Cancer‐associated fibroblast heterogeneity in chordoma†
Jack C Henry, Angus J M Cameron

TL;DR
This paper explores the role of cancer-associated fibroblasts in chordoma, a rare tumor, and how they contribute to tumor progression and poor outcomes.
Contribution
The study identifies an inflammatory CAF subtype in chordoma and suggests its role in driving aggressive tumor behavior.
Findings
Chordoma cells show mixed epithelial and mesenchymal traits.
Inflammatory CAFs are linked to poor prognosis and tumor invasion.
Integrating CAF studies across tumor types could reveal new therapeutic insights.
Abstract
In solid tumours, malignant cells develop relationships with nonmalignant stromal cells to support tumour growth, tissue structure, and nutrient supply. In a recent issue of this journal, Zheng and Guo catalogue the cellular landscape in chordoma using a combination of single‐cell and spatial transcriptomics. Despite the mesenchymal nature of chordoma, malignant cells retain expression of epithelial markers, in addition to mesenchymal, partial‐EMT, and stem‐cell signatures. Cancer‐associated fibroblasts (CAFs) are among the most abundant stromal cells and the authors define an inflammatory CAF subtype (iCAF), which is associated with poor outcome and tumour invasion. It is proposed that iCAFs arise from normal fibroblasts during malignant tumour progression and play a causative role in driving an invasive poor prognosis tumour phenotype. Recent reports by this and other groups have…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
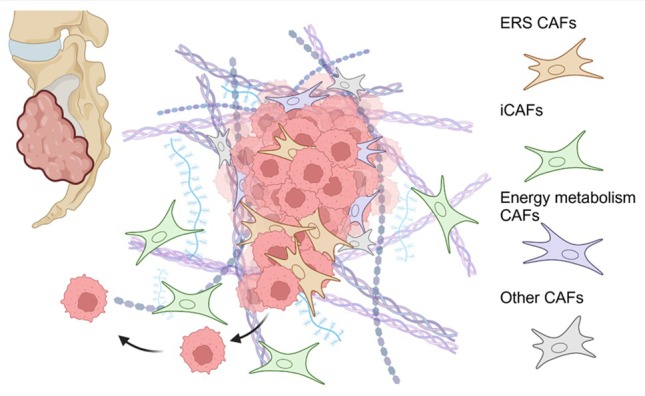 Figure 1
Figure 1| CAF subtypes | Selected markers | Biological functions/notes | Samples/cancer type | References |
|---|---|---|---|---|
|
4 CAF clusters: ‐ iCAFs (inflammatory) ‐ Energy metabolism enriched CAFs ‐ Cell junction enriched CAFs ‐ ECM CAFs |
All clusters iCAF: | iCAFs: tumour invasion, poor prognosis. Localised distant from cancer cells. |
7 primary sacral chordomas 4 nucleus pulposus | Zheng 2025 [ |
|
4 CAF clusters: ‐ myCAFs (myofibroblast) ‐ ECM CAFs ‐ Ribosomal CAFs ‐ Fibroblast‐like |
myCAF: ACTA2, ECM CAF:
| FN1, collagen and TGFβ interactions between CAFs and cancer cells; EMT/TGFβ pathway targeted. |
6 primary skull base chordomas | Zhang 2022 [ |
|
6 CAF clusters: ‐ mCAFs – ECM/matrix ‐ iCAFs ‐ bone CAFs ‐ vascular CAFs (2 clusters) ‐ EMT CAFs |
All clusters: ACTA2, Col1a2, PDGFRB mCAFs: collagens iCAFs: IL1RL1, CCL3, CCL4l2, CCL8 |
20% of cells are fibroblasts. TGFβ most prominent pathway driving invasion and immune suppression. |
6 primary chordomas: ‐ 2 skull base ‐ 3 vertebrae ‐ 1 sacrum | Duan 2022 [ |
|
4 CAF clusters ‐ ERS CAFs ‐ Energy metabolism enriched CAFs ‐ 2 rare CAF clusters |
All clusters: POSTN, ACTA2, FAP, PDGFRA ERS CAFs: ERS and Hypoxia signature genes, HSPs, VEGFA |
Endoplasmic reticulum Stress (ERS) CAFs associated with recurrence, invasion, poor prognosis. ERS CAFs associate closely with cancer cells. |
3 primary chordomas 3 recurrent chordomas 3 nucleus pulposus |
Zhang 2024 [ |
- —Medical Research Council 10.13039/501100000265
- —Cancer Research UK 10.13039/501100000289
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBone Tumor Diagnosis and Treatments · Sarcoma Diagnosis and Treatment · Oral and Maxillofacial Pathology
