Post-vaccination evaluation of an erysipelas/parvovirus bivalent vaccine administered to sows during lactation on follicular development and piglet growth
E. Sánchez-Tarifa, FA. García-Vázquez, A. Vela, L. Sanjoaquín, MV. Falceto, A. Fernández-Fontelo, O. Gómez-Duran, C. Alonso, I. Hernández-Caravaca

TL;DR
Administering a bivalent vaccine to sows during lactation negatively affects follicle development and piglet growth.
Contribution
This study is the first to evaluate the effects of a bivalent ER/PPV vaccine on sow follicle development and piglet growth during lactation.
Findings
Non-vaccinated sows had a greater frequency of large follicles compared to vaccinated sows.
Piglets from non-vaccinated sows had higher weight gain and final weight.
Vaccinated sows consumed less feed on the day of injection.
Abstract
Vaccines against Erysipelothrix rhusiopathiae (ER) and porcine parvovirus (PPV) are employed in sow farms worldwide. This study evaluated the impact of administering a bivalent ER/PPV vaccine during lactation. Multiparous sows (n = 83) of two different farms (A (herd size 900 sows) and B (1000 sows)) were allocated in two different groups: 1) sows inoculated with saline solution (non-vaccinated group); 2) sows inoculated with the bivalent vaccine against ER/PPV (vaccinated group). The injections were administered at ∼11 days of lactation. The study was conducted in four different experiments: the first experiment, evaluating the female genital tract (uterus and ovary) and follicle development, was performed in culled sows (n = 19) of Farm A the day after weaning (∼24 days). The remaining experiments, which examined internal temperature, offspring growth, and sow feed intake, were…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
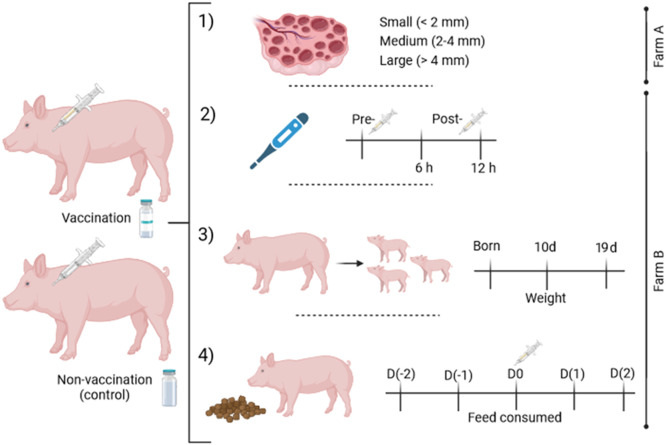 Figure 1
Figure 1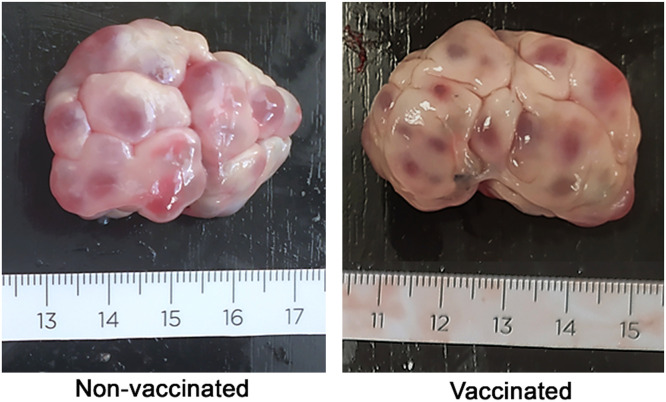 Figure 2
