Metagenome-assembled genomes of acetogenic enrichment obtained from deep subsurface archean granitoids of Koyna Seismogenic Zone, Koyna, India
Rajendra Prasad Sahu, Pinaki Sar

TL;DR
This study presents 14 genomes of acetogenic bacteria from deep underground rocks in India, providing insights into life in extreme environments.
Contribution
The paper provides new metagenome-assembled genomes of acetogens from the deep subsurface of India.
Findings
14 MAGs of acetogenic bacteria were assembled from deep granitoids in the Koyna Seismogenic Zone.
The genomes include species like Thermoanaerobacter pseudethanolicus and Caldanaerobacter subterraneus.
These findings enhance understanding of the deep biosphere's microbial diversity.
Abstract
We report 14 metagenome-assembled genomes (MAGs) of acetogenic bacteria (acetogens) enriched from deep (1,679–2,912 m below surface), hot (55°C–74 °C) granitoids of the Koyna Seismogenic Zone, India. These MAGs include Thermoanaerobacter pseudethanolicus, Exiguobacterium alkaliphilum, Moorella humiferrea, Caldanaerobacter subterraneus, etc. The study allowed access to the genomes of deep biosphere acetogens.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
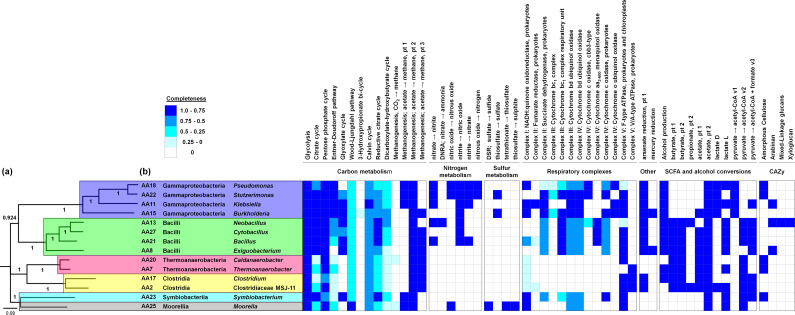 Fig 1
Fig 1| Mag id | Taxonomy (as per GTDBtk) | GeneBank accession | Completeness (%) | Contamination (%) | Genome coverage (x) | GC (%) | Size (Mb) | No. of contig | N50 (Mb) | Total CDS | RNA |
|---|---|---|---|---|---|---|---|---|---|---|---|
| AA2 | Bacteria; Bacillota; Clostridia; Clostridiales; Clostridiaceae; MSJ-11 |
| 100 | 1.61 | 177.34 | 30.3 | 4.06 | 33 | 0.27 | 3,990 | 43 |
| Bacteria; Bacillota; Thermoanaerobacteria; Thermoanaerobacterales; Thermoanaerobacteraceae; |
| 97.6 | 0 | 799.97 | 34.4 | 2.26 | 63 | 0.05 | 2,323 | 58 | |
| AA8 | Bacteria; Bacillota; Bacilli; Exiguobacterales; Exiguobacteraceae; |
| 98.34 | 0 | 164.86 | 52.8 | 2.9 | 48 | 0.11 | 2,950 | 31 |
| AA11 | Bacteria; Pseudomonadota; Gammaproteobacteria; Enterobacterales; Enterobacteriaceae; |
| 99.37 | 0.27 | 4.1 | 55.2 | 4.89 | 154 | 0.06 | 4,671 | 46 |
| AA13 | Bacteria; Bacillota; Bacilli; Bacillales; DSM-18226; |
| 91.81 | 3.38 | 11.06 | 38.7 | 4.8 | 87 | 0.11 | 4,686 | 33 |
| AA15 | Bacteria; Pseudomonadota; Gammaproteobacteria; Burkholderiales; Burkholderiaceae; |
| 100 | 0.42 | 20.16 | 66.2 | 8.66 | 83 | 0.17 | 7,845 | 59 |
| Bacteria; Bacillota; Clostridia; Clostridiales; Clostridiaceae; |
| 98.46 | 1.9 | 8.6 | 30 | 3.67 | 65 | 0.09 | 3,451 | 37 | |
| AA18 | Bacteria; Pseudomonadota; Gammaproteobacteria; Pseudomonadales; Pseudomonadaceae; |
| 90.78 | 0.71 | 8 | 66.9 | 6.17 | 365 | 0.03 | 6,005 | 16 |
| Bacteria; Bacillota; Thermoanaerobacteria; Thermoanaerobacterales; Thermoanaerobacteraceae; |
| 97.2 | 1.13 | 37.85 | 37.7 | 2.27 | 139 | 0.02 | 2,341 | 41 | |
| AA21 | Bacteria; Bacillota; Bacilli; Bacillales; Bacillaceae; |
| 98.61 | 3.86 | 13.63 | 35.1 | 5.28 | 34 | 0.28 | 5,447 | 21 |
| AA22 | Bacteria; Pseudomonadota; Gammaproteobacteria; Pseudomonadales; Pseudomonadaceae; |
| 99.53 | 0.14 | 23.13 | 64.1 | 4.38 | 74 | 0.15 | 4,111 | 51 |
| AA23 | Bacteria; Bacillota; Symbiobacteriia; Symbiobacteriales; Symbiobacteriaceae; |
| 98.01 | 0 | 4.87 | 68.4 | 3.66 | 145 | 0.05 | 3,383 | 46 |
