Bioprospection of Hura crepitans metabolites against oxidative stress and inflammation: An in vitro and in silico exploration
Yu-Cheng Kuo, Bashir Lawal, Halimat Yusuf Lukman, Lung-Ching Chen, Sheng-Liang Huang, Yi-Fong Chen, Adewale O. Fadaka, Femi Olawale, Ayo Olasupo, Olabode T Ajenifujah, Dalia Fouad, Marios Papadakis, Gaber El-Saber Batiha, Saheed Sabiu, Alexander T.H. Wu, Hsu-Shan Huang

TL;DR
This study explores the antioxidant and anti-inflammatory properties of Hura crepitans, identifying key compounds like rutin that show promise for treating oxidative stress and inflammation.
Contribution
The study combines in vitro and in silico methods to uncover the mechanisms of Hura crepitans' bioactive compounds against oxidative and inflammatory targets.
Findings
The crude methanolic extract of H. crepitans showed strong antioxidant activity comparable to ascorbic acid.
Rutin was identified as the most abundant and stable compound in interactions with oxidative and inflammatory targets.
Computational analyses confirmed the safety and binding stability of the identified compounds.
Abstract
Background: Despite the recognized therapeutic potential of Hura crepitans, its mechanistic antioxidant and anti-inflammatory actions remain underexplored. Methods: This study investigates the inhibitory effects, binding stability, and interactions of metabolites from H. crepitans on oxidative and inflammatory biomarkers/targets using in vitro analyses and molecular dynamics (MD) simulations. Results: In vitro experiments revealed significant dose-dependent antioxidant and anti-inflammatory activities. The crude methanolic extract (CMEHC) showed notable half-maximal inhibitory concentration (IC50) values for antioxidant assays, such as diphenyl picrylhydrazine (45.51 µg/mL) and ferric-reducing power (10.86 µg/mL), with comparable performance to standard ascorbic acid. Anti-inflammatory activities, including protein denaturation, proteinase inhibition, and membrane stabilization,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