Figure 2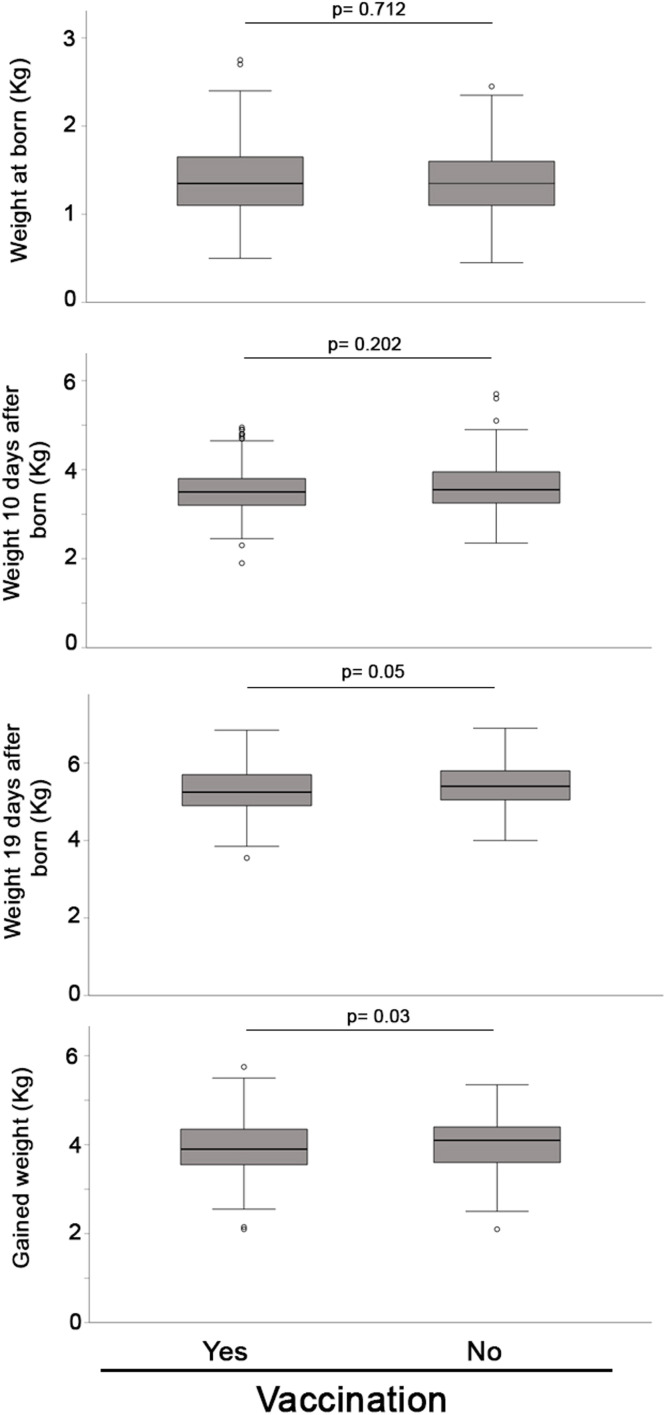 Figure 3
Figure 3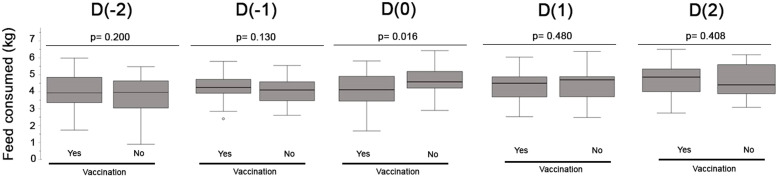 Figure 4
Figure 4Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAnimal Virus Infections Studies · Virus-based gene therapy research · Parvovirus B19 Infection Studies
Introduction
1
Erysipelas in animals is caused by infection with the Gram-positive bacteria Erysipelothrix spp. Erysipelothrix spp. is divided into eight different species which includes Erysipelothrix rhusiopathiae (ER) (Skerman et al., 1980; Buchanan, 1918) belonging to the genus Erysipelothrix in the family Erysipelothrichaceae. Pigs are considered the most important reservoir for ER. It was estimated that 30–50 % of healthy pigs carry the bacteria in tonsils and lymphoid tissues (Steppson & Berman, 1978). Clinically, Erysipelas is characterized by sudden death, pyrexia, red skin patches (red sick-ness), arthritis or cardiac insufficiency (reviewed by Opriessnig et al., 2020). Moreover, reproductive performance is also compromised when Erysipelas is present, being associated with vulval discharges, low fertility, abortions or smaller litter size (Bilkei, 1996; Hoffmann & Bilkei, 2002). So, this bacterium has an important economic influence at different levels of swine industry and can impact all pig production levels such as breeding herd, grow-finish farm or slaughterhouse (reviewed by Opriessnig et al., 2020), reasons why the majority of pig breeding are vaccinated. Essentially, the commercial vaccines available are based in attenuated erysipelas and bacterins. In gilts a common vaccination schedule consists in the i.m. vaccination of the gilts at 22 and 26 weeks of age and ahead of the first insemination. Subsequent revaccinations in multiparous sows are applied in each reproductive cycle 10 days after farrowing because the protective immunity is around 4–6 months (reviewed by Opriessnig et al., 2020).
