Integrative phylogenetic analysis of the genus Episoriculus (Mammalia: Eulipotyphla: Soricidae)
Yingxun Liu, Xuming Wang, Tao Wan, Rui Liao, Shunde Chen, Shaoying Liu, Bisong Yue, Branislav T. Šiler, Branislav T. Šiler, Ishtiyaq Ahmad, Ishtiyaq Ahmad

TL;DR
This study uses genetic and morphological data to clarify the taxonomy of the poorly understood shrew genus Episoriculus in China.
Contribution
The paper provides an integrative analysis confirming six valid species and revises the classification of several taxa within Episoriculus.
Findings
Six species of Episoriculus are recognized as valid, including E. soluensis, which is newly recorded in China.
E. sacratus is confirmed as a valid species, while E. umbrinus is reclassified as a subspecies of E. caudatus.
An undescribed Episoriculus taxon may exist in Xizang, suggesting further biodiversity in the region.
Abstract
Shrews in the genus Episoriculus are among the least-known mammals in China, where representatives occur mainly in the Himalayan and Hengduan mountains. We sequence one mitochondrial and three nuclear genes from 77 individuals referable to this genus, collect morphometric data for five shape and 11 skull measurements from 56 specimens, and use museum collections and GenBank sequences to analyze phylogenetic relationships between this and related genera in an integrated molecular and morphometric approach. Whereas historically anywhere from two to eight species have been