IL-17 signaling pathway in SPP1+ macrophages drives digestive tract cancer progression
Yihong Wei, Ying Xu, Yanqiong Zeng, Amin Zhang, Xiangling Xing, Wancheng Liu

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
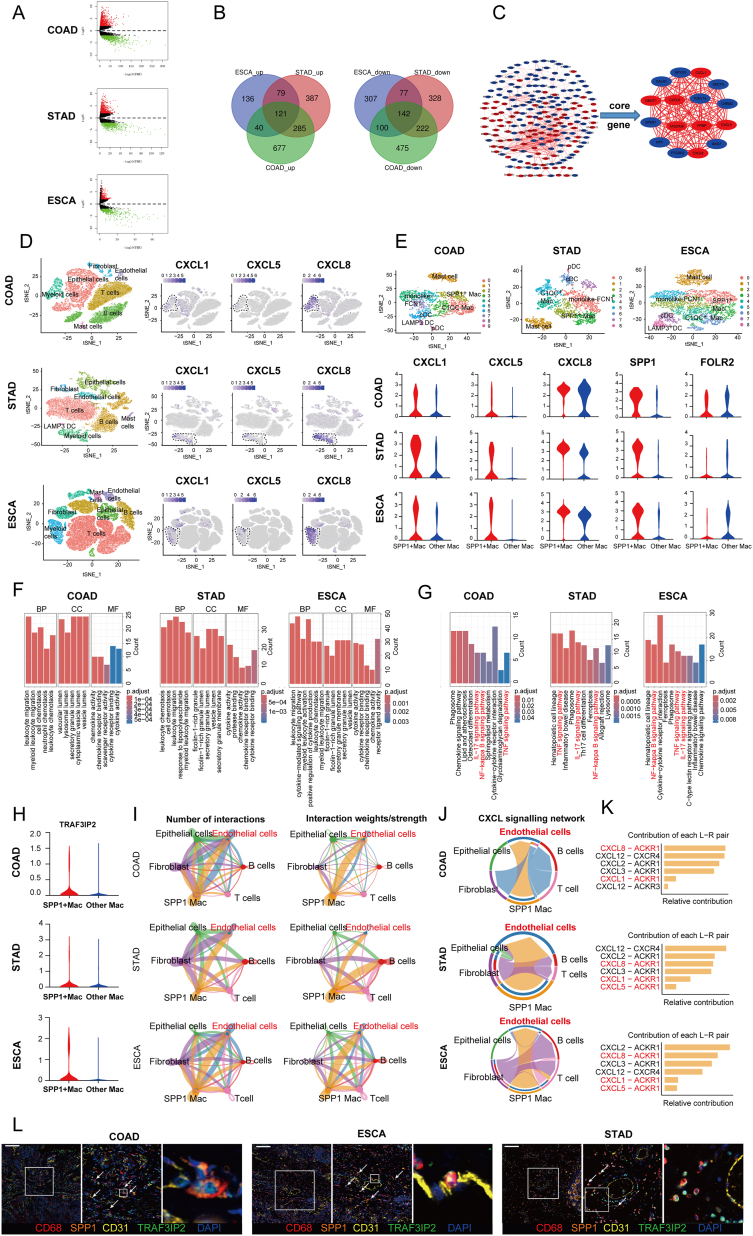 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCancer Immunotherapy and Biomarkers · Phagocytosis and Immune Regulation · IL-33, ST2, and ILC Pathways
Digestive tract cancers (DTCs) are cancers that occur in the gastrointestinal tract and related organs, including esophageal, gastric, and colorectal cancer. Since these types of cancer share similar endoderm developmental origins, the genomic and other molecular features can possess many similarities. Therefore, it is urgent to explore the commonalities of molecular characteristics and signaling pathways in the tumor microenvironment among these diseases.