On the other hand, porcine Parvovirus (PPV) is a virus (family Parvoviridae, Parvovirinae subfamily, and Ungulate protoparvovirus 1 species) present in the majority of pig herds worldwide (van den Born et al., 2020) and considered to be one of the major causes of reproductive failure (Streck & Truyen, 2020). PPV does not cause clinical signs in adults, so this disease can be considered when females return to oestrus with no apparent reasons or delay in parturitions with an increase in the number of mummified fetuses and small litters (Streck & Truyen, 2020), because fetuses infected early in pregnancy usually die, resulting in their mummification or resorption (van den Born et al., 2020). PPV is controlled by a proper management of internal and external biosecurity of the herd and through vaccination. Current commercial PPV vaccines consist of inactivated forms of the original virus (Truyen & Streck, 2012) or recombinant virus (García-Morante et al., 2019). In the case of reproductive females, the vaccination protocols are similar to the ones explained for ER (Truyen & Streck, 2012), because in most of the times the antigens are mixed by the pharmaceutical industry in the same vaccine. As mentioned, vaccines against erysipelas and PPV are routinely used in the pig industry. For user convenience and to reduce the number of injections given to sows in the breeding herd during the lactation period, a typical immunization schedule usually includes the combination of two vaccines as a ready to use product. This approach improves both animal welfare and the labor efficiency of farmers (Sánchez-Matamoros et al., 2019). However, it is also useful for pig health to improve the comfort of the breeding environment because pigs exposed to stress can reduce their immunological responses against pathogens (Morimoto et al., 2022). Certainly, any situation of stress during the lactation period may have an impact on their productivity (feed consumption, milk production, growth of the current litter), risking their maximum productive level or even having a negative impact in the following reproductive cycle. Numerous factors influence the proper development of follicles in sows. These include season, parity, body condition, stress and lactation duration. These factors are considered primary influences on ovarian follicular growth in sows (Quesnel et al., 2005; Peltoniemi et al., 2006; Auvigne et al., 2010; Bertoldo et al., 2011; Knox et al., 2019). In the sows, waves of follicular growth are absent (Lucy et al., 2001). During the luteal and early follicular phases, approximately 50 follicles measuring 1–6 mm are present (Foxcroft et al., 1985). Subsequent to the elimination of inhibitory effects associated with suckling at weaning, follicles that exceed 4 mm (dominant follicles) undergo further growth, while the number of small follicles diminishes and begin atresia (Hafez, 2000). It is important to note, that sows showing smaller follicles at weaning had showed longer intervals for estrus and ovulation and smaller litters at farrowing (Kemp et al., 2005; Lopes et al., 2014; TP Lopes et al., 2020).
Nowadays, vaccines have evolved into safer products noticing their use not only during the lactation period but in a wider range of periods of the reproductive cycle of the sow. Nevertheless, there are no detailed published data about the impact of ER/PPV vaccination on reproductive tract and productive performance with the routinely used schedules. According to the pointed out information, the objective of the present study was to evaluate the effect of the ER/PPV vaccination on sows during lactation, with particular regard to its impact on the reproductive tract, temperature, and feed consumption of the sow, as well as on their offspring (piglets' growth).
Materials and methods
2
Study design
2.1
The design of the study (Fig. 1) was directed to evaluate the impact of the bivalent vaccine (ER/PPV) administered in post-farrowing sows. Concretely, sows were vaccinated (vaccinated group) or injected with sterile physiological saline solution (2 ml) (non-vaccinated) during lactation (∼11 days of lactation in all the experiments) in late summer early autumn. The sows included in the first experiment (experiment 1), were a total of 19 sows (Farm A) of the same batch distributed in non-vaccinated (n = 9, parity= 6.88 ± 0.78) and vaccinated sows (n = 10, parity= 6.60 ± 1.07). They were sacrificed the day after weaning (24.4 ± 4.37 days after farrowing). The follicles of the ovaries were classified according to their size in (i) small (< 2 mm in ø), (ii) medium (2–4 mm in ø), and (iii) large (> 4 mm in ø). Moreover, the weight of the ovaries (g), uterine horns (g), oviducts (g), and diameter of uterine arteries (cm) were recorded.Fig. 1. Scheme of the experimental design. Two groups of sows were allocated in one of the two experimental groups: vaccinated vs. non-vaccinated (control group injected with saline solution). The vaccination was performed ∼11 days of lactation. Experiment 1) Reproductive tracts of weaned sows were recovered from the slaughterhouse and follicles were counted and classified depending on their size (small, medium, or large). Experiment 2) The temperature of the sows was evaluated before vaccination (pre-vaccination) and after vaccination (6 h and 12 h post-vaccination). Experiment 3) The weight of the piglets was analyzed at born, 10 days of age, and before weaning (19 days of age). Moreover, ADWG and total weight gain were calculated. Experiment 4) The feed consumed by the sows of the study was evaluated before (Day(−2) and D(−1)) vaccination, the day of vaccination (D0), and after vaccination (Day(1) and D(2)).Fig 1
In experiment 2, 64 sows (Farm B) of the same batch were distributed in vaccinated (n = 37, parity = 3.85 ± 2.98) and non-vaccinated (n = 27, parity = 3.88 ± 2.80) groups having similar reproductive performance (see Table 3). In both groups, we took the rectal temperature of the sows (pre-vaccination and 6 h and 12 h post-vaccination), and weighted the piglets (at birth, at 10 e and at 19 days of age) and the calculated the (ADWG of piglets from birth to final weight at 19 days of lactation (experiment 3). Finally, the feed consumed by the sows [before (Day(−2) and D(−1)) vaccination, the day of vaccination (D0) and after vaccination (Day(1) and D(2))] were evaluated (experiment 4).