recognized in this genus, we conclude that six (Episoriculus baileyi, E. caudatus, E. leucops, E. macrurus, E. sacratus, E. soluensis) are valid. We dissent from recent systematic reviews of this genus and regard E. sacratus to be a valid taxon, E. umbrinus to be a subspecies of E. caudatus, and transfer E. fumidus to…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
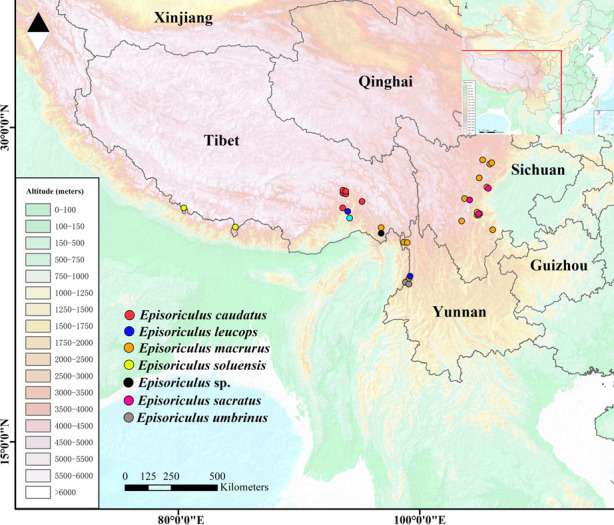 Fig 1
Fig 1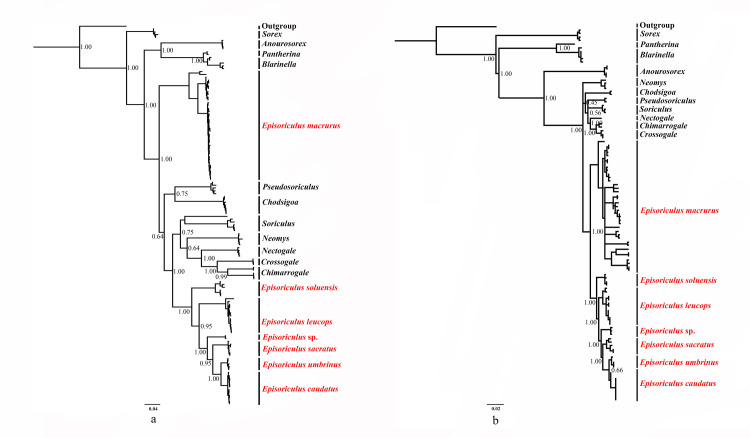 Fig 2
Fig 2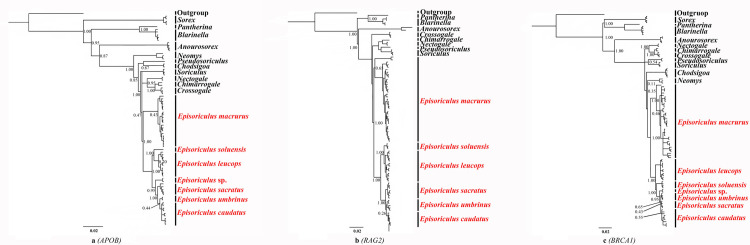 Fig 3
Fig 3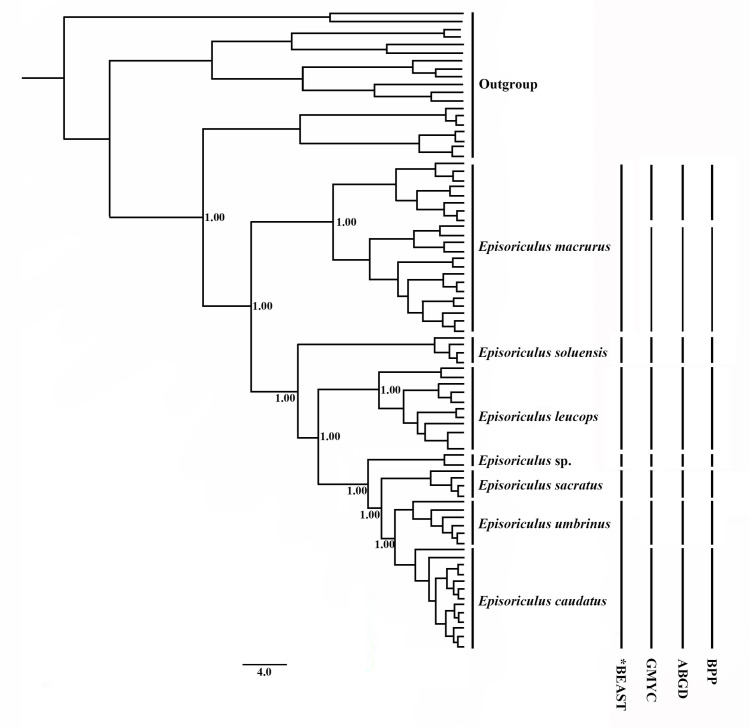 Fig 4
Fig 4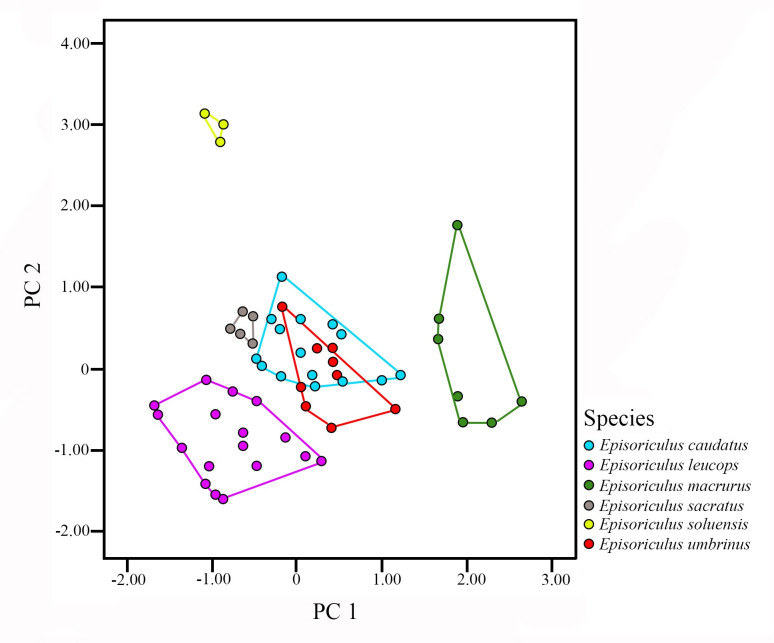 Fig 5
Fig 5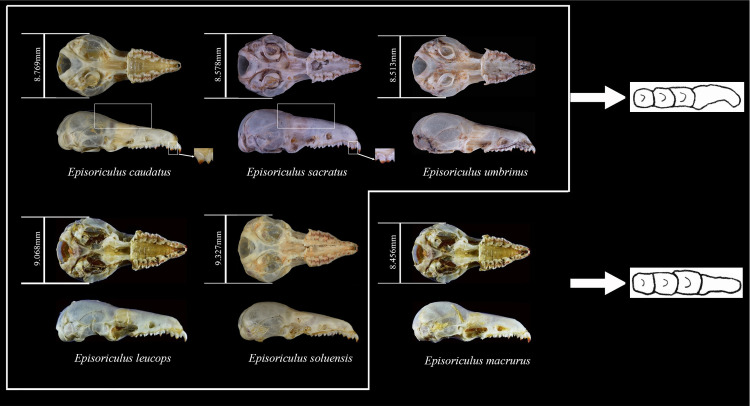 Fig 6
Fig 6- —http://dx.doi.org/10.13039/501100001809National Natural Science Foundation of China
- —http://dx.doi.org/10.13039/501100001809National Natural Science Foundation of China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEvolution and Paleontology Studies · Bat Biology and Ecology Studies · Animal Ecology and Behavior Studies
Introduction
The genus Episoriculus, originally established as a subgenus of Soriculus, occurs throughout southwest China, India, Nepal, and Vietnam [1–9]. This subgenus was assigned full generic status by Repenning [10] on grounds of significant differences in tooth morphology from other species of Chodsigoa and Soriculus—a taxonomy followed by Jameson and Jones [11], Hutterer [12], Wilson and Reeder [13], and Wilson and Mittermeier [3].
