Carbon source, cell density, and the microbial community control inhibition of V. cholerae surface colonization by environmental nitrate
Jamaurie James, Renato E.R.S. Santos, Paula I. Watnick

TL;DR
The study shows how nitrate and nitrite affect Vibrio cholerae's ability to form biofilms in the gut, and how other microbes can help it colonize better.
Contribution
The novel finding is that nitrite-reducing microbes like Paracoccus aminovorans can rescue V. cholerae biofilm formation in the presence of nitrate.
Findings
V. cholerae nitrate respiration leads to nitrite accumulation, which inhibits biofilm formation.
Paracoccus aminovorans metabolizes nitrite, rescuing V. cholerae surface accumulation in co-cultures.
Intestinal microbiota composition influences V. cholerae colonization and cholera susceptibility.
Abstract
The intestinal diarrheal pathogen Vibrio cholerae colonizes the host terminal ileum, a microaerophilic, glucose-poor, nitrate-rich environment. In this environment, V. cholerae respires nitrate and increases transport and utilization of alternative carbon sources via the cAMP receptor protein (CRP), a transcription factor that is active during glucose scarcity. Here, we show that V. cholerae nitrate respiration in aerated cultures is under control of CRP and, therefore, glucose availability. V. cholerae nitrate respiration results in extracellular accumulation of nitrite because V. cholerae does not possess the machinery for nitrite reduction. This nitrite inhibits V. cholerae biofilm formation via an as-yet unelucidated mechanism that depends on the high cell density master regulator HapR. The genome of Paracoccus aminovorans, an intestinal microbe identified in the microbiome of…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
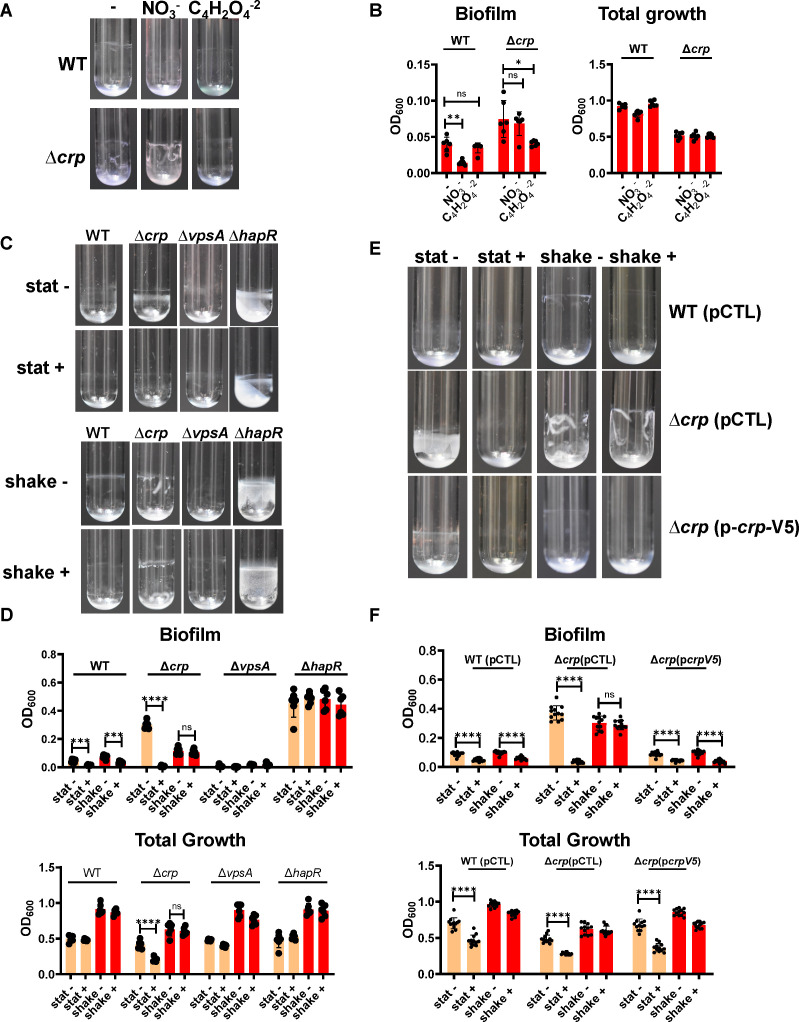 Fig 1
Fig 1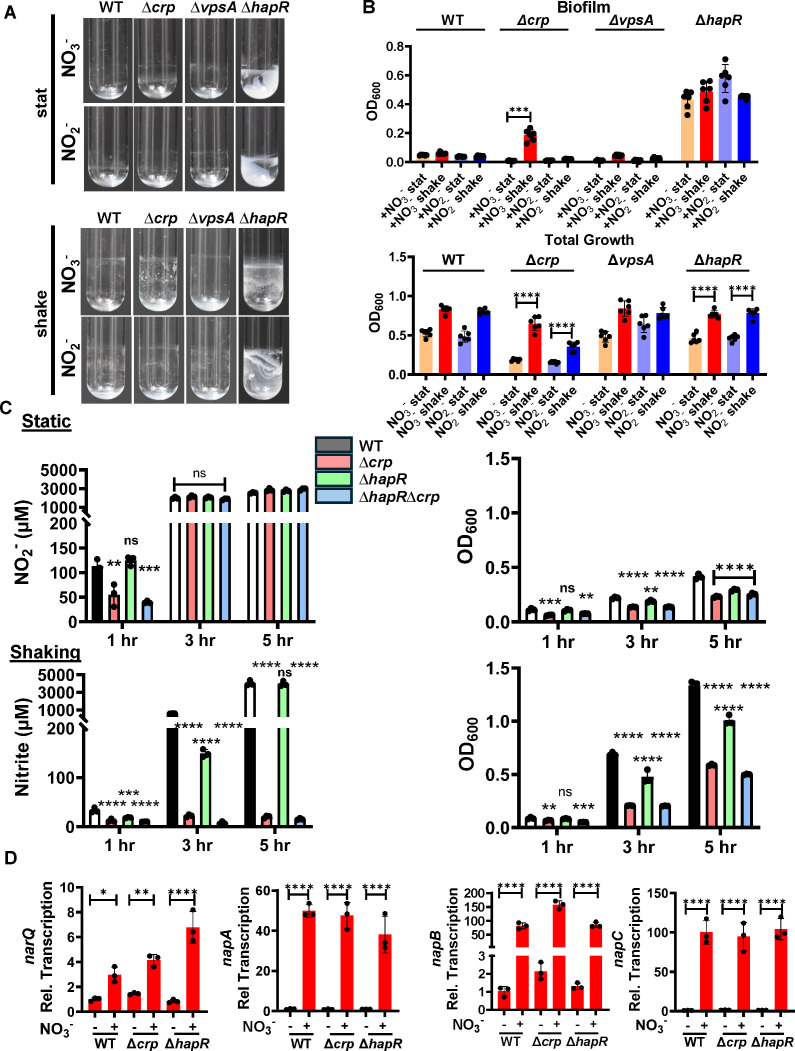 Fig 2
Fig 2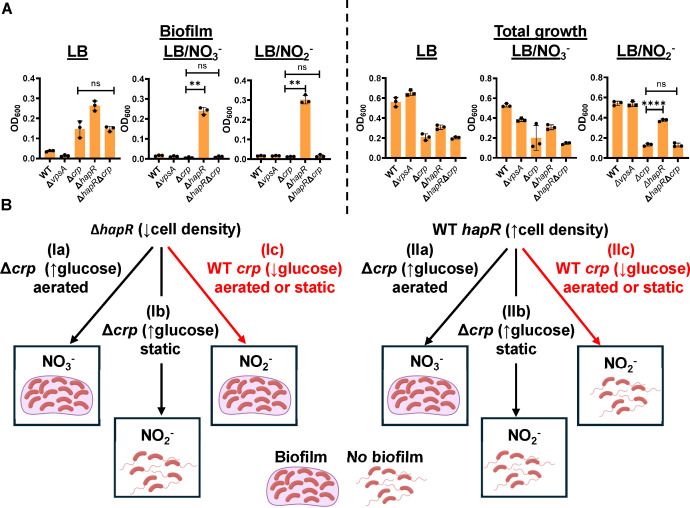 Fig 3
Fig 3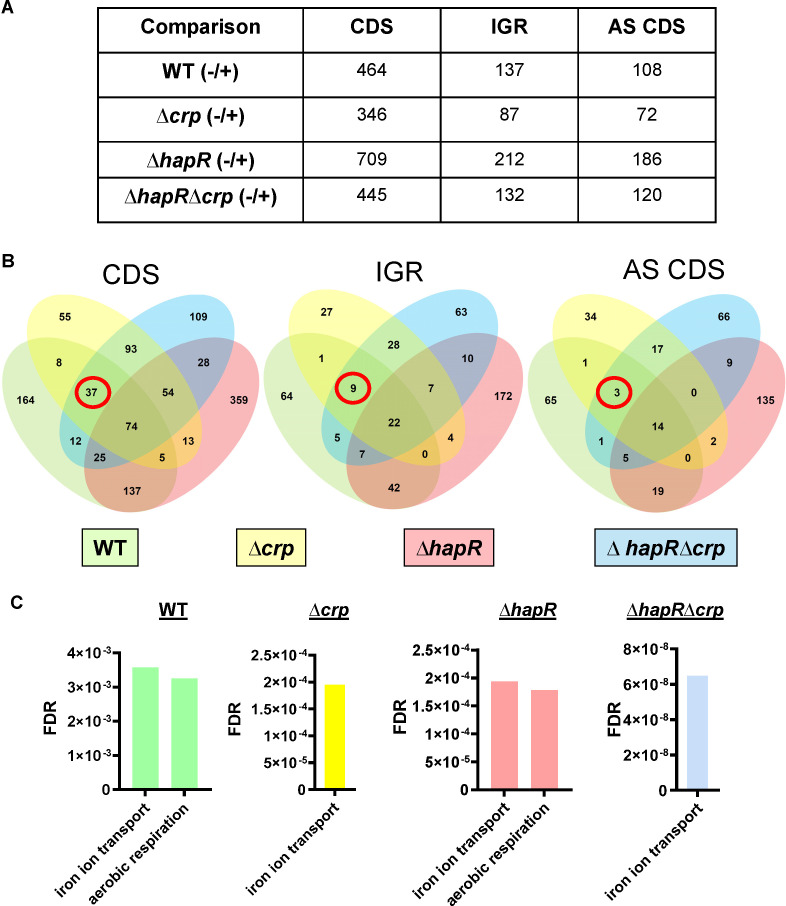 Fig 4
Fig 4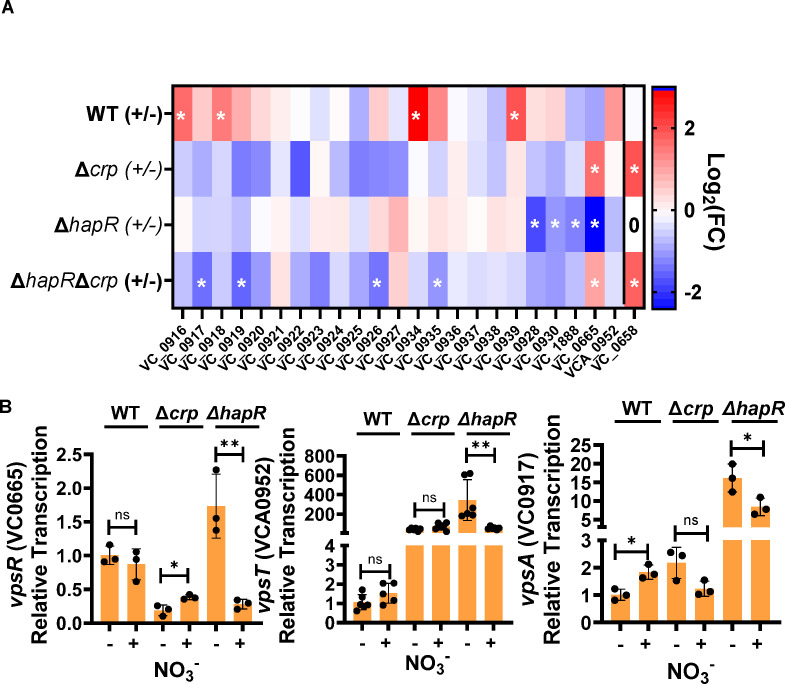 Fig 5
Fig 5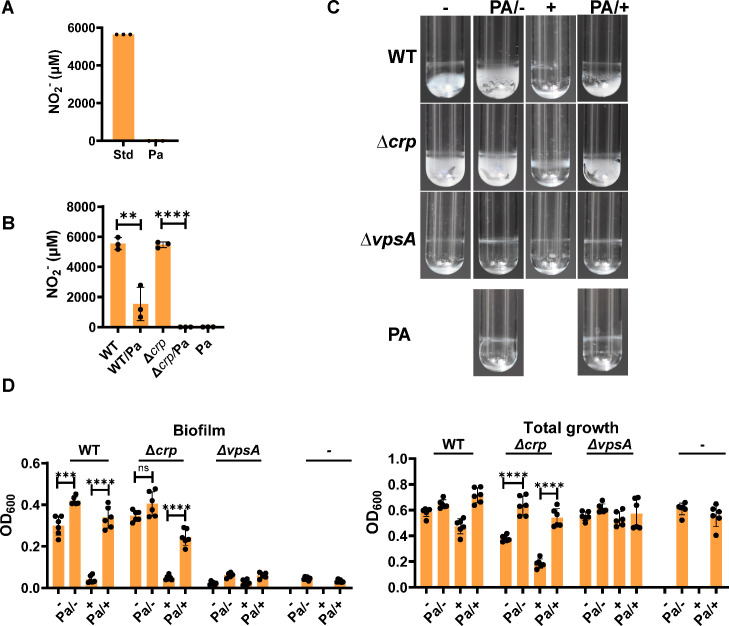 Fig 6
Fig 6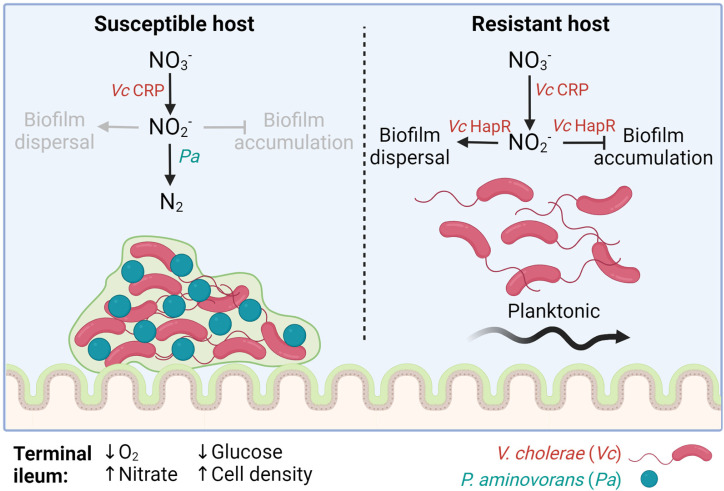 Fig 7
Fig 7- —HHS | NIH | National Institute of Allergy and Infectious Diseases (NIAID)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsVibrio bacteria research studies · Aquaculture disease management and microbiota · Antibiotic Resistance in Bacteria
INTRODUCTION
The gram-negative pathogen Vibrio cholerae is a facultative anaerobe that colonizes the ileum to cause the acute and severe diarrheal disease cholera (1). In an infant mouse model, the principal V. cholerae attachment factor is the type IV toxin co-regulated pilus (2). In contrast, on environmental surfaces, V. cholerae constructs a three-dimensional