Animals and housing
2.2
A total of 83 crossbred sows [Large-White X Landrace] from two commercial sow farms were used for this study. Farm A consisted of a 900-sow farrow-to-nursery (Genetic Topigs Norsvin, TN70), whereas Farm B comprised 1000-sows in a farrow-to-wean intensive conventional system (Genetic Geneplus Iberia, S.L). Both farms were in Spain's North-eastern area, with an industry-standard biosecurity program in place certified by the herd veterinarian. Sows on the farms were actively mass vaccinated against the porcine respiratory and reproductive syndrome virus (PRRSv) every four months, E. coli and Clostridium perfringens type C (bivalent vaccine) in last trimester of every gestation and against Parvovirus and E. rhusiopathiae (bivalent vaccine) during the lactation period in each cycle. Both farms were stable against PRRSv based on the classification system developed by the American Association of Swine Veterinarians (Holtkamp et al., 2011). It is noteworthy that throughout the course of the study's development, no occurrence of disease outbreaks or alterations in animal diets occurred.
During the trial, animals were allocated in farrowing crates (according to the European Commission Directive for Pig Welfare).
Vaccine
2.3
Sows used in this trial received vaccination against ER and PPV bivalent vaccine following manufacture´s protocol [Eryseng® Parvo, HIPRA, Amer, Spain]. The vaccine's adjuvant is alum, the most widely used vaccine adjuvant due to its safety, cost-effectiveness, and ability to enhance the body's immune response to various antigens. (Di Benedetto et al., 2024). The treatment was administered intramuscular (i.m.) into the neck musculature caudal of the ear base and at a volume of administration of 2 ml per animal. All injections were performed by a dedicated worker not involved with data collection.
Uterine tract postmortem examination
2.4
The experiment was conducted using only those sows that had been selected for culling as part of the farm's standard culling procedure following weaning. None of the selected sows exhibited any reproductive problems in their reports prior to culling. The genital tract of the sows (ovarian structures, oviduct, uterine horns, uterine arteries) was evaluated the day after weaning. Measurements of ovarian follicle size (right and left) and uterine artery size were performed with a digital caliber (RS PRO) by an expert under single-blind study conditions. The weight (scale in grams-g) of the ovaries, oviducts, and uterine horns was recorded with a precision digital scale (Nahita 5041).
Insemination procedure, pregnancy diagnosis, farrowing and litter performance
2.5
The insemination was carried on after estrus detection, that was performed twice daily by experienced workers by allowing sows nose-to-nose contact with mature boars and applying back pressure. The occurrence of estrus was defined by the standing reflex in front of a teaser boar and reddening and swelling of the vulva. Only sows with clear signs of estrus were used for the experiment. The post-cervical artificial insemination procedure was performed as described by Hernández-Caravaca et al. (2012). Pregnancy was confirmed by ultrasound 23–28 days after insemination by transabdominal ultrasonography (Echoscan T-300 S, Barcelona, Spain). At 110 days of gestation, pregnant sows were moved from the gestation facilities to the farrowing pens. Farrowing data were recorded for each sow. At the end of farrowing, farrowing rate (%) and the number of piglets born were recorded. Each piglet was classified as piglet born alive, dead, or mummified. The weaning age of the piglets in both farms was ∼24 days. In experiment 2, alive piglets were identified with eartags and were assigned to their mother sow at birth for precise recording. During the lactation period, crossfostering was allowed within each group. Individual body weights (kg) of the piglets were collected on the day of birth and a few days before weaning using a digital handing scale. Moreover, piglet average daily weight gain (ADWG) was obtained for the surviving piglets (calculated from the day of birth to the day of the last weighing at 19 days after birth). The piglets were provided with an ad libitum supply of commercial diets commencing on the sixth day of lactation.
Rectal temperature
2.6
Temperatures were measured rectally in the sows using self-calibrated digital thermometers at approximately the same time each day and recorded in degrees Celsius (°C) units. Pyrexia was defined as an increase in temperature of >1.5 °C respective to the baseline temperature of the individual animal (pre-vaccination temperature) (Noguera et al., 2021).
Feed intake
2.7
Daily feed intake data were individually obtained by sow using control of feeding consumption devices (GESTAL SOLO, JYGA Technologies, Quebec, Canada). Feed was available ad libitum with commercial diets for lactating sows.
Statistical analysis
2.8
Statistical analyses were performed with the SPSS 24.0 software package (IBM SPSS Inc., Chicago, IL, USA). Sow characteristics parameters were analyzed for normality by a Shapiro–Wilk test, in which showed that all parameters had a normal distribution. The Chi-squared test was used to evaluate the distribution of follicles within different follicle size classes between both experimental group (vaccinated vs non-vaccinated). The proportion of each follicle category (small, medium, large) between non-vaccinated and vaccinated animals was compared by t-test (data with normal distribution; small and large categories) or Wilcoxon test (data with non-normal distribution; medium category). The results are represented as the mean ± SD (standard deviation) or mean ± SEM (standard error of the mean) accordingly. Values were considered significantly different when p ≤ 0.05. A statistical trend was considered when p > 0.05 and p < 0.07. The correlation between the number of follicles (small, medium, and large) and ovary weight and uterine artery size was assessed by Pearson´s correlation.