The number of valid species of Episoriculus has been the subject of debate, with 2–8 species recognized (Table 1). Allen [14] described Soriculus macrurus, S. caudatus sacratus, and S. caudatus umbrinus. Ellerman and Morrison-Scott [1] proposed Episoriculus as a subgenus of Soriculus, and included S. leucops and S. caudatus (with subspecies S. c. caudatus, S. c. baileyi, S. c. fumidus, S. c. sacratus, and S. c. umbrinus). Honacki et al. [6] proposed that Episoriculus included four species, and considered S. baileyi and S. fumidus to be valid taxa. Hoffmann [7] similarly recognized four species, although these were not entirely consistent with those of Honacki et al. [6], for S. baileyi was relegated to a subspecies of S. (E.) leucops, and S. (E.) macrurus was placed in this genus. Corbet and Hill [8], and Wilson and Reeder [9, 13] followed this arrangement. Motokawa and Lin [15] elevated S. baileyi to full species based on morphology. Based on the karyotypes and differences in skull morphology, Motokawa et al. [16] considered that Episoriculus caudatus should be divided into the larger E. caudatus and smaller E. sacratus (with subspecies E. s. soluensis from Nepal and Sikkim, E. s. umbrinus from Assam, Myanmar, and Yunnan, China, and E. s. sacratus from Sichuan, China). He et al. [17] noted that Pseudosoriculus fumidus did not belong to Episoriculus. Based on CYTB gene sequences, Abramov et al. [2] promoted E. soluensis to full species, assigned E. fumidus to a new genus Pseudosoriculus, and arranged seven species (E. baileyi, E. caudatus, E. leucops, E. macrurus, E. sacratus, E. soluensis, and E. umbrinus) in Episoriculus. Wilson and Mittermeier [3] recognized eight species, including P. fumidus.
Table 1: Major classification systems of the genus Episoriculus.A name of the species in parentheses indicates that the taxon is a subspecies of the previous species.
Throughout these various classifications the taxonomic status of E. caudatus, E. leucops, and E. macrurus has been relatively stable, but the taxonomy of P. fumidus, E. sacratus, E. umbrinus, E. baileyi, and E. soluensis has not. We report new molecular data and morphological comparisons in an integrated phylogenetic and morphological analysis to clarify the taxonomic status of species in the genus Episoriculus.
Materials and methods
Field methodology
In the study, traplines were installed to capture shrews. We use red plastic buckets as traps. Each of these 7L buckets was 22 cm high and had an upper and lower diameter of 26 cm and 19 cm, respectively. We typically position these barrels in the more humid forest, primarily around the roots of massive, or falling trees, 3-5m apart. The buckets were often put in the afternoon of the first day and checked the following morning to see whether any shrews have fallen into the trap. The time between placement and inspection is assured to be more than 12 hours.
After euthanasia with eugenol, following the ASM guidelines [18], tissue samples were taken from the thigh muscle and stored in absolute ethyl alcohol at ambient temperature. Voucher specimens fixed in 10% formalin before transferal to 95% ethanol for long term preservation and deposition of vouchers at the Zoological Museum of Sichuan Academy of Forestry (SAF), Chengdu, Sichuan, China. All specimens were collected in accordance with regulations in China for implementation of the protection of terrestrial wild animals (State Council Decree [1992] No. 13). Collecting protocols and the research project were approved by the Ethics Committee of Sichuan Academy of Forestry (2024–001).
Sampling and sequencing
All specimens were identified morphologically following the original literature species established [19–23], and some subsequent studies on this group, such as Wilson and Mittermeier [3], Hoffmann [7], and Smith and Xie [24] in which, some specimens which were be identified as E. sacratus and E. soluensis, were collected adjacent of the type locality (Table 2). For verifying our species accuracies, some sequences collected form type locality or adjacent of the type locality, were downloaded [2, 17, 25–32] (Table 3). Of recognized species, we had no specimens or sequences of E. baileyi. Through preliminary molecular and morphological identification, 77 specimens collected from China, 31 were attributed to E. macrurus, 18 to E. caudatus, 10 to E. leucops, 5 to E. umbrinus, 4 of each to E. sacratus and E. soluensis, and 2 to E. sp. (Table 1 and Fig 1)
Map of genus Episoriculus, showing localities sampled for this study.Reprinted from Fan et al. (2022) [Doi: https://doi.org/10.1002/ece3.9404] under a CC BY license, with permission from Fan Ronghui, original copyright 2022. And edited and supplemented by Adobe Photoshop 2022 and Arcgis 10.8.
