromising their immune response [24]. The above studies suggest that ACOX2 has a critical role in the maintenance of body homeostasis and lipid metabolism.
In mammalian liver tissue, ACOX2 plays a critical role in lipid metabolism. ACOX2 is the rate-limiting enzyme in the β-oxidation of branched-chain fatty acids, catalyzing the oxidation of long-chain fatty acids to short-chain fatty acids [16,25]. ACOX2 can also convert cholesterol to bile acids (BAs), which activate the farnesoid X receptor (FXR). The activation of FXR, in turn, induces the expression of phosphoenolpyruvate carboxykinase (PEPCK) and regulates glucose levels in human and rat hepatocytes, as well as in mouse liver [26]. In the gut microbiota, BAs are essential for the absorption, transport, and metabolism of dietary fats and fat-soluble vitamins [27]. These studies highlight the important role of ACOX2 in lipid regulation. This is consistent with the result of the present study that ACOX2 significantly affects milk fat traits. In addition, it was significantly associated with milk yield and milk protein traits in this study. This is probably due to the strong genetic correlation between milk traits.
SNPs can cause phenotypic variation by affecting gene function or expression [28,29]. In this study, the SNP rs109066086 in the 5′ UTR of ACOX2 changed a predicted TFBS for the transcript factors NR2C2 and TFAP4. NR2C2 was reported to bind the promoters of 9 downregulated lncRNAs in olaparib-treated human uveal melanoma cells [30]. TFAP4 can suppress the transcription of the human homolog of murine double minute 2 (HDM2) gene in HCT116 cells [31,32]. For rs109066086, the TT genotype exhibited significantly lower 305-day milk, fat, and protein yields compared with the CC genotype. This may be due to the allele T producing TFBSs for the transcript factors NR2C2 and TFPA4, thereby impacting ACOX2 expression. Thus, the SNP rs109066086 could be a potential key mutation affecting milk fat traits and deserves further in-depth study.
In addition, we found that the H3H3 haplotype combination from block 1 was associated with higher milk, fat, and protein yields. It is possible to consider adding these SNP combinations into the chip array for a genomic selection program in dairy cattle. In the future, the dual-luciferase report assay (DLRA), chromatin immunoprecipitation (ChIP), electrophoretic mobility shift assay (EMSA), single base mutation, and other tests can be used to verify how these mutations regulate the biological functions of the ACOX2 genes and their influence on milk production, thus providing valuable molecular markers for the genomic selection of dairy cattle.
5. Conclusions
In our previous proteomic research on the livers of Holstein cows, the ACOX2 gene was identified as a potential candidate gene associated with milk production traits. This study identified polymorphisms in the ACOX2 gene and provided the first evidence that the ACOX2 gene had significant genetic effects on the milk yield and composition traits in dairy cattle. The SNP rs109066086 in the 5′ UTR was predicted to altered the binding sites of the transcription factors NR2C2 and TFAP4, suggesting that they might regulate ACOX2 expression. Our findings provided valuable genetic markers for a genomic selection program in dairy cattle breeding.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Pereira P.C. Milk Nutritional Composition and Its Role in Human Health Nutrition 20143061962710.1016/j.nut.2013.10.01124800664 · doi ↗ · pubmed ↗
- 2Jauhiainen T. Korpela R. Milk Peptides and Blood Pressure J. Nutr.2007137825 S 829S 10.1093/jn/137.3.825S 17311982 · doi ↗ · pubmed ↗
- 3Rumbold P. Mc Cullogh N. Boldon R. Haskell-Ramsay C. James L. Stevenson E. Green B. The Potential Nutrition-, Physical- and Health-Related Benefits of Cow’s Milk for Primary-School-Aged Children Nutr. Res. Rev.202235506910.1017/S 095442242100007 X 33902780 · doi ↗ · pubmed ↗
- 4Thomasen J.R. Liu H. Sørensen A.C. Genotyping More Cows Increases Genetic Gain and Reduces Rate of True Inbreeding in a Dairy Cattle Breeding Scheme Using Female Reproductive Technologies J. Dairy Sci.202010359760610.3168/jds.2019-1697431733861 · doi ↗ · pubmed ↗
- 5Schaeffer L.R. Strategy for Applying Genome-Wide Selection in Dairy Cattle J. Anim. Breed. Genet.200612321822310.1111/j.1439-0388.2006.00595.x 16882088 · doi ↗ · pubmed ↗
- 6Liu A. Lund M.S. Boichard D. Karaman E. Fritz S. Aamand G.P. Nielsen U.S. Wang Y. Su G. Improvement of Genomic Prediction by Integrating Additional Single Nucleotide Polymorphisms Selected from Imputed Whole Genome Sequencing Data Heredity 2020124374910.1038/s 41437-019-0246-731278370 PMC 6906477 · doi ↗ · pubmed ↗
- 7Zhang Z. Ober U. Erbe M. Zhang H. Gao N. He J. Li J. Simianer H. Improving the Accuracy of Whole Genome Prediction for Complex Traits Using the Results of Genome Wide Association Studies P Lo S ONE 20149 e 9301710.1371/journal.pone.009301724663104 PMC 3963961 · doi ↗ · pubmed ↗
- 8de Las Heras-Saldana S. Lopez B.I. Moghaddar N. Park W. Park J.-E. Chung K.Y. Lim D. Lee S.H. Shin D. van der Werf J.H.J. Use of Gene Expression and Whole-Genome Sequence Information to Improve the Accuracy of Genomic Prediction for Carcass Traits in Hanwoo Cattle Genet. Sel. Evol.2020525410.1186/s 12711-020-00574-232993481 PMC 7525992 · doi ↗ · pubmed ↗