The reproductive performance of the sows used in the study was analyzed for normality by the Kolmogorov-Smirnov test, which showed that all parameters had non-normal distribution, so, the non-parametric U-Mann Whitney test was used. The results are represented as the mean ± SD (standard deviation). Values were considered significantly different when p ≤ 0.05.
Rectal temperature data were analyzed using the Statistic Analysis Software (SAS, University Edition 2016). Sphericity of the data was checked for the temperature of the sows (pre-vaccination, 6 h, and 12 h post-vaccination). All the variables confirmed the sphericity of the data, and they were analyzed using Proc Mixed procedures. The model included the treatments (vaccinated vs. non-vaccinated sows), the three points of evaluation (pre-vaccination, 6 h post-vaccination, and 12 h post-vaccination), and the interaction between these as the main effect, with different samples and sows as the random effect. Data temperatures are represented as the mean ± SD (standard deviation).
The weights of piglets were analyzed for normality by a Kolmogorov-Smirnov test, which showed that all parameters had a non-normal distribution except weight gain from birth to the final weight, piglet weight at the end of the study and ADWG from birth to the final weight. Then, the U-Mann-Whitney test was used for non-parametric parameters and the T-test for parametric parameters. Values were considered significantly different when p ≤ 0.05. A statistical trend was considered when p > 0.05 and p < 0.07. Weight data in the text are represented as the mean ± SD (standard deviation). The feed consumed per sow was analyzed for normality by a Shapiro–Wilk test, which showed that the data had a normal distribution, so a t-student test was applied. Values were considered significantly different when p ≤ 0.05. Feed consumed data in the text are represented as the mean ± SD (standard deviation).
Results
3
Sows used in experiment 1 (Table 1; Farm A) showed similar characteristics between both experimental groups. Gross examinations did not reveal significant differences between vaccinated and non-vaccinated females based on the weights of the ovaries, uterus, and oviduct.Table 1. Characteristics (number of parities and reproductive tract) of the sows from the two experimental groups (non-vaccinated vs. vaccinated sows). Data are expressed as mean ± SD. The values are the mean of right and left structures (ovary, oviduct, uterine horn, and uterine arteries) and the mean total number of follicles per sow. No significant differences were observed (p > 0.05).Table 1NParityOvaries weight (g)Total number of follicles per sowOviduct weight (g)Uterine horns weight (g)Uterine arteries (cm)Non-vaccinated96.88 ± 0.787.83 ± 1.6067.22 ± 16.023.31 ± 0.44304.20 ± 47.970.57 ± 0.07Vaccinated106.60 ± 1.077.01 ± 1.6374.60 ± 18.233.26 ± 1.28309.00 ± 51.900.53 ± 0.06p-value0.2350.8330.3640.1160.9370.685
The frequencies of each follicle size were significantly different between vaccinated vs non-vaccinated sows (p < 0.001), with vaccinated sows showing a lesser frequency of large follicles and a higher frequency of small follicles than non-vaccinated sows. When the percentage of follicles in each category (small, medium or large) was compared, a significant higher rate of large follicles was observed in non-vaccinated compared to vaccinated animals (Table 2), (Fig. 2).Table 2. Percentage of small, medium and large follicles per ovary from the two experimental groups (non-vaccinated vs. vaccinated sows). Data are expressed as mean ± SEM. No significant differences were observed (p > 0.05). Dots-graph showing the percentage of follicular size [small (represented in blue), medium (represented in grey) and large (represented in green)].Table 2. OvariesSmallfollicles (%,Image, table 2)Mediumfollicles (%,Image, table 2)Largefollicles (%,Image, table 2)Dots-graphNon-vaccinated1824.95 ± 4.0439.36 ± 6.3535.65 ± 4.97Image, table 2Vaccinated2035.41 ± 5.6144.71 ± 5.7819.89 ± 3.39Image, table 2p-value0.120.330.035Fig. 2Ovaries of non-vaccinated and vaccinated sows the day after weaning at the beginning of the follicular phase, after lactation anestrus. Different follicle sizes are observed in both groups. Scale in cm.Fig 2
No correlation was detected between the size of the follicles and ovary weight (p > 0.05), but a positive significant correlation was established between the number of large follicles and the size of the uterine artery (r = 0.678; p = 0.022). In addition, when this last correlation was performed separately in the two experimental groups, a statistical trend was found for non-vaccinated sows (r = 0.872; p = 0.054) but correlation was not observed for the vaccinated sows (r = 0.323; p = 0.533).