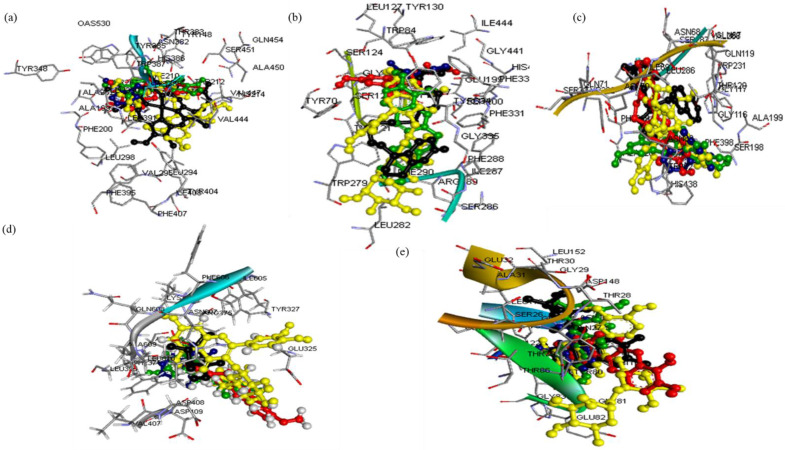 Figure 1
Figure 1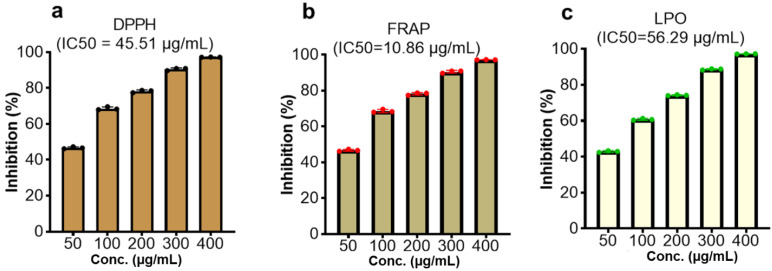 Figure 2
Figure 2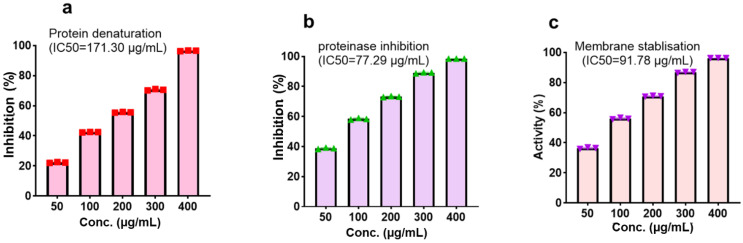 Figure 3
Figure 3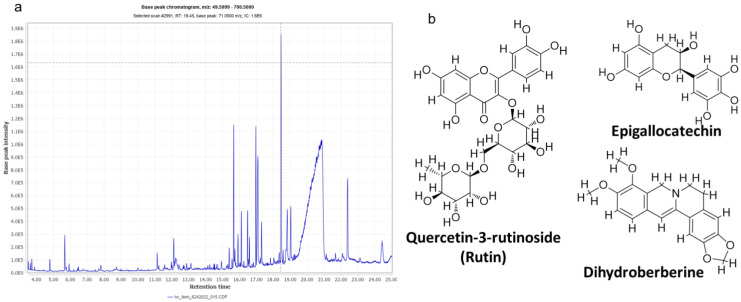 Figure 4
Figure 4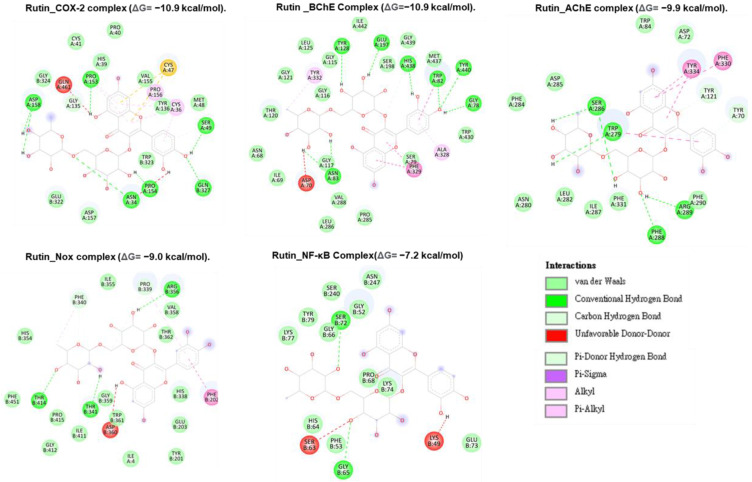 Figure 5
Figure 5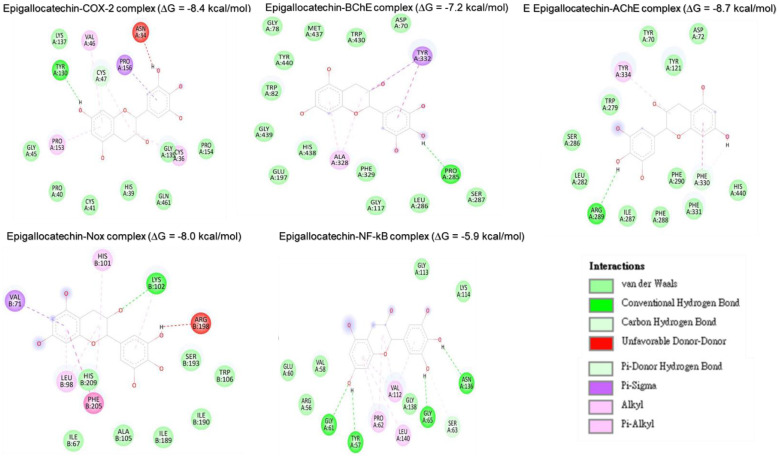 Figure 6
Figure 6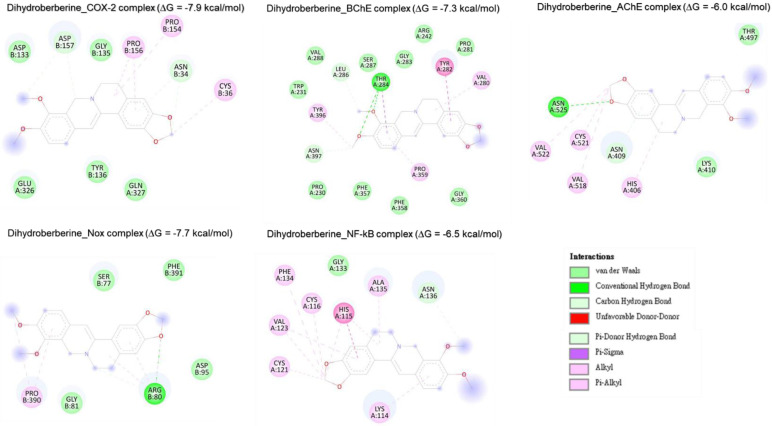 Figure 7
Figure 7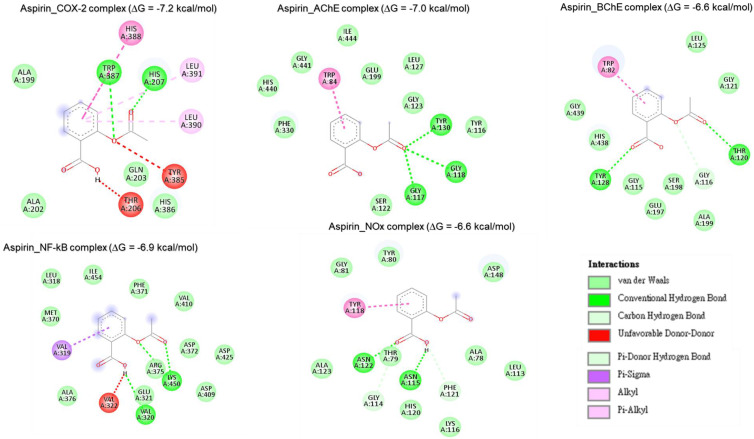 Figure 8
Figure 8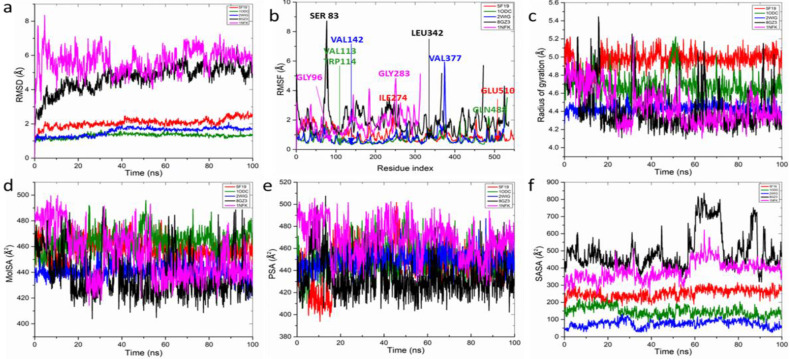 Figure 9
Figure 9 Figure 10
Figure 10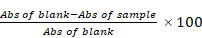 Figure 11
Figure 11Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNatural product bioactivities and synthesis
Introduction
Vascular and organ abnormalities stemming from several reaction pathways that lead to free radical production and inflammatory responses are central to the pathogenesis of metabolic dysfunctions, including diabetes, cancer, cardiovascular, liver, kidney, neurodegenerative, and pulmonary diseases contributing significantly to global morbidity and mortality 1. In diabetes, oxidative stress and inflammation impair insulin signaling and secretion 2, while in cardiovascular diseases, oxidative stress and chronic inflammation exacerbate vascular endothelial dysfunction and atherosclerosis 3. Reactive oxygen and nitrogen species, such as superoxide and nitric oxide, activate pathways like cyclooxygenase and transcription factors such as NF-κB, driving disease progression 4. Addressing these pathways with effective antioxidant and anti-inflammatory agents is critical for therapeutic intervention.
Phenolics, naturally occurring compounds found in plants such as vegetables, fruits, and cereals, are recognized for their potent antioxidant and anti-inflammatory properties, with potential benefits for managing diabetes, cancer, and cardiovascular diseases 5. Although synthetic antioxidants like ascorbic acid and endogenous agents such as superoxide dismutase play a role in mitigating