To uncover the shared molecular characteristics in DTCs, we obtained the recent RNA sequence profiles from The Cancer Genome Atlas (TCGA) database, encompassing esophageal carcinoma (ESCA), stomach adenocarcinoma (STAD), and colon adenocarcinoma (COAD). Differentially expressed gene (DEG) analysis revealed that ESCA exhibited 376 up-regulated and 626 down-regulated mRNAs, STAD showed 872 up-regulated and 769 down-regulated mRNAs, and COAD had 1123 up-regulated and 939 down-regulated mRNAs (Fig. 1A; Fig. S1A and Table 1). To investigate the existence of co-expression of DEGs in DTCs, we employed Venn diagrams to analyze the overlapping and distinct DEGs in DTCs (Fig. 1B). Subsequently, we explored the relevant signaling pathways associated with these DEGs in the mentioned cancer types using the DAVID tool. Based on the number of genes involved, we prioritized the first eight signal pathways with the highest gene enrichment (Fig. S1B). Additionally, we constructed a protein–protein interaction network complex involving common DEGs to identify key differentially expressed core genes within these cancer types. As shown in Figure 1C, sixteen core genes were identified, of which seven were up-regulated (including CXCL1/5/6/8), indicating that the CXCL family might play a critical role in DTCs.Figure 1IL-17 signaling pathway in SPP1^+^ macrophages drives digestive tract cancer (DTC) progression. (A) The volcano plot of differentially expressed genes (DEGs) in DTCs. (B) Co-expression of DEGs in DTCs was detected by Venn diagram. (C) Protein–protein interaction analysis of common DEGs by STRING online database and Cytoscape. (D) T-distributed stochastic neighbor embedding (t-SNE) plots of DTCs, which are grouped into seven main parts including endothelial cells, fibroblasts, endothelial cells, T/natural killer cells, B cells, mast cells, and myeloid cells. The feature plot of CXCL1, CXCL5, and CXCL8 expression in the seven main groups was shown on the right. (E) t-SNE plots of the myeloid cells in DTCs. The violin plot of CXCL1, CXCL5, CXCL8, SPP1, and FOLR2 expression in myeloid cells of DTCs was shown. (F) The bar plots showing differences in GO function pathways based on the DEGs of SPP1^+^ macrophages among colon adenocarcinoma (COAD), stomach adenocarcinoma (STAD), and esophageal carcinoma (ESCA). (G) The bar plots showing differences in KEGG pathways based on the DEGs of SPP1^+^ macrophages among COAD, STAD, and ESCA. (H) Violin plot of TRAF3IP2 expression in myeloid cells of DTCs. (I) Interaction net count plot between SPP1^+^ macrophages and other cells. (J) The chord plot showing the inferred intercellular communication network of CXCL in COAD, STAD, and ESCA. (K) The relative contribution of CXCL receptor ligands in COAD, STAD, and ESCA. (L) Representative images of multiplex-immunostaining showed the relationship between SPP1^+^/TRAF3IP2^+^ macrophages and vascular endothelium cells. Scale bar = 100 μm.Figure 1
The CXCL family is a key group of chemokines capable of inducing leukocyte migration and infiltration. Furthermore, CXCL1/5/8 have the capacity to stimulate angiogenesis and promote the proliferation of endothelial cells, processes that are intimately linked to tumor initiation and development. To verify the reliability of our bioinformatics analysis results, we collected specimens from cancer patients for verification. The PCR results showed that compared with the paracancerous tissue, the expression of CXCL1/5/8 was up-regulated in DCT cancer tissue compared with adjacent tissues (Fig. S2). To gain deeper insights into the common features of CXCL1/5/8 in these three types of cancer, we downloaded scRNA sequencing profiles from the GEO database and conducted an extensive analysis.1, 2, 3 After strict quality control and filtration, we collected 37,768 (COAD), 21,983 (STAD), and 79,261 (ESCA) single cells originating from DTC patients. We classified all cells into 6–10 major clusters representing major cell types, including CD45-negative cells (comprising epithelial cells, fibroblasts, and