surface-attached structure known as the Vibrio polysaccharide (VPS)-dependent biofilm by synthesizing a VPS exopolysaccharide scaffold via enzymes encoded by the vps genes as well as the three secreted lectins RbmA, RbmC, and Bap1 (3–6). After secretion, these lectins bind to the VPS exopolysaccharide and form bridges between neighboring cells and between cells and the surface to erect a stable, three-dimensional structure. There is evidence from infant mouse studies that this VPS-dependent biofilm increases small bowel colonization in vivo when members of the human intestinal microbiota that are associated with cholera are present (7–10).
V. cholerae is found in the small and large bowel of the infant mouse during infection, but only in the large bowel of germ-free or streptomycin-treated adult mice (11, 12). Large bowel colonization is asymptomatic in mice and independent of the toxin co-regulated pilus, a principal colonization factor (13). Asymptomatic carriage of V. cholerae also occurs in humans and may reflect large bowel colonization (14, 15). This suggests that the conditions encountered by V. cholerae in the small intestine are essential for activation of virulence determinants and development of disease.
Availability of the terminal electron acceptors oxygen and nitrate varies along the length of the intestine, and these modulate virulence behaviors in pathogens other than V. cholerae (16–22). The partial pressure of oxygen is highest in the duodenum at approximately 60 mmHg and decreases to under 10 mmHg in the ileum (22). Despite the lower oxygen concentrations, V. cholerae colonization of the small bowel relies heavily on aerobic respiration that utilizes the terminal oxidases cytochrome bd-type ubiquinol oxidase I (bd-1) and the heme copper oxidase Cbb3 (23, 24).
Nitrate, which is generated through the actions of iNOS and NOX1, is present in the adult mouse ileum at concentrations approaching 6 mM and at much lower concentrations in other regions of the small intestine (20, 25).
V. cholerae’s use of nitrate as a terminal electron acceptor is limited. While other bacterial genomes encode multiple systems for nitrate reduction to nitrite, the V. cholerae genome harbors only a periplasmic nitrate reductase encoded by the napDABC genes (24, 26). Other bacteria are able to further reduce nitrite to nitrogen gas via a pathway comprised of multiple reductive enzymes that carry out the following reactions: nitrite (NO_2_^-^)→nitric oxide (NO)→nitrous oxide (N_2_O)→nitrogen gas (N_2_) (27). In contrast, V. cholerae reduces nitrate to nitrite but cannot carry out additional reductions (28). This results in accumulation of conditionally toxic nitrite in the extracellular environment (29).