oxidative damage, limitations in cost, availability, and adverse effects often hinder their use. Similarly, non-steroidal anti-inflammatory drugs (NSAIDs) like aspirin are effective but not without risks 4. Consequently, there is a growing interest in plant-derived bioactive compounds as safer and more accessible alternatives.
Hura crepitans L., commonly known as the "sandbox tree" or "monkey dinner bell," is a versatile member of the Euphorbiaceae family, valued for its traditional and medicinal uses across tropical regions of the Americas 7,8. This tree has long been recognized for its therapeutic properties, including its use as a laxative and remedies for inflammation, microbial infections, liver damage, leprosy, and as an emetic 9,10. Modern studies have validated these traditional applications, revealing a rich profile of bioactive compounds such as flavonoids, phenolic acids, alkaloids, tannins, carotenoids, terpenes, fatty acids, and essential amino acids like methionine and lysine 15-19. Notably, compounds such as rutin, myricetin, ferulic acid, and daphnane diterpenes contribute significantly to its pharmacological potential 9,20.
Research has demonstrated various pharmacological effects of H. crepitans, including hepatoprotective, antihypertensive, antidiabetic, antioxidant, and anti-inflammatory activities 16,21-25. However, while these findings underscore its therapeutic promise, the mechanisms underlying its antioxidant and anti-inflammatory effects remain poorly understood, particularly through computational approaches. Such methods, in combination with in vitro studies, have proven essential in modern drug discovery, providing efficient ways to explore the pharmacodynamics of medicinal plants and translate traditional knowledge into therapeutic innovations 26-32. Consequently, this study investigated the in vitro antioxidant and anti-inflammatory of H. crepitans and computationally explored its mechanism of action, setting the stage for future drug development endeavors.