Chordomas are rare mesenchymal tumours that develop in the base of the skull, spine, or sacrum. They are widely understood to arise from remnants of the embryonic notochord, based on anatomic, histological, and molecular features. Despite their mesenchymal nature, single‐cell (sc) transcriptional analyses indicate retention of epithelial markers in malignant cell populations, expressed alongside mesenchymal and epithelial‐mesenchymal transition (EMT) markers [1, 2]. Cancer‐stem cell populations have also been described in multiple studies [1, 2]. Chordomas are typically characterised by brachyury (TBXT) dependence, indicative of their notochord origins, and high levels of TGFβ pathway activation. Chordomas contain numerous noncancerous stromal cells, dominated by subsets of heterogeneous cancer‐associated fibroblasts (CAFs) and diverse immune populations.
In a recent issue of this journal, Zheng and Guo [3] use scRNA sequencing to describe the presence of an abundant subset of inflammatory CAFs (iCAFs), which are associated with chordoma invasion and poor outcome (Figure 1). While the majority of CAF populations in solid tumours express a variety of extracellular matrix (ECM) and myofibroblast markers, iCAFs are distinguished by their abundant expression of inflammatory genes, including cytokines, chemokines, and inflammatory signalling components. Spatial transcriptomic and immunofluorescence studies indicate that these iCAFs are largely distal to malignant cells and associate with high levels of immune infiltrate, as has been described for iCAF populations in other solid tumours [4]. The authors propose that iCAFs drive tumour progression through various mechanisms, predominated by reciprocal paracrine interactions with immune cells. Using a powerful multiplexed quantitative immunofluorescence (QIF) dataset from 116 patients, with associated 10‐year outcome data, the authors show that high iCAF density associates with both clinicopathological features, including local invasion and adverse outcome; this is corroborated in bulk RNA‐seq datasets using an iCAF gene‐signature scoring system.
Causation versus correlation for distinct CAF subtypes
Evidence for iCAFs populations, alongside myofibroblast CAFs and other subtypes, have been described in many solid tumour types, and association of inflammatory tumour signatures with poor outcome has been variously reported (reviewed in ref. [4]). However, there remain open questions about causal versus correlative relationships between distinct CAF subtypes and disease outcome. While iCAFs may be associated with sites of invasion, their phenotype could be driven by invasion and inflammation rather than playing a causative role. Further, co‐occurrence of multiple CAF types can make attribution of causation problematic. Indeed, a previous scRNA‐seq analysis of chordoma from the lead author of the current study helped define a distinct endoplasmic reticulum stress‐related CAF (ERS‐CAF) subpopulation, which also predicts invasion and poor outcome [2]. Critically, implication of this ERS CAF subtype in tumour progression, by examining transcriptomic signatures or through immunofluorescence, used the same tumour cohorts as the current study. In contrast to the iCAFs described here, ERS CAFs associate closely with malignant cells, where they are proposed to promote malignant tumour cell behaviour directly, through paracrine growth‐factor secretion, ECM modulation, and adhesion signalling (Figure 1). In stroma‐rich tumours, the abundance of distinct CAF subtypes is strongly correlated; myofibroblast CAF‐rich tumours tend to also be rich in iCAFs and immune cells. Which of these characteristics is critical for defining outcome is therefore difficult to untangle. Indeed, the current authors previously reported that chordoma outcome could be predicted by the tumour‐stroma ratio, without qualitative assessment of stromal content [5]. These parallel studies beg the question as to which CAFs are drivers and which passengers on the road to malignant progression.