| Bacteria; Bacillota; Moorellia; Moorellales; Moorellaceae; |
| 99.48 | 1.36 | 3.84 | 54 | 2.5 | 205 | 0.03 | 2,739 | 49 | |
| AA27 | Bacteria; Bacillota; Bacilli; Bacillales; DSM-18226; |
| 97.53 | 1.14 | 16.77 | 39.1 | 4.01 | 64 | 0.1 | 3,901 | 20 |
- —Ministry of Earth Sciences, India
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenomics and Phylogenetic Studies · Plant Pathogenic Bacteria Studies · Plant Disease Resistance and Genetics
ANNOUNCEMENT
Acetogens are important members of energy-starved deep biosphere (1). Despite their critical ecological roles in driving primary production in deep biosphere through the Wood–Ljungdahl pathway (2) and biotechnological potential in C1 gas valorization (3), the genomes of these organisms remain understudied. Here, we present 14 bacterial MAGs enriched from deep, hot, Archean basement granitoids.
Seven rock cores spanning a depth range of 1,679–2,912 m and temperature gradient 50°C–74°C were sampled through 3,000 m deep Koyna pilot-borehole (Koyna seismogenic zone, Koyna, 17°17′57.27″ N, 73°44′19.07″ E), India. Standard procedures were followed to minimize samples’ contamination (4, 5). Details of drilling, contamination-free rock sample collection, etc. were mentioned elsewhere (6, 7). Acetogens were enriched by adding rock powder into acetogenic medium (2 g/100 mL) in serum vial supplemented with H_2_ + CO_2_ (80:20 v/v) in the headspace (8). An A35 Don Whitely anaerobic workstation was used. Considering the in situ condition, three different incubation temperatures (50°C, 60°C, and 70°C) were used for the enrichment (details of the sampling depth, in situ and incubation temperatures of rock samples are provided as Table S1 at https://doi.org/10.6084/m9.figshare.28215026).
Following 6 months’ incubation, 10 mL culture from each set was centrifuged (14,000×g, 30 min), cell pellet was used for total DNA extraction using QIAGEN Powersoil DNA isolation kit. Pair-end library was synthesized using Illumina DNA Prep, (M) Tagmentation kit followed by shotgun metagenome sequencing using Illumina NovaSeq 6000 (2 × 101 bp chemistry). Default parameters were used for sequencing and all software unless otherwise stated. Sequenced reads were demultiplexed (Illumina bcl2fastq [2.20]) and following adaptor removal (Skewer v0.2.2 [9]), their quality was analyzed (FastQC v0.12.1 [10]) and trimmed (Trimmomatic v0.39 [11]). Human contamination reads were removed (bowtie2 v2.5.3 together with samtools v1.7 and human host genome GRCh38 bowtie2 database [12, 13]). Quality filtered reads were assembled using MEGAHIT-1.2.9 (14), followed by binning through METABAT2 v1.2.9 (15), CONCOCT v1.1.0 (16), and MaxBin2 v2.2.7 (17). Resulting bins were consolidated and refined using Bin_refinement module (metaWRAP v1.3 [18]). GC%, completeness, and contamination percentage were determined (checkM v1.2.2 [19]), followed by genome coverage calculation (quant_bins module of metaWRAP v1.3 [18]). High-quality bins [>90% completeness, <5% contamination, total contigs < 500, N50 >20 kb [20, 21]) were taxonomically affiliated through GTDB-Tk v2.1.1 (22), and resulting alignments were used to build phylogenetic tree using FigTree v1.4.4 (http://tree.bio.ed.ac.uk/software/figtree/). Bins were annotated with PGAP v6.7 and kb_DRAM v0.1.2 (23, 24).