Materials and Methods
Collection and preparation of the extract
Hura crepitans plant was collected within Minna metropolis, Niger state, Nigeria, identified and authenticated at the herbarium unit of the Federal University of Technology (FUT), Minna, Nigeria where a voucher number was deposited. Crude methanolic extract of H. crepitans (CMEHC) was obtained by firstly rinsing dirt and dust off the plant with clean water and air-dried until a constant dry weight was obtained. The dried plant material was then ground into a fine powder and extracted with methanol (1:5) for 72 h with intermittent shaking for complete extraction. Subsequently, the mixture was filtered using Whatman filter paper (No. 1), and the filtrate concentrated in a rotary evaporator. The CMEHC obtained was then stored in a refrigerator (4°C) for further use.
In vitro antioxidant analysis
The antioxidant potential of the extract was evaluated by analyzing its DPPH radical-scavenging, ferric-reducing antioxidant power (FRAP), and lipid peroxidation (LPO) inhibitory potentials. For the DPPH assay, varying concentrations of CMEHC (50-400 µg/mL) and reference standard (ascorbic acid) were mixed with a DPPH solution in methanol and allowed to react in darkness for 45 min. The reduction in absorbance indicating DPPH scavenging activity was measured at 517 nm 33. In the FRAP assay, varying concentrations of CMEHC were reacted with a mixture containing potassium hexacyanoferrate (III) and trichloroacetic acid in sodium phosphate buffer. Following centrifugation, ferric chloride was introduced to the supernatant for color development, and absorbance was recorded at 700 nm 34. The LPO analysis involved treating a mixture of egg homogenate and FeSO_4_ with CMEHC, followed by incubation and subsequent addition of acetic acid and an acid-reactive substance. After heating, butanol was added, the mixture was centrifuged, and absorbance was read at 532 nm 35. The percentage inhibition was calculated using the equation displayed, while the half-maximal inhibitory concentration (IC_50_) was determined using linear regression.
In vitro anti-inflammatory analysis
The anti-inflammatory potential of the extract was investigated for its human red blood cell (RBC) membrane stability, protein denaturation inhibition, and proteinase inhibitory capabilities. For the RBC membrane stability assay, freshly collected human plasma was processed and mixed with reference standard aspirin or varying CMEHC concentrations in a 10% red blood suspension. Following incubation at 56 °C for 30 min and centrifugation, the absorbance of the supernatant was measured at 560 nm 36. The protein denaturation inhibition assay involved mixing CMEHC or aspirin with a 1% bovine serum albumin (BSA) solution. The mixture was heated at 55 °C for 30 min and allowed to cool to observe turbidity 37. For the proteinase inhibitory assay, a reaction mixture containing trypsin, Tris-HCl buffer, and CMEHC was incubated, followed by the addition of casein and further incubation. The reaction was terminated with perchloric acid, and after centrifugation, the absorbance of the supernatant was measured at 210 nm against a Tris-HCl buffer 38. The percentage inhibition was calculated using the equation displayed, while the half-maximal inhibitory concentration (IC_50_) was determined using linear regression.
Characterization of the CMEHC with a liquid chromatographic mass spectrometric (LC-MS) analysis
The LC-MS analysis of the crude methanolic extract of Hura crepitans (CMEHC) was done using a Shimadzu LC-MS-8040 ultrafast mass spectrometer, equipped with a Shim-pack FC-ODS analytical column. The analysis involved a dual mobile phase system: mobile phase A with 5 mmol/L ammonium acetate in water and mobile phase B with 5 mmol/L ammonium acetate in methanol. The gradient elution program varied the concentration of phase B from 15% to 95% over a 40-minute period, maintaining a flow rate of 0.2 mL/min and a column temperature of 40 °C. Mass spectrometry conditions included a +4.5 kV probe voltage in ESI-positive mode, nebulizing gas flow of 1.5 L/min, drying gas of 10 L/min, with temperatures set at 250 °C for the drying gas and 400 °C for the heat block. Scans ranged from 100 to 1000 m/z at a speed of 5000 u/s in the positive ionization mode, with spectra monitored using Shimadzu Lab Solution software over a 0.00-50.00 retention time frame. Data analysis was conducted in stages, starting with mass detection and chromatogram construction, followed by peak deconvolution and filtering using mzmine software (version 2.53) for compound identification, after exporting the results in CDF format for enhanced peak analysis efficiency 39,40.
In silico pharmacokinetics studies
The pharmacokinetics (PK), drug-likeness, medical, and physicochemical properties of the most abundant biologically active compounds of CMEHC were assessed using the ADMERLab, ADMETSar, and SWISSADME online databases 41.