The V. cholerae genes required for nitrate respiration under hypoxic laboratory conditions have been exhaustively identified by Tnseq and include the nap genes, the nqr genes, which encode the Na+-translocating NADH-quinone reductase, and the ccm cytochrome maturation genes (30). These authors found a small but significant colonization defect for a ∆napA mutant in a streptomycin-treated adult mouse model, although colonization was quantified in the caecum and colon only (29, 30). A microarray analysis of V. cholerae in human cholera stool found that the nap genes were highly expressed as compared with laboratory conditions (31). However, expression varied greatly between the three patients studied, suggesting significant effects of the intestinal environment.
We hypothesized that nitrate reduction might play an important role in V. cholerae colonization of the terminal ileum. Because this environment contains oxygen, we began by defining the regulation of V. cholerae nitrate reduction in aerobic cultures and its effect on surface colonization. Here, we describe an unusual role for two transcription factors, the cAMP receptor protein (CRP) and HapR in the regulation of V. cholerae nitrate reduction to nitrite and the inhibition of V. cholerae biofilm formation by nitrite, respectively.
CRP is a conserved global regulator of carbon metabolism that binds DNA in the presence of the second messenger cyclic adenosine monophosphate (cAMP) (32). cAMP is synthesized by the enzyme adenylate cyclase. When glucose is available, adenylate cyclase is inactive, cAMP is not synthesized, and CRP does not activate transcription. Therefore, the behavior of a ∆crp mutant should approximate that of wild-type (WT) V. cholerae in a glucose-rich environment. When glucose is scarce as is the case in Luria-Bertani (LB) broth, adenylate cyclase is active. The resulting elevated levels of intracellular cAMP lead to formation of a cAMP-CRP complex, which activates transcription of hundreds of genes including those encoding transporters and enzymes required to utilize alternative sugars and amino acids as carbon sources (33–35). V. cholerae CRP also represses many processes required for virulence including cholera toxin, the toxin co-regulated pilus, and biofilm formation (33–38). In general, genes in the CRP regulon change their transcription in response to the availability of glucose as a carbon source.
The V. cholerae transcription factor HapR regulates transcription at high cell density. At low cell density, hapR mRNA is destabilized by the four small RNA’s qrr1-4 (39). Therefore, ∆hapR mutant behavior should approximate that of WT V. cholerae at low cell density. As the cell density increases, qrr’s are repressed in response to four secreted V. cholerae-derived small molecules known as autoinducers, and hapR mRNA is stable and transcribed (40). CRP and HapR interact at many levels to regulate gene transcription. For instance, hapR transcription is activated by CRP (34, 41). In addition, CRP and HapR share many binding sites, and joint binding alters transcription of target genes (42).
We investigated the observation that V. cholerae biofilm formation is inhibited in the presence of nitrate. We find that nitrite accumulates extracellularly when V. cholerae is cultured aerobically in LB broth supplemented with nitrate. The accumulated nitrite, rather than nitrate, directly inhibits V. cholerae surface accumulation. Interestingly, CRP is essential for nitrite production in shaking but not in static cultures, and this is manifested as resistance of the ∆crp mutant to inhibition of biofilm formation by environmental nitrate. We provide evidence that nitrite generation is inhibited by environmental oxygen and that the ∆crp mutant does not sufficiently deplete oxygen in aerated cultures to initiate nitrate reduction. We further report that a ∆hapR mutant is not susceptible to inhibition of V. cholerae biofilm accumulation by nitrite, but this is rescued by deletion of crp. While RNA-seq analysis identified many genes and putative small RNAs that respond to nitrate in WT V. cholerae but not the ∆hapR mutant, none had a previously described role in biofilm formation or dispersal. This suggests that nitrite inhibits V. cholerae biofilm accumulation in glucose-poor environments only at high cell density by an as-yet unelucidated transcriptional or post-transcriptional mechanism.