Clues may be found in CAF origins and abundance across the disease time course. In the two separate CAF studies described above, chordoma CAF populations were compared with CAFs from nucleus pulposus samples, and little overlap was seen between the normal tissue resident fibroblasts and the chordoma‐associated CAFs. Pseudotime analysis suggests that all CAF subtypes in chordoma differentiate from resident fibroblasts, which progressively evolve into activated myofibroblast CAF populations, from which iCAFs eventually emerge. This concurs with powerful longitudinal studies in other cancers, including pancreatic cancer [6]. To complicate matters, myofibroblast ERS CAFs were almost exclusively found in recurrent chordomas, which might implicate prior therapy as a factor driving the ERS phenotype, in addition to endogenous signals. When it comes to attribution of adverse outcome to individual CAF subtypes, the picture remains unclear, as both ERS and iCAFs arise as tumours progress and both separately associate with invasion (Figure 1).
To address causation in the current study, in vivo data are presented where chordoma isolated iCAFs are shown to promote malignant behaviours in vitro and support tumour growth in a subcutaneous xenograft model; normal fibroblasts are used as a control in these studies. While encouraging, caution must be applied here. Co‐culture of malignant cells with CAF models has been widely shown to confer survival advantage and promote growth and invasive behaviour in many cancer models both in vitro and in vivo, but this has not always translated to appropriate genetic models. Perhaps lessons can be learned here from pancreatic ductal adenocarcinoma (PDAC), where targeting CAFs and the fibrotic microenvironment has been the subject of extensive work. Numerous studies have indicated that CAFs, derived from pancreatic stellate cells and resident fibroblasts, play diverse tumour supporting roles, including promotion of invasion and suppression of antitumour immunity. Surprisingly, however, ablation of α‐SMA expressing CAFs in spontaneously developing pancreatic tumours resulted in more aggressive metastatic disease [7]; this has been corroborated in other related pancreatic cancer models, with consensus suggesting that loss of myofibroblast CAFs can result in more inflammatory and aggressive tumours [4, 8]. Attributing causative functions to specific subsets of CAFs thus requires a more nuanced approach. As a useful example, Dominguez et al identified a minor subset of LRCC15+ CAFs in PDAC, which arise during late pancreatic tumour development [6]; specific targeting of this subset in vivo in a pancreatic tumour model (KPR; Kras ^ LSL.G12D/wt ^ ;p16/p19 ^ fl/wt ^ ;p53 ^ LSL.R270H/wt ^ ;Pdx1.Cre) was effective at limiting tumour growth, while also promoting antitumour immunity and checkpoint inhibitor response [9]. Informatively, the aggressive inflammatory tumours observed with the deletion of α‐SMA‐positive CAFs also enhanced tumour response to anti‐CTLA4 checkpoint blockade [7]. Such studies highlight the need to separate correlation from causation, which will be critical when considering therapeutic targeting of the tumour stroma.
Can targeting the stroma in chordoma provide clinical benefit?
To date, at least four separate studies have reported the single‐cell transcriptomic landscape of chordoma, alongside immune‐cell focussed studies (Table 1) [1, 2, 10, 11]. All studies concur on the identification of TBXT‐expressing cancer cells exhibiting high epithelial‐mesenchymal signatures and TGFβ pathway activation. Cancer‐stem cells and a highly complex immune infiltrate are also common features. Abundant fibroblast populations are identified in each of these datasets, although little effort has been made to cross‐compare the transcriptomic data. A more comprehensive description of the evolving CAF landscape during malignant progression, therapy response and disease relapse would provide important context for the development of stromal targeting strategies. As an example, the ERS CAFs described by Zhang et al are almost exclusively found in relapsed chordoma [2]. Relapse occurs in cases refractory to surgery and radiotherapy, where immunotherapy is emerging as a promising option. To what degree would ERS CAFs suppress antitumour immunity and would targeting these cells be useful? In contrast, iCAFs are shown to appear late in poor prognosis, primary invasive disease, and may play an important role in sculpting the immune infiltrate, with implications for immunotherapy responses. Combining data from multiple studies will be required to better understand commonalties and heterogeneity between CAF subtypes identified in chordoma.