Molecular docking and dynamics simulation
The three-dimensional (3D) structures of the ligands (rutin, dihydroberberine, and epigallocatechin) were acquired using the Avogadro molecular builder and visualization tool (version 1.XX), initially in mol2 format, and then converted to PDB format with the PyMOL Molecular Graphics System (version 1.2r3pre). Inflammatory hub targets [COX-2 (5F19), acetylcholinesterase (1H23), butyrylcholinesterase (5LKR), nuclear factor-κB [NF-κB (1NFK), and NADPH oxidase (NOx] were sourced from the Protein Data Bank (PDB) in PDB format 42. These targets underwent preparation for docking and saved as PDBQT files for compatibility 43-45. The docking process utilized Avogadro software, and the results visualized in both PyMOL and Discovery Studio Visualizer (version 19.1.0.18287, BIOVIA, San Diego, CA, USA) 44,46-48. Docking validation was conducted to prevent pseudo-positive binding by ensuring superimposition of the docked phenolic compounds at the target's active site as the native ligands with root mean square deviation (RMSD) of 0.5 Å for each target with the visualization done using Discovery Studio v21.1.0 23 (Figure 1).
Molecular dynamics (MD) simulations of the docked rutin complexes were executed using the Schrodinger suite (2020-2), specifically the Desmond software integrated within the system builder module of Maestro (version 12.4). These simulations spanned a 100 ns timeframe, adhering to methodologies detailed in prior studies 49,50. Post-simulation analyses including root-mean-square fluctuation (RMSF), root-mean-square deviation (RMSD), radius of gyration (rGyr), solvent-accessible surface area (SASA), changes in secondary structure, and hydrogen bond counts 51 were analyzed. The binding free energy of the complexes was calculated employing the molecular mechanics Poisson-Boltzmann surface area (MM-PBSA) method, analyzing 1000 trajectory files to ensure thorough assessment.
Data analysis
Data were analyzed using GraphPad Prism software. All values are expressed as the mean ± standard error of the mean (SEM) of three independent replicates (n = 3) to reflect the consistency and reliability of the experimental outcomes. The IC_50_, defined as the concentration required to achieve 50% inhibition of the targeted activity, was determined and compared between the extract and standard compounds.
Results
In vitro antioxidant and anti-inflammatory effects of the CMEHC
The evaluation of the CMEHC for its antioxidant and anti-inflammatory potential demonstrated a dose-dependent activity across the various assays. For antioxidant activity, the extract showed significant inhibition in the DPPH, FRAP, and lipid peroxidation (LPO) assays, with IC_50_ values of 45.51 µg/mL, 10.86 µg/mL, and 56.29 µg/mL (Figure 2), respectively while IC_50_ values of 20.90 µg/mL, 15.32 µg/mL, and 21.87 µg/mL, respectively, were recorded for ascorbic acid. The highest inhibition percentages for ascorbic acid were recorded at a concentration of 250 µg/mL, demonstrating values of 98.46 ± 0.15%, 98.13 ± 0.02%, and 98.69 ± 0.13% for each assay, respectively.
With regards to the anti-inflammatory activity, CMEHC exhibited dose-responsive effectiveness in inhibiting protein denaturation, proteinase inhibition, and stabilizing cell membranes, with IC_50_ values of 171.30 µg/mL, 77.29 µg/mL, and 91.78 µg/mL (Figure 3)., respectively. Aspirin had inhibition percentages of 96.86 ± 0.08%, 99.80 ± 0.01%, and 97.76 ± 0.15% at a concentration of 250 µg/mL, and presented IC_50_ values of 39.40 µg/mL, 8.15 µg/mL, and 10.12 µg/mL for the respective assays.
High-Performance LC-MS Analysis of the Bioactive Compounds in CMEHC
The high-performance LC-MS analysis of CMEHC identified five principal compounds, with their mass detection facilitated by mzmine software, applying a retention time (RT) tolerance of 0.01 min and a mass-to-charge ratio (m/z) tolerance of 0.02 or 5.0 ppm. These compounds were identified using comprehensive libraries and databases, presenting a sequence of increasing percentage areas: epigallocatechin, quercetin-3-rutinoside, dihydroberberine, 2-(3,4-dihydroxyphenyl)-5,7-dihydroxychromene-4-one, and hexadecanoic acid. The RT values observed were 25.234 min for epigallocatechin, 14.139 min for quercetin-3-rutinoside, 21.480 min for dihydroberberine, 26.433 min for 2-(3,4-dihydroxyphenyl)-5,7-dihydroxychromene-4-one, and 24.926 min for hexadecanoic acid. Quercetin-3-rutinoside exhibited the highest base peak value (m/z) of 61.11 ppm, followed by dihydroberberine (33.825 ppm), epigallocatechin (30.425 ppm), 2-(3,4-dihydroxyphenyl)-5,7-dihydroxychromene-4-one (28.435 ppm), and hexadecanoic acid (256.30 ppm) (Table 1). The chromatogram and structural elucidation of the phenolic compounds is represented (Figure 4a, b).