Table 2: Samples and sequences of Episoriculus used for molecular analyses.
Table 3: GenBank accession numbers of download sequence from NCBI.
Following He et al. [17] and Chen et al. [32, 33], we amplified the complete mitochondrial cytochrome b (CYTB) and three partial nuclear genes (apolipoprotein B (APOB), recombination-activating gene 2 (RAG2), and breast cancer 1 (BRCA1)). Primer sets are detailed in Table 4 [17, 30, 34, 35]. PCR amplifications were carried out in a 25 μl reaction volume mixture containing 12.5 μl of 2×Taq Master Mix (Vazyme, Nanjing, China), 1 μl of each primer, 1 μl of genomic DNA, and 9.5 μl of double-distilled water. PCR conditions for CYTB amplifications consisted of an initial denaturing step at 94°C for 5 min followed by 38 cycles of denaturation at 94°C for 45 s, annealing at 49°C for 45 s, an extension at 72°C for 90 s, and a final extension step at 72°C for 12 min. PCR conditions for nuclear genes were basically the same as those for CYTB, with a few modifications (annealing temperatures for each nuclear gene were APOB (49°C), BRCA1 (51°C), and RAG2 (52°C)). PCR products were checked on a 1.0% agarose gel and purified by ethanol precipitation. Purified PCR products were directly sequenced using the BigDye Terminator Cycle Kit v 3.1 (Applied Biosystems, Foster City, CA, USA) and an ABI 310 Analyzer (Applied Biosystems).
Table 4: Gene symbol, primer sequences and the best model of evolution for each gene segments used in the study.
To test phylogenetic relationships within the genus Episoriculus, sequences of these four genes from other Nectogalini species generated in previous studies [2, 17, 25–32] were downloaded from GenBank (Table 2).
Sequence analyses
All CYTB sequences were aligned and examined. Screening for heterozygous nuclear gene fragments was performed in Mega 5 [36]. For analysis, we concatenated the three nuclear genes following He et al. [17]. Using all sequence data, phylogenetic analyses were conducted on: 1) CYTB data, 2) concatenated sequences for the three nuclear genes, and 3) each nuclear gene. Modeltest v 3.7 [37] was used to select the best-fitting evolutionary model, based on the Akaike Information Criterion in Table 3. MrBayes v 3.1.2 [38] was used for Bayesian analysis. Crocidura fuliginosa was selected as the outgroup. Each run was carried out with four Monte Carlo Markov chains (MCMCs), and 10,000,000 generations for single gene datasets and 30,000,000 generations for concatenated gene datasets. All runs were sampled every 10,000 generations. Convergences of runs were accepted when the average standard deviation of split frequencies was < 0.01. Ultrafast bootstrap values (UFBoot) of ≥ 95 and posterior probabilities (PP) of ≥ 0.95 were considered strong support [39].
Species tree and species delimitation
To appraise current taxonomic systems based on morphology, and to evaluate trees derived from phylogenetic analyses, we used species delimitation and species tree construction based on coalescence theory [40, 41]. The basis of abductive theory-based species definition is to use the multispecies coalescent model to identify evolutionarily independent lineages in genomic data. which is significantly distinct from defining species as stable differences between monophyletic groups or taxa [42]. Since the *BEAST model requires that every gene segment in each sample be complete, we didn’t use all samples. We estimated the *BEAST coalescent species tree in Beast v 1.7.5 using partial nuclear and mitochondrial genes [43, 44]. Model settings were selected with reference to the optimal replacement model of each gene (Table 3). Each analysis was run for 100 ×10^6^ generations and sampled every 10,000 generations. Posterior distribution and effective sample size of each parameter were calculated using Tracer v 1.6. *BEAST analyses were repeated four times to ensure convergence on the same posterior distribution.
“Splits” (Species Limits by Threshold Statistics) v 1.0 14 was used for species delimitation in the context of R statistics. The program defines species using the generalized mixed Yule-coalescent model (GMYC) [45, 46]. Analysis requires a gene tree that has been corrected by a molecular clock as a reference; we use the bifurcated time tree constructed with Beast v 1.7.5 as basic data for this analysis.
BPP v 2.2 was used to species delimitation [47, 48]. Analyses of species boundaries were limited to E. caudatus, E. sacratus, and E. umbrinus; analyses used the combined nuclear gene data set. Since this program also requires a prior guide tree, and the topological structure of the guide tree will impact the result of species delimitation, we used the aforementioned species tree built by BEAST v 1.7.5 as the guide tree for BPP analysis [49]. For BPP, we set the Gamma prior distribution of the population size parameter (θs) to G (6, 6,000), and the initial differentiation time parameter (τ0) of the species tree to G (4, 1,000). Then 12 parameter combinations were generated using algorithms 0 and 1, and combining the values of Locusrate = 1 or Heredity = 1. The above 12 operations were performed on the two data groups; each operation was set to 1,000,000 generations of reverse-jump MCMC, and samples were taken every 10 generations; the first 10,000 generations were discarded [50].