Chordoma is a clinically important but relatively rare disease. It may therefore also be useful to cross‐compare CAF subsets with those arising in other mesenchymal bone tumours. No doubt the clusters identified in chordomas, and other related tumours, would share significant overlap, and this would allow advances in clinical trials in related tumour types to be more readily translated to chordoma. As in other cancers, the need to identify discriminatory biomarkers to define functionally distinct CAF types remains critical; the chordoma studies published here can support this endeavour. The rapidly advancing technology for examining spatial transcriptomics at single‐cell resolution also provides an increasing opportunity to explore the local relationships between these distinct stromal populations, the malignant components, and the immune landscape. Ultimately, this has the potential to define biomarkers to identify, and even target, cell types of therapeutic interest. A framework for integrating single‐cell and spatially resolved data from diverse studies will, however, be necessary to better define mechanisms of tumour progression and therapeutic vulnerabilities.
Author contributions statement
AJMC and JCH conceived and wrote the article, compiled the table, and designed the figure.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Zhang Q , Fei L , Han R , et al. Single‐cell transcriptome reveals cellular hierarchies and guides p‐EMT‐targeted trial in skull base chordoma. Cell Discov 2022; 8: 94.36127333 10.1038/s 41421-022-00459-2PMC 9489773 · doi ↗ · pubmed ↗
- 2Zhang TL , Xia C , Zheng BW , et al. Integrating single‐cell and spatial transcriptomics reveals endoplasmic reticulum stress‐related CAF subpopulations associated with chordoma progression. Neuro Oncol 2024; 26: 295–308.37772937 10.1093/neuonc/noad 173PMC 10836767 · doi ↗ · pubmed ↗
- 3Zheng BW , Guo W . Multi‐omics analysis unveils the role of inflammatory cancer‐associated fibroblasts in chordoma progression. J Pathol 2025; 265: 69–83.39611243 10.1002/path.6369 · doi ↗ · pubmed ↗
- 4Menezes S , Okail MH , Jalil SMA , et al. Cancer‐associated fibroblasts in pancreatic cancer: new subtypes, new markers, new targets. J Pathol 2022; 257: 526–544.35533046 10.1002/path.5926 PMC 9327514 · doi ↗ · pubmed ↗
- 5Zou MX , Zheng BW , Liu FS , et al. The relationship between tumor‐stroma ratio, the immune microenvironment, and survival in patients with spinal chordoma. Neurosurgery 2019; 85: E 1095–E 1110.31501892 10.1093/neuros/nyz 333 · doi ↗ · pubmed ↗
- 6Dominguez CX , Müller S , Keerthivasan S , et al. Single‐cell RNA sequencing reveals stromal evolution into LRRC 15+ myofibroblasts as a determinant of patient response to cancer immunotherapy. Cancer Discov 2020; 10: 232–253.31699795 10.1158/2159-8290.CD-19-0644 · doi ↗ · pubmed ↗
- 7Özdemir BC , Pentcheva‐Hoang T , Carstens JL , et al. Depletion of carcinoma‐associated fibroblasts and fibrosis induces immunosuppression and accelerates pancreas cancer with reduced survival. Cancer Cell 2014; 25: 719–734.24856586 10.1016/j.ccr.2014.04.005PMC 4180632 · doi ↗ · pubmed ↗
- 8Murray ER , Menezes S , Henry JC , et al. Disruption of pancreatic stellate cell myofibroblast phenotype promotes pancreatic tumor invasion. Cell Rep 2022; 38: 110227.35081338 10.1016/j.celrep.2021.110227 PMC 8810397 · doi ↗ · pubmed ↗