In silico analysis of the pharmacokinetics and drug-likeness properties of selected CMEHC compounds
The in-silico evaluation of three phenolic compounds from CMEHC—rutin, dihydroberberine, and epigallocatechin—focused on their pharmacokinetic (PK) profiles, drug-likeness, and physicochemical characteristics (Table 2). The analysis revealed that all compounds exhibited satisfactory bioavailability scores and intestinal absorption, except for rutin. Besides rutin, both dihydroberberine and epigallocatechin had molecular weights less than 500 g/mol and demonstrated synthetic accessibility scores below 400. The trio showed high plasma protein binding rates (over 80%) and extensive volume of distribution. Notably, dihydroberberine alone was identified as blood-brain barrier permeable. Despite these differences, all compounds were characterized by favorable half-lives and clearance rates, alongside minimal interaction with cytochrome P450 isoenzymes, either as inhibitors or substrates.
Molecular docking analysis reveals rutin as a potent inhibitor of neuronal and anti-inflammatory targets
Molecular docking studies were performed to evaluate the interaction between CMEHC compounds (rutin, dihydroberberine, and epigallocatechin) and key neuronal and inflammatory targets, including [cyclooxygenase (COX-2), acetylcholinesterase (AChE), butyrylcholinesterase (BChE), nuclear factor-kappa B (NF-κB), and NADPH oxidase] (Figures 5-7). The reference standard (aspirin) binding with the targets are shown in Figure 8. Except for AChE-dihydroberberine and NF-kB-epigallocatechin complexes with lower negative docking score that the standards, phenolic compounds in CMEHC presented higher negative docking scores than the standards when docked with the investigated targets (Table 3). Comparatively, the compounds exhibited strong binding affinities to the target proteins, with COX-2 emerging as the most vulnerable and NF-κB as the least (Table 3). Among the compounds, rutin displayed the most significant binding efficiency, with the lowest docking scores (ΔG) ranging from -7.2 to -10.9 kcal/mol, relative to the epigallocatechin (ΔG= -5.9 to -8.7 kcal/mol) and dihydroberberine (ΔG= -6.0 to -7.9 kcal/mol). Binding interactions between the targets and the compounds showed several interacting amino acids including H-bonding, alkyl interactions pi-interactions and several van der Waal forces with rutin again having more interacting amino acid residues with the targets (Table 4). The superior binding affinity of rutin was further confirmed by its higher MMGBSA scores (Table 5), ranging from -38.16 to -61.65 kcal/mol, compared to epigallocatechin (-20.30 to -34.60 kcal/mol) and dihydroberberine (-31.29 to -47.36 kcal/mol). These findings, corroborated by various docking models, pinpoint rutin as the leading candidate for targeting these targets. Consequently, rutin was chosen for further exploration through molecular dynamics (MD) simulation.
Molecular dynamics simulation of top ranked H. crepitans phenolics with anti-inflammatory proteins
The structural dynamics and interactions of rutin which demonstrated the lowest free binding energy when complexed with key anti-inflammatory targets were analyzed (Figure 9 and Table 6). The RMSD plot showed stable rutin-target patterns with AChE-rutin complex having fluctuated higher relative to the other complexes (Figure 9a) over the 100-ns simulation period, displaying a range of RMSD values: AChE (1.31±0.12) < BChE (1.55±0.24) < COX-2 (2.06±0.25) < NOx (4.68±0.76) < NF-kB (5.67±0.70). The RMSF plot varying fluctuation peak patterns in each target complex (Figure 9b), with mean values spanning 0.71±0.38 to 1.89±0.87 Å (Table 6). Notably, certain residues exhibited significant interactions with rutin, demonstrating minimal fluctuations, such as GLU510 in COX-2 and VAL113 and TRP114 in AChE, whereas others, like ILE274 in COX-2 and GLN488 in AChE, showed greater fluctuations due to lesser interactions (Table 6).
The rGyr plot demonstrated initial instability across all systems within the first 20 ns, however, formed stable complexes from 60 ns to the end of the simulation (Figure 9c) with slightly different mean values ranging from 4.41±0.22 to 4.96±0.09 Å^2^ for all the targets (Table 6). With regards, the molecular surface area (MolSA) and polar surface area (PSA), minimal fluctuating plots were observed in all the target-rutin bound complexes (Figure 9d, e) with NOx-rutin complex having the least mean values of 438.26±18.16, 433.79±17.39 Å^2^ among all the targets (Table 6). However, increasing fluctuating SASA was observed (Figure 9e) exhibiting significantly different mean values with NOx having the highest mean SASA value of 496.11±109.47 Å^2^ (Table 6).
Discussion
Oxidative stress and inflammation result in dysfunctions in lipid, nucleic acid, protein, and carbohydrate metabolism 52 with effects culminating in cellular/organ failure and subsequent morbidity and mortality risk 53. This is because the incessant assault of biological molecules by free radicals produced during oxidative stress and inflammatory responses are precursors to numerous diseases. The defense against these radicals is fortified by antioxidants, which mitigate their harmful effects 54. The myriads of therapeutic benefits of medicinal plants as alternative drug agents have been recognized and documented extensively, highlighting their pivotal role in combating various ailments through ages 55. This study elucidated the antioxidant and anti-inflammatory capabilities of Hura crepitans, employing a combination of in vitro and in silico methodologies.