Automatic barcode gap discovery (ABGD) software was used to divide samples based on genetic distance; samples within the same group were identified as one species [51]. CYTB sequences of the sample online submission to ABGD website (http://wwwabi.snv.Jussleu.fr/publicabgd/abgdweb.hml), the prior intraspecific divergence (P) ranged 0.001–0.1, and the minimum relative gap width (X) was 0.5.
We also used the Kimura 2-parameter (K2p) distance with 10,000 bootstrap replicates to summarize sequence divergences based on CYTB in MEGA5 [36].
Morphometrics
Because we have found no evidence for sexual dimorphism in shrews in related taxa, we do not consider sex when selecting specimens and skulls [52]. All samples used for analysis were adults with intact skulls. Specimens of Episoriculus used for study have been deposited in the Sichuan Academy of Forestry, and Kunming Institute of Zoology. A total of 56 complete skulls of adult intact specimens were assigned to E. macrurus (17), E. caudatus (11), E. umbrinus (8), E. leucops (7), E. sacratus (6), and E. soluensis (3). Details of localities and museums are listed in S1 Table.
Measurements including head and body length (HBL), tail length (TL), hind foot length (HFL), and ear length (EL) were recorded from specimen labels or field notes. We measured the skulls of these specimens with a digital Vernier caliper (accuracy 0.01 mm). Eleven craniomandibular variables were taken: profile length (PL), cranial height (CH), cranial breadth (CB), interorbital breadth (IBO), palatoincisive length (PIL), postpalatal length (PPL), maxillary breadth (MB), upper toothrow length (UTR), maximum width across the upper second molars (M^2^–M^2^), mandibular length (ML), and lower toothrow length (LTR). Measurements dates were used for principal component analysis (PCA) in SPSS 22.0 (SPSS Inc., USA). Sample localities and measurements for each specimen are presented in S1 Table. Measuring methods followed Chen et al. [25], Yang et al. [53].
Cranial measurements were analyzed by PCA in SPSS v19.0 (SPSS Inc., USA). PCAs were conducted on log_10_ transformed variables on two data sets. Before analysis, the Kaiser–Meyer–Olkin (KMO) test (to check correlations or partial correlations between variables), and Bartlett’s sphere test (to determine if the correlation matrix is an identity matrix) were performed.
Results
Phylogenetic analysis
We obtained 69 mitochondrial sequences, and 156 nuclear sequences. All CYTB, APOB. BRCA1 and RAG2 files are available in GenBank (GenBank Accession No.: MK962157-MK962237, MN032125-MN032174, MN032175-MN032241, and MN032242-MN032310) (Table 2).
Bayesian reconstruction using CYTB revealed eight monophyletic clades of Episoriculus, corresponding to E. macrurus, E. soluensis, E. leucops, E. caudatus, E. umbrinus, E. sacratus, and E. sp. (Fig 2A). P. fumidus clustered with Chodsigoa with strong support (PP = 0.95). E. macrurus represented a basal lineage of Nectogalini with strong support (PP = 1.00), and E. soluensis, E. leucops, E. sacratus and E. sp. formed a separate, strongly supported lineage (PP = 0.95–1.00). E. umbrinus and E. caudatus, located at the tip of this tree, formed sister clades, that were less-well supported (PP = 0.75).
Bayesian phylogenetic analyses based on (a) CYTB, and (b) three concatenated nuclear genes. Numbers at nodes refer to Bayesian posterior probabilities. Scale bars represent substitutions per site.
Bayesian reconstruction using the three concatenated nuclear genes also revealed eight monophyletic clades (Fig 2B), but with a slightly different topological structure to the CYTB tree. P. fumidus mixed with Chodsigoa and Soriculus, with weak support. All species of Episoriculus formed a monophyletic clade, with E. macrurus at its base. Lineages of E. soluensis, E. leucops, E. sacratus, and E. sp. were strongly supported (PP = 1.00). E. umbrinus and E. caudatus formed sister clades at the tree’s tip, although support for them was weak (PP = 0.66).
Structures of three individual nuclear gene trees differed from the CYTB and three concatenated nuclear gene trees (Fig 3), with some nodes having very low support. P. fumidus did not cluster with Episoriculus. E. macrurus, E. soluensis, E. leucops, E. sp., and E. sacratus remained monophyletic with strong support based on APOB and RAG2 (PP = 1.00) genes, and E. umbrinus and E. caudatus formed a sister group with weak support. E. sacratus, E. umbrinus and E. caudatus were mixed on the tree based on the BRCA1 gene, with very low support.