During this study 113.62 million quality filtered reads were assembled into 29,442 contigs. These were binned into 14 high-quality MAGs with average size and GC content 4.25 Mb and 48.06%, respectively (Table 1). MAGs consisted of 33–365 contigs (average N50 value 108.57 kb) with coverage varied between 3.84x and 799.97x. Annotation of MAGs showed presence of 2,323–7,845 number of coding DNA sequences (CDS). Taxonomic diversity and functional potential of the MAGs are presented in Fig. 1.
(a) Maximum likelihood tree of 14 metagenome-assembled genomes (MAGs). The phylogenetic tree was constructed using a concatenated set of 120 conserved bacterial marker genes from each of these MAGs using GTDB-Tk “classify” function and FigTree v1.4.4.. (b) Heatmap displays the pathways fully or partially present in each MAG, as predicted by kb_DRAM (24).
These MAGs provide access to the genomes of deep rocky-biosphere acetogens allowing us to gain insights into their significance in functioning and adaptation of deep life.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Beaver RC, Neufeld JD. 2024. Microbial ecology of the deep terrestrial subsurface. ISME J 18:wrae 091. doi:10.1093/ismejo/wrae 09138780093 PMC 11170664 · doi ↗ · pubmed ↗
- 2Momper L, Casar CP, Osburn MR. 2023. A metagenomic view of novel microbial and metabolic diversity found within the deep terrestrial biosphere at De MMO: A microbial observatory in South Dakota, USA. Environ Microbiol 25:3719–3737. doi:10.1111/1462-2920.1654337964716 · doi ↗ · pubmed ↗
- 3Bae J, Park C, Jung H, Jin S, Cho B-K. 2024. Harnessing acetogenic bacteria for one-carbon valorization toward sustainable chemical production. RSC Chem Biol 5:812–832. doi:10.1039/d 4cb 00099 d 39211478 PMC 11353040 · doi ↗ · pubmed ↗
- 4Colwell FS, Boyd S, Delwiche ME, Reed DW, Phelps TJ, Newby DT. 2008. Estimates of biogenic methane production rates in deep marine sediments at hydrate ridge, cascadia margin. Appl Environ Microbiol 74:3444–3452. doi:10.1128/AEM.02114-0718344348 PMC 2423016 · doi ↗ · pubmed ↗
- 5Kieft TL, Phelps TJ, Fredrickson JK. 2007. Drilling, coring, and sampling subsurface environments. Man Environ Microbiol:799–817.
- 6Sahu RP, Kazy SK, Bose H, Mandal S, Dutta A, Saha A, Roy S, Dutta Gupta S, Mukherjee A, Sar P. 2022. Microbial diversity and function in crystalline basement beneath the deccan traps explored in a 3 km borehole at Koyna, western India. Environ Microbiol 24:2837–2853. doi:10.1111/1462-2920.1586734897962 · doi ↗ · pubmed ↗
- 7Roy S. 2017. Scientific drilling in Koyna region, Maharashtra. Curr Sci 112:2181.
- 8Hallbeck L, Pedersen K. 2008. Characterization of microbial processes in deep aquifers of the fennoscandian shield. Appl Geochem 23:1796–1819. doi:10.1016/j.apgeochem.2008.02.012 · doi ↗