Medicinal plants have historically been a cornerstone in the treatment of diverse health conditions, owing to their rich repository of phytochemicals that underpin their therapeutic efficacy 56. The in vitro antioxidant analysis of CMEHC showcased its potent DPPH-inhibitory effect, FRAP, and LPO, protein denaturation, proteinase and enhanced membrane stability. The observed lower IC_50_ values in the standards relative to the extract might be due to their pure and refined state which might influence their affordability and availability in contrast to herbal medications. The protective capabilities of plants against these pathways, as evidenced by their ability to counteract inflammation and inhibit protease activities, are crucial 13. Our findings align with these observations, as CMEHC inhibited protein denaturation, proteinase activity, and membrane destabilization effectively, supporting its potential as a dual-functional therapeutic agent, mirroring the in vivo anti-inflammatory and antidiabetic study of Lawal et al. 30.
Complementing these observations, Vassallo et al. 13 identified key compounds such as epigallocatechin in H. crepitans, with epigallocatechin showing promise in mitigating oxidative and endoplasmic reticular stress, among other biological activities 57. This aligns with our findings where CMEHC compounds not only demonstrated significant pharmacokinetic and drug-likeness properties but also indicated their suitability for oral therapeutic applications.
Consequent of the findings from the in vitro study, the potential anti-inflammatory mechanism of Hura crepitans metabolites were investigated for possible hit compounds. Additionally, delving into the molecular mechanisms through protein-ligand interactions, a critical component in myriad biological processes, offered further insights on the biological mechanism of action of metabolites 58. While virtual screening informs the pharmacokinetics, drug-likeness, synthetic ability, and toxicity profiles of a compound, molecular docking and MD simulation provides information on the binding fitness, and binding stability and interactions, respectively 59,60.
Naturally occurring antioxidants contribute significantly to the health-promoting properties of plants, capable of donating hydrogen atoms to neutralize the destabilizing effects of free radicals in reactions such as DPPH and LPO, besides catalyzing reduction processes 61,62. Due to the relationship between oxidative stress and inflammation 52, there is a high chance of a molecule to affects both conditions. On the premise of phenolics being potent antioxidant and anti-inflammatory agents 63, the identified phenolic compounds in the extract were subjected to in silico analysis. The three phenolic compounds demonstrated significant binding fitness and orientation with the targets better than the reference standard and further underscores the capability of H. crepitans modulatory effect on the investigated targets and as safer and more cost-effective alternative antioxidant and anti-inflammatory agents.
Although rutin demonstrated lesser pharmacokinetics, drug-likeness properties relative to the other two compounds, it presented greater binding fitness. Based on this, rutin was subsequently selected for MD simulations. Furthermore, reactions leading from free radical generation to inflammatory responses, mediated through mechanisms like RBC peroxidation and lysosomal membrane destabilization, underline the importance of anti-inflammatory agents 64-66. Thus, rutin binding interaction was analyzed with neuronal inflammatory targets including COX-2, AChE, BChE, NADPH oxidase, and NF-κB to solidify its anti-inflammatory potential, dictated by low binding energies and favorable non-covalent interactions observed in molecular docking studies. The free binding energy describes the binding affinity of a compound to the active site of a target with lower free binding energy indicating better binding affinity and pointer to the level of stability of the complex 67. The simulations hinted the stability and minimal conformational changes of rutin, especially notable in complexes with COX-2, AChE, and BChE, throughout the 100-ns simulation period.
Post dynamics parameters such as RMSD, RMSF, and rGyr describe stability of a complex in term of convergence, interaction of the compound in the binding pocket of the target, and complex compactness, respectively 68. With regards RMSD, the stability is inversely proportional to the binding stability 69. The RMSD analysis indicated that the complexes remained stable over the 100-ns simulation period with Nox- and NF-kB-rutin complexes having > 3.0 limit reported for a stable complex 70 indicative of lesser stability with rutin. The observed stability of the rutin-target bond complexes is consistent with the free binding energy for all the complexes where NOx with lower free binding energy had high mean RMSD and hence lesser stability. The flexibility of a protein's binding pocket upon binding of a compound suggests the stability of the complex formed relative to the important amino acids in the active site of the protein and 71. Notably, certain residues in the active site of the investigated targets exhibited reduced fluctuations suggestive of significant binding interactions with rutin, such as GLU510 in COX-2 and VAL113 and TRP114 in AChE, whereas others, like ILE274 in COX-2 and GLN488 in AChE, showed greater fluctuations due to lesser interactions. Finding from the RMSF values underscored the differential stability conferred by rutin binding, particularly enhancing the stability of AChE, BChE, and COX-2 complexes and its potential to modulate the targets differently.