endothelial cells) and CD45-positive cells (involving T/natural killer cells, B/plasma cells, myeloid cells, and mast cells) (Fig. 1D; Fig. S1C). To gain a deeper understanding of the role of *CXCL1/5/*8 in DTCs, we analyzed the origin of these core genes. Our observation revealed that these genes in DTCs were predominantly found in myeloid cells (Fig. 1D). Consequently, we conducted further sub-clustering of the myeloid cell subtype, employing conventional marker genes to identify dendritic cells, monocytes, macrophages, and mast cells (Fig. S1D). Interestingly, we discovered a significant overlap in the expression of CXCL1/5/8 with secreted phosphoprotein 1-positive (SPP1^+^) macrophages. Additionally, the expression of CXCL molecules was mutually exclusive with complement C1q C chain-positive (C1QC^+^)/folate receptor beta-positive (FOLR2^+^) macrophages (Fig. 1E; Fig. S1E). Although CXCL1 and CXCL8 are also expressed in the mono-like FCN1 cell subset, our study was aimed at macrophages. Considering that both C1QC^+^/FOLR2^+^ and SPP1^+^ macrophages may arise from the state of monocyte-like FCN1 cells in the tumor, we believe that this does not affect the fact that SPP1^+^ macrophages are the main source of CXCL molecules. Collectively, these findings suggest that SPP1^+^ macrophages may serve as the primary source of chemokines CXCL1/5/8 within the tumor microenvironment of DTCs. This role is distinct from other macrophages, including C1QC^+^/FOLR2^+^ macrophages.
With the advancement of scRNA sequencing technology, there is a preference for defining macrophages using SPP1, FOLR2, and TREM2, rather than the traditional M1 and M2.4 Considering the critical role of SPP1^+^ macrophages in the DTC tumor microenvironment, we employed the GEPIA2 website to analyze the correlation between the presence of SPP1^+^ macrophages and the survival of DTC patients. We selected the top 10 most significant genes to serve as representatives of SPP1^+^ macrophages and assessed their impact on patient survival. The results indicated that higher infiltration of SPP1^+^ macrophages led to shorter overall survival in DTC patients (Fig. S1F). Additionally, we conducted Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analyses on these three kinds of cancer. The KEGG enrichment analysis revealed the shared pathways in SPP1^+^ macrophages among COAD, STAD, and ESCA, including interleukin (IL)-17, nuclear factor-kappa B (NF-κB), and tumor necrosis factor-alpha (TNF-α) signaling pathways. Furthermore, it was observed that the C1QC^+^/FOLR2^+^ macrophages in DTCs did not exhibit significant enrichment in these pathways, particularly the IL-17 signaling pathway (Fig. 1F, G; Fig. S1G), which indicated that the IL-17 signaling pathway might hold a distinctive role in SPP1^+^ macrophages.
It is widely accepted that the IL-17 activity leads to the creation of a pro-tumor microenvironment, relying on the production of inflammatory mediators that reshape the tumor microenvironment by CXCL1/5/8. To investigate this further, we evaluated the primary IL-17 receptor (IL17RA) (Fig. S1H). However, it was disappointing to find that there was no significant difference in IL17RA expression between SPP1^+^ and C1QC^+^/FOLR2^+^ macrophages, suggesting that there might be other mechanisms contributing to the differential secretion of CXCL1/5/8 between these two types of macrophages in the IL-17 signaling pathway. We further estimated other molecules related to IL-17 signaling pathways. TRAF3 interacting protein 2 (TRAF3IP2) is the key adaptor molecule directly recruited to IL-17R and is required for both the transcriptional and post-transcriptional changes induced by IL-17.5 We found that TRAF3IP2 was highly expressed in the SPP1^+^ macrophages compared with other macrophages (Fig. 1H), suggesting that TRAF3IP2 deficiency might be the reason for the low expression of CXCL1/5/8 in C1QC^+^/FOLR2^+^ macrophages. It is the high expression of TRAF3IP2 that enables SPP1^+^ macrophages to play a role as a source of CXCL1/5/8 in the tumor microenvironment.