At farm B, all animals showed normal reproductive performance prior to the experiments without differences between the experimental groups (Table 3). The vaccination (or placebo in the case of the control group) of the sows was performed after ∼11 days of lactation (11.27 ± 1.52 and 11.22 ± 1.39 days for vaccinated and non-vaccinated sows, respectively; p = 0.939).Table 3. Reproductive performance in sows used for both experimental groups (non-vaccinated vs. vaccinated sows). Data are shown as mean ± SD. No significant differences were observed (p > 0.05).Table 3NParityTotal piglets bornPiglets born alivePiglets born deadMummified pigletsVaccinated333.85 ± 2.9814.15 ± 3.4013.01 ± 3.340.97 ± 1.520.06 ± 0.37Non-vaccinated313.88 ± 2.8013.98 ± 3.6812.63 ± 2.811.30 ± 1.900.14 ± 0.60p-value0.9270.8770.4810.2650.345
In experiment 2, the sows’ rectal temperature during the trial (pre- and post-vaccination) did not display any differences between vaccinated and non-vaccinated sows (p > 0.05). Moreover, no adverse local reactions were observed in any of the investigated groups.
In experiment 3, the number of lactation days until the final weight studied were 19.18 ± 1.34 and 19.25 ± 1.43 days for vaccinated and non-vaccinated sows respectively (p = 0.617). The weights of piglets between groups at birth and at 10 days of lactation were similar (1.39 ± 0.39 and 1.37 ± 0.37 kg for piglets from vaccinated and non-vaccinated sows at birth; p = 0.712 and 3.54 ± 0.54 and 3.64 ± 0.59 kg for piglets from vaccinated and non-vaccinated sows at 10 days; p = 0,202). Statistical differences were observed at final weight near weaning (5.28 ± 0.57 and 5.40 ± 0.56 kg for piglets from vaccinated and non-vaccinated sows, respectively; p = 0.05). When the total weight gained per piglet between birth and the final weigh was analyzed, the results indicated a significant improvement for the piglets from non-vaccinated sows (3.89 ± 0.62 and 4.02 ± 0.60; for vaccinated and non-vaccinated sows respectively, p = 0.03) (Fig. 3).Fig. 3. Box-plot comparing the weight (kg) (at born, after 10 days n= and around 19 days after born, and total gained weight (n = 221 and n = 158 in vaccinated and non-vaccinated group respectively) of the piglets born in the two groups of sows (non-vaccinated vs. vaccinated). Significant differences were observed when p < 0.05. Small circles (°) and asterisks (*) in the box-plot represents outliers and extreme cases, respectively.Fig 3
ADWG during the studied period showed a statistical trend in favor the non-vaccinated group (0.204 ± 0.032 and 0.210 ± 0.034; for vaccinated and non-vaccinated sows respectively, p = 0.06).
When the feed consumed per sow (experiment 4) was analyzed the 2 days previous to vaccination (D(−2) and D(−1)), the day of vaccination (D(0)), and 2 days after vaccination (D(1) and D(2)), a significant difference was observed the D0 (p = 0.016) showing a lower feed consumption per sow in the vaccinated group compared to the non-vaccinated group (4.09 ± 1.03 and 4.62 ± 0.82 kg, Fig. 4).Fig. 4. Box-plot comparing the feed consumed per sow (kg) the 2 days previous to vaccination (D(−2) and D(−1)), the day of vaccination (D(0)) and 2 days after vaccination (D(1) and D(2 in the two groups of sows (vaccinated vs. non-vaccinated). Significant differences were observed when p < 0.05. Small circles (°) in the box-plot represents outliers and extreme cases, respectively.Fig 4
Discussion
4
Successful swine production aims to deliver as many live-born and healthy piglets as possible by, among other factors, preventing infectious diseases affecting reproductive output. Vaccination is one of the most successful and cost-effective interventions to prevent swine diseases such as erysipelas and parvovirus. However, vaccination strategies on farms should be established to achieve not only an adequate level of immunity against pathogens but also to decrease the stress of the animals in critical periods of swine production to maximize its production and welfare. Our study has shown the effect of the administration of a combined vaccine of ER and PPV during the lactation period on reproductive and productive efficiency.
Few studies have analyzed the putative impact of vaccination on the reproductive tract and ovary function, except those related to ovarian regulation (Jinshu et al., 2005; Wang et al., 2012) or contraception (Ma et al., 2012; Dalmau et al., 2015) through vaccines. First, we compared the effect of vaccination during day 11 of lactation on the ovary and reproductive tract. The study demonstrates that this ER/PPV vaccine has no impact on the female reproductive tract and the total number of follicles in the ovaries did not vary between non-vaccinated and vaccinated sows. This fact agrees with a previous report comparing PRRSV -vaccinated and non-vaccinated sows concerning gross measurements of ovaries and uterus (Papatsiros et al., 2015). However, our results indicated that the non-vaccinated sows showed a higher frequency of larger follicles than vaccinated sows. In pigs, some studies have shown, that developmental competence of pig oocytes improves as follicular size increases (Marchal et al., 2002). To the best of our knowledge, this is the first study analyzing the impact of vaccination on porcine follicular size. This is of paramount importance because sows showing smaller follicles at weaning had showed longer intervals for estrus and ovulation and smaller litters at farrowing (TP Lopes et al., 2020). Lopes et al. also stated that differences in follicles size in the early stage of lactation persisted throughout the entire period and weaning, reinforcing the notion that vaccinations and stress should be minimized during this stage (TP Lopes et al., 2020).