Bayesian phylogenetic analyses based on (a) APOB, (b) RAG2, and (c) BRCA1 genes. Numbers at nodes refer to Bayesian posterior probabilities. Scale bars represent substitutions per site.
Species delimitation
The topology of *BEAST species’ trees differed slightly from those of mitochondrial and nuclear genes (Fig 4). E. macrurus, E. soluensis, E. leucops, E. sacratus, and E. sp. also had high support in these trees (PP = 1.00). E. umbrinus and E. caudatus were sister clades, but with weak support in the tree (PP = 0.75). This result did not support recognizing E. umbrinus as a distinct species.
*Results of species delimitation using splits, GMYC, BPP, and species trees reconstructed using the BEAST model.Node numbers indicate Bayesian posterior probabilities supporting each clade as two putative species.
GMYC analysis reveals five clades as valid species (Fig 4), of which E. soluensis, E. leucops, and E. sp. are separate, and E. macrurus comprised two species, with individuals from Sichuan and Yunnan provinces differing. This analysis suggests that E. caudatus, E. sacratus and E. umbrinus are conspecific.
For the ABGD analysis, the transition/transversion value (3.5) first calculated by Mega 5 was used as the starting parameter, with 0.5, 1.0, 1.5 and 2.0 used as relative gap widths. The 81 samples divided into eight species: E. caudatus, E. macrurus, E. sacratus, E. soluensis, E. leucops, E. macrurus (Sichuan samples), E. macrurus (Yunnan samples), and E. sp. (Fig 4 and S2 Table).
BPP analysis revealed E. caudatus and E. umbrinus to be separate species in 12 groups of BPP data based on the combined nuclear gene data set, with support for E. sacratus being a valid taxon also being high (Fig 4 and S3 Table).
Kimura-2-parameter (K2p) distances between Episoriculus species ranged 0.027–0.160 (Table 5). The average K2p distance between P. fumidus and Episoriculus species was 0.177. The K2p distance between E. caudatus and E. sacratus was 0.067, and between E. caudatus and E. umbrinus, 0.027, and E. sacratus and E. umbrinus, 0.071.
Table 5: The Kimura-2-parameter distances between Episoriculus species based on the CYTB gene.
Morphology
Morphological data (HBL, TL, HFL, EL, and body weight (BW)) of 56 specimens with intact skulls are presented in Table 6. The TL of E. macrurus ranged 71–106mm, while values for congeners ranged 46–82.5 mm. The HFL of E. macrurus was 15–16 mm, while for other species it was 12–13 mm. E. macrurus had the lowest HBL/TL ratio (0.64), and its TL was ~1.5 times its HBL, while the HBL of congeners was approximately equal to or greater than TL. E. leucops had the largest body, and E. sacratus differed from E. caudatus and E. umbrinus in HBL/TL values. Morphological indices are detailed in Table 7, and complete measurement data are provided in S1 Table.
Table 6: The results of body morphologic measurements.
Table 7: Morphological measurement data of Episoriculus species skulls.
Bartlett’s test rejected the null hypothesis (χ^2^ = 568.01, P = 0.000), indicating that the data were spherical and variables were somewhat independent of each other. A KMO of 0.813 indicated a strong correlation existed among the various skull data, which was suitable for factor analysis. Two principal components explaining 74.69% of morphological variation were extracted from the analysis. Factor loading values were most positive, indicating that it was mainly related to overall skull size (Table 8). Features with factor loads > 0.8 included PIL, ML, CIL, UTR, and LTR.
Table 8: Character loadings, eigenvalues, and proportion of variance explained by the first two axes (PC 1 and PC 2) of a principal component analysis using the log10-transformed measurements of Episoriculus.The meanings of variable abbreviations are given in the Materials and Methods Section.
Using PC 1 and PC 2 maps (Fig 5), E. macrurus plotted in the positive region of PC 2, while other species were mainly in the negative region of PC 2. The larger E. leucops plotted in the negative region of PC 2, and E. soluensis plotted in the positive region of PC 2. E. sacratus was distinguished with E. umbrinus and E. caudatus, with the latter two species being mixed and not effectively differentiated.
Results of principal component analysis of Episoriculus taxa based on 19 log10 transformed craniodental measurements.
Compared to skulls of other Episoriculus species (Fig 6), the braincase of E. macrurus is more dome-shaped, the rostrum is shorter; and the upper unicuspids are quadrangular and wider than long (those of other species are similarly sized). Compared to skulls of E. caudatus, the frontal region of the skull of E. sacratus is more arched, and the posterior cusp of its upper incisor is lower than its first unicuspid, whereas the height of the posterior cusp of the upper incisor and first unicuspid of E. caudatus are similar.