Within the tumor microenvironment, the intercellular signaling network plays a vital role in regulating tumor initiation and development. Recognizing the importance of CXCL1/5/8 in reshaping the vascular network and extracellular matrix to create a conducive environment for tumor cell growth, we employed Cellchat to assess how SPP1^+^ macrophages impact other cells within the tumor microenvironment of DTCs. Cellchat allowed us to understand intracellular interactions by predicting protein–protein interactions. The interaction net count plot demonstrated that SPP1^+^ macrophages exhibited the highest level of interactions with endothelial cells (Fig. 1I; Fig. S1I). Furthermore, the CXCL signaling pathways between SPP1^+^ macrophages and endothelial cells were further analyzed and confirmed (Fig. 1J, K; Fig. S3A). These results suggested that CXCL1/5/8 secreted by SPP1^+^ macrophages might play a significant role in communicating with vascular endothelial cells, enhancing angiogenesis, and exerting their tumor-promoting effects in DTCs. Additionally, we utilized multiple fluorescence immunohistochemistry to detect the relationship between SPP1^+^ macrophages and endothelial cells. Our results indicated that there were many SPP1^+^ macrophages near the vascular endothelium, which also highly expressed TRAF3IP2 (Fig. 1L). These results also suggested that SPP1^+^ macrophages might play a role in promoting the progression of cancer through communication with vascular endothelial cells.
In summary, our study first establishes the connection between SPP1^+^ macrophages and CXCL1/5/8 in DTCs (Fig. S4). SPP1^+^ macrophages constitute the primary source of CXCL1/5/8 in DTCs. Furthermore, we speculated that TRAF3IP2 might serve as the crucial adaptor protein for the production of CXCL1/5/8 and exhibits high expression exclusively in SPP1^+^ macrophages, which have distinctive functions in the IL-17 signaling pathway. Consequently, SPP1^+^ macrophages can recruit myeloid cells and augment angiogenesis through CXCL1/5/8, thereby fostering a cancer-promoting microenvironment. Targeting TRAF3IP2 presents a novel therapeutic strategy for DCTs.
Ethics declaration
Our study was approved by the Medical Ethical Committee of Qilu Hospital of Shandong University (KYLL-202311-043) and consent was gained from all participants.
Funding
This work was supported by the National Science Funds of China (No. 82300219), and the 10.13039/100017445Natural Science Foundation of Shandong Province, China (Youth Program; No. ZR2023QH249).
CRediT authorship contribution statement
Yihong Wei: Formal analysis. Ying Xu: Methodology. Yanqiong Zeng: Writing – original draft. Amin Zhang: Data curation. Xiangling Xing: Writing – original draft. Wancheng Liu: Conceptualization, Funding acquisition.
Data availability
The single-cell RNA sequencing datasets analyzed during the current study are available in the GEO repository (GSE166555 for COAD, GSE167297 for STAD, and GSE160269 for ESCA). For other original data, please contact the corresponding author.
Conflict of interests
The authors declared no competing interests.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Uhlitz F.Bischoff P.Peidli S.Mitogen-activated protein kinase activity drives cell trajectories in colorectal cancer EMBO Mol Med 13102021 e 1412310.15252/emmm.202114123 PMC 849545134409732 · doi ↗ · pubmed ↗
- 2Jeong H.Y.Ham I.H.Lee S.H.Spatially distinct reprogramming of the tumor microenvironment based on tumor invasion in diffuse-type gastric cancers Clin Cancer Res 27232021652965423438529610.1158/1078-0432.CCR-21-0792 · doi ↗ · pubmed ↗
- 3Zhang X.Peng L.Luo Y.Dissecting esophageal squamous-cell carcinoma ecosystem by single-cell transcriptomic analysis Nat Commun 121202152913448943310.1038/s 41467-021-25539-x PMC 8421382 · doi ↗ · pubmed ↗
- 4Bill R.Wirapati P.Messemaker M.CXCL 9:SPP 1 macrophage polarity identifies a network of cellular programs that control human cancers Science 381665720235155243753572910.1126/science.ade 2292 PMC 10755760 · doi ↗ · pubmed ↗
- 5Zhao J.Chen X.Herjan T.Li X.The role of interleukin-17 in tumor development and progression J Exp Med 21712020 e 2019029710.1084/jem.20190297 PMC 703724431727782 · doi ↗ · pubmed ↗