In our study, a positive correlation was also found between large follicle proportion and uterine artery size in the non-vaccinated group. Although in another species the positive correlation observed between ovarian follicular size and uterine artery vascular indices was demonstrated in cows (Yama et al., 2022).
The populations of follicles observed in the present study agree with the physiology of the ovary during lactation. Small and medium-sized follicles (no larger than 3 mm) are present during the second week of lactation for a later increase in their size because of a higher frequency of luteinizing hormone (LH) pulses but not developed beyond 5 mm in diameter during lactation (Quesnel, 2009). One aspect to consider is the impact of feed reduction intake on the day of vaccination on follicular development. Our results indicated a reduction of follicle size population in the vaccinated sows and a significant decrease in feed intake on the day of vaccination in this group of animals. It has been demonstrated that gilts deprived of glucose reduced LH pulse frequency (Barb et al. 2001) which may be related to the reduction in the follicular growth observed in our study. Restriction feed to sows were reported to inhibit secretion of LH and result in fewer follicles larger than 4 mm at weaning and two days later (Quesnel et al., 1998). So, nutritional deficiency during lactation can alter follicular growth.
Overall, the administered vaccine did not induce a significant increase in the mean rectal temperature compared to non-vaccinated animals. Furthermore, none of the animals of the study had a rectal temperature increase above 1.3 °C (including vaccinated and non-vaccinated animals) after injection except two animals injected with the vaccine which had an increase of 1.9 °C. These findings agree with previous reports using the same bivalent vaccine in pregnant gilts (Noguera et al., 2021) or different PPV vaccine in lactating sows (García-Morante et al., 2019).
The number of pigs weaned per sow is commonly used as a measurement to benchmark the productivity of breeding herds (Koketsu et al., 2017). However, other factors such as piglet quality should be considered. A previous study using a different vaccine showed no differences between the vaccinated group and non-vaccinated regarding ADWG in piglets (García-Morante et al., 2019), and vaccine applied was only against PPV but not ER, and the study was longer than our study (19 vs. 28 days). Although in this study similar to our results a significant difference in piglet weight gain from birth to the final weight at weaning in favor of non-vaccinated group was found. Moreover, the AWDG trend to be higher in the piglets from the non-vaccinated group. This aspect may be associated with the reduced sow feed intake in the period around vaccination, where the feed consumed significantly decreased the day of inoculation in the vaccinated group. Feed intake is a critical point in lactating sows and lower lactation feed intake has been associated with a lower average weaning weight of piglets and reproductive failures (Koketsu et al., 1996) which agrees with our results.
Despite the mild nature vaccine's effects, our study indicates that its administration during lactation can have several adverse consequences on sow physiology and offspring performance, as previously discussed. In this regard, adverse reactions to vaccination may result to the inflammatory response to the antigen or the presence of an adjuvant in the vaccine (Spickler et al., 2003). The alum used as an adjuvant in the vaccine administered in our study can activate cellular transcription factors and stimulate the production of various signaling molecules. For example, it can trigger the Nalp3 inflammasome, leading to a humoral adaptive immune response (IgE production) (Eisenbarth et al., 2008; Kool et al., 2008; Kuroda et al., 2011). Additionally, alum can activate dendritic cells through receptor-independent uptake, induce lipid mediators such as prostaglandins, and stimulate the innate immune system by host DNA released from alum-injured cells (Flach et al., 2011; Kuroda et al., 2011; Marichal et al., 2011). Moreover, high doses of aluminum hydroxide injected without antigens has demonstrated to cause granulomas in pigs (Valtulini et al., 2005). However, alum remains the most widely used vaccine adjuvant due to its safety, cost-effectiveness, and ability to enhance the immune response to various antigens (Di Benedetto et al., 2024). Including an experimental group receiving only the adjuvant could have provided a more comprehensive interpretation of the results. We acknowledge this is a limitation of the present study; however, it is important to note that all commercially available bivalent ER/PPV vaccines come as ready-to-use suspension for injection, and the focus is on evaluating whether the complete product available to veterinarians (adjuvant + antigen) influences reproductive performance.
Conclusions
5
This study determined that administering the bivalent ER/PPV vaccine to sows during lactation can have a detrimental effect on follicle development, leading to a reduced frequency of large follicles. Additionally, piglets from vaccinated sows exhibited lower final weight, reduced weight gain, and a tendency toward lower average daily weight gain during lactation. Furthermore, vaccinated sows had lower feed consumption on the day of vaccination compared to the non-vaccinated group.
It is well established that ER and PPV are major contributors to reproductive failure in the swine industry, and vaccination remains the most cost-effective strategy to prevent these diseases worldwide.