Comparison of Episoriculus skulls.
Discussion
In the study, we collected a huge number of shrews in China and, via morphological and molecular analysis, concluded that genus Episoriculus contains six distinct species and one cryptic species. The majority of Episoriculus species have their type origin in India. We further validated the accuracy of our species classification by utilizing sequence collected in India from Ohdachi et al. [29]. We did comprehensive studies on some species whose classification is contested.
Thomas [54] described six specimens from Mount Emei as E. sacratus, and considered it most likely the local representative of E. caudatus, from which they differed in having a much smaller braincase. Allen [55] described the subspecies E. caudatus umbrinus from Mucheng, Yunnan, which was most alike E. sacratus, but differed from it in its much darker-brown color and in having a uniformly dark rather than bicolor tail. Allen [14] demoted E. sacratus to a subspecies of E. caudatus—an opinion with which Ellerman and Morrison-Scott [1] agreed. Hoffmann [7] examined many specimens and concluded that there were similarities in skull and cranial size between E. c. caudatus, E. c. sacratus and E. c. umbrinus. Wilson and Reeder [9, 13] recognized these three taxa to be distinct species based on karyotypes and differences in skull size, while Motokawa and Lin [15] considered E. sacratus and E. umbrinus to be subspecies of E. caudatus. Motowaka et al. [16] considered the larger E. caudatus and smaller E. sacratus to be distinct species, and included three subspecies: E. soluensis, E. umbrinus, and E. sacratus as subspecies of E. caudatus. Wilson and Mittermeier [3] elevated E. umbrinus to full species without reason, while Wei et al. [56] regarded it to be a subspecies of E. caudatus. Our data support recognizing E. sacratus as a valid species, and recognizing E. umbrinus to be a subspecies of E. caudatus. In morphology, E. sacratus can be differentiated from E. caudatus, but E. umbrinus cannot. Our data do not support the opinion of Motowaka et al. [16]. For the two species, we considered that the Jinsha River and Hengduan Mountains shut off communication between E sacratus and E. caudatus, limiting the species to the edge of the western Sichuan Plateau. And E. caudatus is widely distributed in northeastern India, northern Burma, northern Vietnam, and Xizang, Yunnan, Guizhou, China [2, 3, 17, 57].
Gruber [23] described E. soluensis and considered it a separate species. Abe [58] reviewed specimens collected in central Nepal and compared them with those from eastern Nepal by Gruber [23], and both suggested that E. soluensis was a subspecies of E. caudatus, but also possibly synonymous with E. sacratus. Hoffmann [7] treated E. soluensis as a synonym of E. caudatus—an opinion with which Wilson and Reeder [9, 13] and Motokawa and Lin [15] agreed. Ohdachi et al. [29] regarded two samples from Nepal to be E. caudatus soluensis following Abe [58], and sequenced CYTB. Abramov et al. [2] then used these two sequences to reconstruct a system tree, and after determining that E. soluensis constituted a distinct clade from E. caudatus, advocated for them being treated as distinct species. In our tree, four samples from Yadong and Nyalam cluster with the two E. soluensis samples of Ohdachi et al. [29], and these four specimens are similar in having dark-brown ventral hair, and light-yellowish-brown dorsal hair. The tail length of our four specimens is longer than the head length, the skull parietal bone is relatively protruding, there are four upper single cusp teeth, the posterior cusp teeth of the maxillary incisor are similar in height to the first upper single cusp teeth, and the cusp teeth are light brown. These features are basically consistent with Gruber’s [23] original description, and the description of E. soluensis of Wilson and Mittermeier [3]. Accordingly, we regard E. soluensis to be a distinct species, and report it for the first time from China. And We discovered that this species lives on both sides of the middle Himalayas (Nepal, northeast India, and Shigatse region, China).
While Ellerman and Morrison-Scott [1] regarded Episoriculus baileyi to be a subspecies of E. caudatus, Abe [58, 59] identified the two to be morphologically distinct and sympatric in Nepal. Hoffmann [7] examined species from Burma and Nepal and considered E. baileyi to be a subspecies of E. leucops, an opinion with which Wilson and Reeder [9, 13] agreed. Based on external and cranial morphology, Motokawa and Lin [15] re-evaluated the taxonomic status of E. baileyi, and considered it to be a valid species of Episoriculus. This species could be distinguished from other Episoriculus in the combination of its robust first upper incisor, long rostrum and upper unicuspid row, large tympanic ring, and high ascending ramus of the mandible—an opinion with which Wilson and Mittermeier [3] agreed. Because of a lack of specimens, we cannot investigate the status of E. baileyi. We provisionally follow Motokawa and Lin [15], but the taxonomic status of this species requires further investigation.