The V. cholerae machinery for oxygen and nitrate respiration is expressed under microaerophilic conditions, and nitrate is abundant in the ileal lumen (23). Therefore, we hypothesized that inhibition of V. cholerae biofilm accumulation by the nitrite resulting from nitrate reduction might be responsible for the minimal contribution of the VPS-dependent biofilm to V. cholerae colonization of the infant mouse small intestine (43, 44). Paracoccus aminovorans has been identified in the microbiome of cholera-afflicted individuals and increases V. cholerae surface attachment in both laboratory conditions and the infant mouse (7, 8). Here, we show that P. aminovorans respires the nitrite generated by V. cholerae in co-culture, thus rescuing V. cholerae biofilm accumulation. We propose that, upon arrival in the nitrate-rich terminal ileum, the contribution of VPS-dependent biofilm accumulation to V. cholerae intestinal colonization is determined by glucose availability via CRP, cell density via HapR, and the microbial environment via nitrite-reducing microbes. Thus, differences in nitrate and nitrite utilization by the intestinal microbial communities of neonatal and adult mammalian models as well as the human population may contribute to the observed variability in susceptibility to cholera.
RESULTS
Nitrate inhibition of V. cholerae biofilm accumulation in shaking cultures is dependent on the transcription factor CRP
During ongoing studies of CRP, a subtle, qualitative difference was observed between the biofilms formed by WT V. cholerae and a Δcrp mutant in shaking but not in static cultures. We hypothesized that this could be the result of differential availability of oxygen as a terminal electron acceptor in shaking WT V. cholerae and the ∆crp mutant cultures. To test this, we formed WT V. cholerae and Δcrp mutant biofilms in shaking LB cultures either alone or supplemented with 5 mM sodium nitrate or sodium fumarate, both of which can serve as alternative terminal electron acceptors. As shown in Figure 1A and B, nitrate significantly decreased biofilm accumulation by WT V. cholerae but not the ∆crp mutant. In contrast, fumarate did not alter WT V. cholerae biofilm accumulation, while it decreased biofilm accumulation by the Δcrp mutant. Nitrate and fumarate did not alter total growth (Fig. 1B). This demonstrates that the impact of nitrate on V. cholerae biofilm formation is not simply a reflection of its ability to serve as an alternative electron acceptor. Because nitrate is present in the terminal ileum and, therefore, could be important for V. cholerae intestinal colonization, we elected to further investigate inhibition of WT V. cholerae biofilm formation by nitrate and the role played by CRP in this process.
*Inhibition of biofilm accumulation by nitrate is dependent on HapR in both aerated and static cultures, and dependent on CRP in aerated cultures only. (A) Representative biofilm images and (B) quantification of biofilm formation and total growth by WT V. cholerae and a ∆crp mutant after approximately 18 h of incubation at 27°C in LB alone (–) or LB supplemented with 5 mM sodium nitrate (NO3−) or 5 mM sodium fumarate (C4H2O4−2). Significance for total growth measurements was calculated using an ordinary one-way ANOVA with Dunnett’s multiple comparisons test. Significance for biofilm accumulation was calculated using a Brown-Forsythe and Welch’s ANOVA with Dunnett’s T3 multiple comparisons test. (C) Representative biofilm images and (D) quantification of biofilm formation and total growth by WT V. cholerae and the indicated mutants cultured under static (stat) or aerated (shake) conditions in LB alone (–) or supplemented with 5 mM NaNO3 (+). An expanded view of the WT biofilm quantification is shown in Figure S1A. (E) Representative biofilm images and (F) quantification of biofilm formation and total growth by WT V. cholerae and the indicated mutants. Strains were rescued with the empty vector pFLAG-CTC (pCTL) or the same vector encoding CRP (pcrp-V5) as noted. Strains carrying plasmids were cultured with added ampicillin (100 µg/mL) and 0.5 mM IPTG. Significance for biofilm accumulation was calculated using a Welch’s t-test. Significance for total growth measurements was calculated using a Student’s t-test. In all biofilm experiments, the mean of six biological replicates is shown, and error bars reflect the standard deviation. ****P < 0.0001; ***P < 0.001; **P < 0.01; P < 0.05; ns, not significant.
The impact of nitrate on V. cholerae growth and surface association has principally been studied under hypoxic conditions (30, 45). To better understand the effect of nitrate on V. cholerae biofilm accumulation when both oxygen and nitrate are available as terminal electron acceptors, we quantified static and shaking biofilms formed by WT V. cholerae, a Δcrp mutant, and a ∆vpsA mutant in the presence and absence of nitrate. Nitrate supplementation caused a significant reduction in WT V. cholerae biofilm accumulation under both static and aerated conditions but did not alter total growth (Fig. 1C and D; Fig. S1A). In contrast, Δcrp mutant biofilm accumulation was unaffected by nitrate in shaking cultures but greatly reduced in static ones. Nitrate supplementation also reduced total growth of the Δcrp mutant in static cultures. However, the effect of nitrate on biofilm accumulation by the Δcrp mutant was approximately two orders of magnitude greater than that on its growth. Therefore, the reduction in total growth does not account for the decrease in Δcrp mutant biofilm accumulation. As expected, the ΔvpsA mutant accumulated very little biofilm under any of the conditions tested, and no difference in total growth was observed.