The rGyr describes the compactness and active site stability of protein-ligand complex 72. Findings from this study revealed that the target-rutin complexes had initial instability across all systems within the first 20 ns, which gradually achieved stability, maintaining a constant state from 60 ns to the end of the simulation. However, the reduced lower mean rGyr of rutin-NOx suggests greater stability and hence the potential of rutin to form compact complex with NOx for modulatory effect. The molecular surface area (MolSA) describes interactions of a molecule with surrounding molecules and environment including its interaction with Vander waal surface and area open to steric hinderance 73. The polar surface area (PSA) describes molecules' exposed surface to polar, charged, or functional groups thus describing the hydrophilic nature of a molecule. The more polar a molecule is, the more difficult it transport across the membrane 74. The observation that the MolSA and PSA plots fluctuated less and had marginally different mean values indicating the formation of stable complexes and ability to interact with surrounding molecules and polar environments. The SASA describes the accessibility of a protein surface area to solvent with lower values indicating greater binding stability 74,75. However, the observed fluctuations in SASA plot for all the complexes indicate the differences in accessibility to solvent. The recorded higher mean SASA value of NOx-rutin complex suggests its reduced accessibility to solvent and stability. The observed deviation in the MolSA, PSA, and SASA values for NOx-rutin complex suggests lower binding interaction of rutin with NOx. This implies that rutin possesses reduced potential for modulating NOx relative to other targets in eliciting anti-inflammatory effects.
A notable limitation of this study is the potential variability in the bioactivity of Hura crepitans extracts, which may arise from ecological factors such as geographic origin, soil composition, and climate conditions that influence phytochemical profiles. These variations underscore the importance of standardizing extraction protocols and characterizing the bioactive compounds to ensure reproducibility. Additionally, while the in vitro and in silico methodologies provided valuable insights into the antioxidant and anti-inflammatory properties of the extract and its metabolites, the absence of in vivo validation limits the ability to extrapolate these findings to physiological contexts. Future studies should focus on preclinical in vivo models to confirm the therapeutic potential of these compounds and further elucidate their pharmacodynamics and safety profiles.
Conclusions
This study highlights the antioxidant and anti-inflammatory potential of Hura crepitans through a combination of in vitro and in silico analyses. Key phytochemicals, including epigallocatechin and rutin, were identified as significant contributors to the observed biological activities. The phenolic constituents of CMEHC demonstrated strong binding affinities and stability with key protein targets, suggesting their ability to scavenge free radicals generated during oxidative stress and mitigate inflammatory responses. These findings support the medicinal value of H. crepitans and its potential as a source of therapeutic agents. Further in vivo and clinical studies are needed to validate its efficacy and safety.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Klisic A Ahmad R Haddad D Bonomini F Sindhu S Editorial: The role of oxidative stress in metabolic and inflammatory diseases Front Endocrinol (Lausanne)2024151374584 doi: 10.3389/fendo.2024.13745843839021010.3389/fendo.2024.1374584 PMC 10882066 · doi ↗ · pubmed ↗
- 2Higashi Y Roles of oxidative stress and inflammation in vascular endothelial dysfunction-related disease Antioxidants (Basel)202211101958 doi: 10.3390/antiox 111019583629068110.3390/antiox 11101958 PMC 9598825 · doi ↗ · pubmed ↗
- 3Vassalle C Gaggini M Type 2 Diabetes and Oxidative Stress and Inflammation: Pathophysiological Mechanisms and Possible Therapeutic Options Antioxidants (Basel)202211953 doi: 10.3390/antiox 110509533562481710.3390/antiox 11050953 PMC 9137541 · doi ↗ · pubmed ↗
- 4Rahman MM Rahaman MS Islam MR Role of Phenolic Compounds in Human Disease: Current Knowledge and Future Prospects Molecules (Basel)2021271233 doi: 10.3390/molecules 2701023310.3390/molecules 27010233 PMC 874650135011465 · doi ↗ · pubmed ↗
- 5Spencer JPE Biomarkers of the intake of dietary polyphenols: Strengths, limitations and application in nutrition research Br J Nutr 20089912221766614610.1017/S 0007114507798938 · doi ↗ · pubmed ↗
- 6Pandey KB Rizvi SI Plant Polyphenols as Dietary Antioxidants in Human Health and Disease Oxid Med Cell Longev 200922702782071691410.4161/oxim.2.5.9498 PMC 2835915 · doi ↗ · pubmed ↗
- 7Owojuyigbe OS Firempong SK Larbie C Hepatoprotective Potential of Hura crepitans L.: A Review of Ethnomedical, Phytochemical and Pharmacological J Complementary Altern Med Res 202092110
- 8Adindu EA Elekwa I Ogwo JI Phytochemical comparative screening of aqueous extracts of the leaves, stem barks, and roots of Hura crepitans (L) using GC-FID Nat Sci 20151312112119 doi: 10.7537/marsnsj 131215.15