While Ellerman and Morrison-Scott [1] considered E. fumidus to be a subspecies of E. caudatus, Jameson and Jones [11] considered it to be a distinct species based on its geographical isolation and morphological divergence—an opinion with which Hoffmann [7], Wilson and Reeder [9, 13], and Motokawa and Lin [15] agreed. Dubey et al. [29] inferred that E. fumidus (the only representative of the genus in their study) was a sister group of Chodsigoa with strong support in the APOB gene tree—an opinion with which He et al. [17] agreed. Based on the sequence of Dubey et al. [29] and He et al. [17], Abramov et al. [2] regarded fumidus do not belong in Episoriculus, and established the genus Pseudosoriculus for it—an opinion supported by our analyses.
We identify what appears to be a new cryptic species (E. sp.) from low-elevation areas in Motuo County, Xizang, which forms a separate branch in our system tree. However, with only two specimens available, we cannot accurately describe its morphology. Further specimens and molecular data are required to accurately resolve the taxonomic status of this taxon.
Our phylogenic analysis consistently roots E. macrurus as an individual lineage. This species, which has large genetic distance from congeners, in phylogenetic trees is usually located in the outermost or most basal part of the genus Episoriculus. It has the longest tail in the genus, and differs from congeners in skull and tooth morphology. For these reasons we speculate it retains some of the most primitive traits in genus Episoriculus or Nectogalini. However, in phylogenetic trees based on different genes, the position of a varies: in the phylogenetic tree based on mitochondrial gene, E. macrurus is at the base of tribe Nectogalini and forms a single monophyletic group, whereas in the nuclear gene-based tree, E. macrurus is included in Nectogalini and clustered with other species of Episoriculus on the same clade. Similar conflicting phylogenetic signals have been reported in other studies [60]. This phenomenon could be explained from the perspectives of genetic background [61], ancient hybridization [62], incomplete lineage sorting [63], adaptive evolution, and burst Formula speciation [64]. The long tail and developed hind feet of this species lend it a semi-arboreal appearance. This specific niche adaption, which is not shared by other species, may result in huge differences in energy metabolism patterns between E. macrurus and other species of Nectogalini. This was reflected in mitochondria, which caused early differentiation in the species tree built using mitochondrial genes, resulting in incomplete lineage sorting. While neither mtDNA nor nDNA alone resolved phylogenetic relationships in the genus Episoriculus, combining data from these two genetic pathways did improve results. Species tree construction in the coalescent framework also produced a consistent topology with high statistical support. Therefore, we deem that a combined approach using mitochondrial and nuclear gene information is more appropriate for resolving phylogenetic relationships in the genus Episoriculus.
Conclusion
Based on molecular and morphological analyses, the genus Episoriculus comprises at least six valid species: E. baileyi, E. caudatus, E. leucops, E. macrurus, E. sacratus, E. soluensis, and the potentially undescribed E. sp.
Supporting information
S1 TableExternal and selected cranial measurements of Episoriculus species.(DOCX)
S2 TableResults of ABGD species definition based on CYTB gene.(DOCX)
S3 TablePosterior probabilities supporting three species (Episoriculus caudatus, E. sacratus, and E. umbrinus) as potential species using different algorithms and priors.(DOCX)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abramov AV, Bannikova AA, Chernetskaya DM, Lebedev VS, Rozhnov VV. The first record of Episoriculus umbrinus from Vietnam, with notes on the taxonomic composition of Episoriculus (Mammalia, Soricidae). Russian Journal of Theriology. 2017; 16(2):117–128. 10.15298/rusjtheriol.16.2.01. · doi ↗
- 2Wilson DE, Mittermeier RA. Handbook of the Mammals of the World–Volume 8: Insectivores, Sloths and Colugos. Barcelona: Lynx Edicions; 2018.
- 3Walker EP, Warnick F, Hamlet SE, Lange KI, Davis MA, Uible HE, et al. Mammals of the world. Third ed. Baltimore: Johns Hopkins University Press; 1975.
- 4Corbet GB. The mammals of the Palaearctic region: A taxonomic review. London: British Museum; 1978 c.
- 5Honacki JH, Kinman KE, Koeppl JW. Mammal species of the world. Laurence, Kansas: Allen Press, Inc. and Association of Systematic Collections; 1982.
- 6Hoffmann RS. A review of the genus Soriculus (Mammalia: Insectivora). Journal of the Bombay Natural History Society. 1985; 82: 459–481.
- 7Corbet GB, Hill JE. Mammals of the Indomalayan region. A systematic review. Oxford: Oxford University Press; 1992.
- 8Repenning CA. Subfamilies and genera of the Soricidae. Geological Survey Professional Paper. 1967; 565:1–74.