To establish that the inhibition of V. cholerae biofilm accumulation by nitrate was dependent on CRP in aerated cultures, we rescued the ∆crp mutant with a wild-type copy of crp provided on a plasmid behind an isopropyl-β-D-thiogalactopyranoside (IPTG)-inducible promoter. Inhibition of biofilm accumulation by nitrate in shaking cultures was restored to the ∆crp mutant by provision of crp in trans (Fig. 1E and F). In all strains carrying the plasmid, small but significant decreases in total growth were observed only in static but not in aerated cultures supplemented with nitrate, but this again did not account for the larger decreases in biofilm accumulation (Fig. 1F). This supports the conclusion that CRP is essential for inhibition of biofilm accumulation by nitrate in shaking cultures. Because CRP is active only in glucose-poor environments, we propose that the available carbon source plays a role in inhibition of V. cholerae biofilm accumulation by nitrate in aerated cultures.
A ∆hapR mutant is resistant to inhibition of biofilm accumulation by nitrate in both static and shaking cultures
CRP activates expression of HapR, the master regulator of high cell density quorum sensing, to repress biofilm formation (41, 46, 47). Furthermore, HapR is essential for biofilm dispersal at high cell density (48). To determine whether HapR plays a role in inhibition of biofilm accumulation by nitrate, we tested the impact of nitrate on biofilm accumulation by a ∆hapR mutant. ΔhapR mutant biofilm accumulation as well as total growth was unaffected by nitrate regardless of aeration (Fig. 1C and D). Because a ΔhapR mutant is locked in the low cell density state, this result suggests that V. cholerae biofilm accumulation is resistant to nitrate at low cell density.
Nitrate conversion to nitrite is essential for inhibition of WT and ∆crp mutant V. cholerae biofilm accumulation by nitrate
Because V. cholerae cannot reduce nitrate beyond nitrite, nitrite accumulates extracellularly when V. cholerae uses nitrate as an alternative electron acceptor. We reasoned, therefore, that inhibition of V. cholerae biofilm accumulation in the presence of nitrate could be a direct response to nitrate or a response to its metabolite nitrite. To test this, we formed WT, ∆crp, ∆vpsA, and ∆hapR mutant biofilms in LB supplemented with either 5 mM nitrate or nitrite under static and shaking conditions. As shown in Figure 2A and B, although aeration improved the growth of all strains to some extent, WT V. cholerae biofilm accumulation in the presence of nitrate and nitrite was negligible independent of aeration, while the ∆hapR mutant formed robust biofilms. In contrast, while nitrate inhibited ∆crp mutant biofilm accumulation only in static cultures, nitrite inhibited ∆crp mutant biofilm accumulation regardless of aeration. Because the phenotypes of the ∆crp and ∆hapR mutants were distinct, we conclude that CRP and HapR act at least in part independently to modulate the V. cholerae response to nitrate.
*V. cholerae reduction of nitrate to nitrite is required for inhibition of biofilm accumulation and occurs in aerobic cultures only when CRP is present. (A) Representative images of biofilms formed by WT V. cholerae and the indicated mutants after approximately 20 h of incubation at 27°C in LB supplemented with 5 mM nitrate (+NO3-) or 5 mM nitrite (+ NO2-). Biofilms were either formed statically (stat) or with aeration (shake). (B) Quantification of total growth and biofilm formation by the indicated strains cultured under the conditions indicated. The mean of six biological replicates is shown. Error bars reflect the standard deviation. For biofilm accumulation, significance was calculated using a Welch’s t-test. For total growth, significance was calculated using a Student’s t-test. (C) Quantification of nitrite (NO2-) generation (left graph) and growth as assessed by OD600 (right graph) over time for the indicated strains cultured in LB supplemented with nitrate. Measurements were made under both static and shaking conditions. The mean of biological triplicates is shown. Error bars reflect the standard deviation. For both nitrite and growth measurements, significant differences at each time point were assessed using an ordinary one-way ANOVA with Dunnett’s multiple comparisons test. (D) qRT-PCR quantification of expression of the narQ and nap genes required for nitrite generation from nitrate in the indicated strains. Cultures were aerated and harvested at 8 h. The mean of biological triplicates is shown. Error bars reflect the standard deviation. Significance was calculated using an ordinary one-way ANOVA with Dunnett’s multiple comparisons test. ****P < 0.0001; ***P < 0.001; **P < 0.01; P < 0.05; ns, not significant.