The findings of this study highlight the need for further research to evaluate alternative vaccination strategies, such as administration during gestation or as part of a mass vaccination program that reduces vaccine administration during lactation, to mitigate potential negative effects on sow reproductive performance and piglet growth.
Institutional review board statement
Through the experiments, animals were handled carefully avoiding any unnecessary stress. All experiments were performed following relevant guidelines and regulations. The study was carried out in compliance with the ARRIVE guidelines (https://arriveguidelines.org/). An Internal consultation regarding the need of a full review from an Ethics committee was carried out with two experts in Boehringer Ingelheim: Spanish Regulatory and Pharmacovigilance Head and European Head of Clinical Studies. The ethics approval was deemed unnecessary according to European and national legislation (Directive 2010/63/EU of the European parliament and RD53/2013). In both regulation state that the ethics approval is not necessary in case of: a) non-experimental agriculture practices, b) non-experimental clinical veterinary practices and d) practices undertaken for the purposes of recognized animal husbandry. This study is framed in these assumptions because the procedures carried out are commonly used in the farms.
Informed consent statement
Ganados Gili, S.A. (Owner of the animals) consented to participate and stated that the study is about the performance impact of current vaccine protocols in their farms. Likewise, the study utilized reproductive tracts obtained from sows that were culled as part of the farm's routine culling process. All their staff involved are trained to use of the best welfare practices in swine production and part of their veterinarians actively monitored and performed the research.
CRediT authorship contribution statement
E. Sánchez-Tarifa: Writing – review & editing, Investigation, Data curation, Conceptualization. FA. García-Vázquez: Writing – review & editing, Writing – original draft, Conceptualization. A. Vela: Investigation. L. Sanjoaquín: Investigation. MV. Falceto: Investigation. A. Fernández-Fontelo: Formal analysis, Data curation. O. Gómez-Duran: Writing – review & editing. C. Alonso: Writing – review & editing. I. Hernández-Caravaca: Writing – review & editing, Writing – original draft, Conceptualization.
Declaration of competing interest
The authors declare the following financial interests/personal relationships which may be considered as potential competing interests:
E. Sanchez-Tarifa reports financial support, article publishing charges, and equipment, drugs, or supplies were provided by Boehringer Ingelheim Vetmedica GmbH. E. Sanchez-Tarifa reports a relationship with Boehringer Ingelheim GmbH that includes: employment. O.G. Duran reports a relationship with Boehringer Ingelheim Vetmedica GmbH that includes: employment. C. Alonso reports a relationship with Boehringer Ingelheim Vetmedica GmbH that includes: employment. If there are other authors, they declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Auvigne V.Leneveu P.Jehannin C.Peltoniemi O.SalléE.Seasonal infertility in sows: A five year field study to analyze the relative roles of heat stress and photoperiod Theriogenology 7412010606610.1016/j.theriogenology.2009.12.019Jul 1Epub 2010 Mar 1. PMID: 2018963620189636 · doi ↗ · pubmed ↗
- 2Barb C.R., Barrett J., Kraeling R.R., Rampacek GB 2001 b. Serum leptin concentrations, luteinizing hormone and growth hormone secretion during feed and metabolic fuel restriction in the prepuberal gilt. Domestic Animal Endocrinology 20 47–63.10.1016/s 0739-7240(00)00088-611164333 · doi ↗ · pubmed ↗
- 3Bertoldo M.J.Holyoake P.K.Evans G.Grupen C.G.Seasonal effects on oocyte developmental competence in sows experiencing pregnancy loss Animal Reproduction Science 1241–2201110411110.1016/j.anireprosci.2011.02.012Mar Epub 2011 Feb 17. PMID: 2138267721382677 · doi ↗ · pubmed ↗
- 4Bilkei G.Sauenmanagement. Vet spezial. Gustav Fischer Verlag Jena and Stuttgart 1996357
- 5Buchanan R.E.Studies in the nomenclature and classification of the bacteria Journal of Bacteriology 31918276110.1128/JB.3.1.27-61.191816558776 PMC 378743 · doi ↗ · pubmed ↗
- 6Dalmau A.Velarde A.Rodríguez P.Pedernera C.Llonch P.Fàbrega E.Use of an anti-Gn RF vaccine to suppress estrus in crossbred Iberian female pigs Theriogenology 843201534234710.1016/j.theriogenology.2015.03.025Aug 25908572 · doi ↗ · pubmed ↗
- 7Di Benedetto R.Massai L.Wright M.Adjuvanted modified bacterial antigens for single-dose vaccines International Journal of Molecular Sciences 252120241146110.3390/ijms 252111461 Published 2024 Oct 2539519015 PMC 11546299 · doi ↗ · pubmed ↗
- 8Eisenbarth S.C.Colegio O.R.O'Connor W.Sutterwala F.S.Flavell R.A Crucial role for the Nalp 3 inflammasome in the immunostimulatory properties of aluminium adjuvants Nature 453719820081122112610.1038/nature 0693918496530 PMC 4804622 · doi ↗ · pubmed ↗