Based on our findings, we hypothesized that nitrite is the direct inhibitor of V. cholerae biofilm accumulation at high cell density and that CRP is essential for efficient reduction of nitrate to nitrite in aerated cultures. To test this, we monitored growth and nitrite generation in static and aerated cultures of WT V. cholerae as well as the Δcrp and ΔhapR mutants in LB supplemented with nitrate. In aerated cultures of WT V. cholerae, nitrite generation from nitrate was delayed by 2–3 h as compared with static cultures even though aeration improved growth (Fig. S1B). This suggests that oxygen availability inhibits nitrate reduction.
In static cultures, nitrite generation was similar for all strains tested despite significant differences in growth (Fig. 2C). In contrast, in aerated conditions, the Δcrp mutant culture generated a negligible amount of nitrite over the course of the experiment, while the WT and ΔhapR mutant strains had converted almost all the nitrate to nitrite after 5 h. While the Δcrp mutant grew more slowly than WT V. cholerae, this difference was too small to account for its inability to generate nitrite in aerated cultures. We hypothesize that oxygen inhibits nitrate generation by V. cholerae and that a Δcrp mutant, which does not initiate nitrate reduction to nitrite when cultured aerobically, may not be able to deplete aerated cultures of oxygen.
To further establish the role of nitrite in V. cholerae biofilm inhibition, we studied a ∆napC mutant that cannot generate nitrite. We first compared nitrite generation by WT V. cholerae and a ΔnapC mutant in LB supplemented with nitrate. Nitrite concentrations in 6- and 24-h ΔnapC mutant cultures were not significantly different from that of sterile nitrate-supplemented media (Fig. S1C). We then tested the ability of the ΔnapC mutant to form a biofilm in the presence of nitrate (Fig. S1D and E). While nitrate supplementation inhibited biofilm accumulation by WT V. cholerae, it had no effect on the ΔnapC mutant biofilm and no effect on total growth by either strain. We conclude that nitrate inhibits WT V. cholerae biofilm accumulation through conversion to nitrite.
Differential transcription of the genes encoding NarQ and NAP complex components does not underlie the absence of nitrite generation by the ∆crp mutant in shaking cultures
Our results showed that CRP is essential for nitrite generation in aerated cultures. Because CRP is a transcription factor, we reasoned it might play a role in oxygen-responsive transcriptional activation of the napA, napB, and napC genes as well as the gene encoding the nitrate-responsive histidine kinase NarQ (49). Therefore, in an aerated Δcrp mutant culture, transcription of these genes would not increase in response to nitrate as they would in aerated WT and ΔhapR mutant cultures. However, we found that the nap genes and narQ were similarly regulated regardless of genetic background and aeration (Fig. 2D; Fig. S2). We conclude that CRP does not activate transcription of the narQ and nap genes in shaking cultures supplemented with nitrate and that another function of CRP must enhance nitrite generation in aerated cultures.
Evidence that conversion of nitrite to nitric oxide is not responsible for inhibition of biofilm accumulation by nitrite
Nitric oxide is known to cause dispersal of the V. cholerae biofilm (50, 51). However, the V. cholerae genome does not encode the enzymes that convert nitrite to nitric oxide, and spontaneous reduction of nitrite to nitric oxide seems unlikely in the oxidizing conditions of aerobic cultures (51). Nevertheless, to test this possibility, we added the nitric oxide scavenger 2-carboxyphenyl-4,4,5,5-tetramethylimidazoline-1-oxyl-3-oxide (c-PTIO) to V. cholerae cultured statically in LB supplemented with nitrite (51). As shown in Figure S3, this treatment did not increase biofilm accumulation in nitrite-supplemented cultures.
CRP is essential for nitrate reduction by a ∆hapR mutant in aerated cultures and for the resistance of the ∆hapR mutant biofilm to inhibition by nitrate and nitrite
To further probe the resistance of the ∆hapR mutant biofilm to inhibition by nitrate and nitrite, we created a ΔhapRΔcrp double mutant and tested its ability to convert nitrate to nitrite and to form biofilms in the presence of nitrate under static and aerated conditions. Similar to the ∆crp mutant, the double mutant was unable to convert of nitrate to nitrite in aerated cultures (Fig. 2C). In static cultures, biofilm accumulation by the ΔhapRΔcrp double mutant in the presence of nitrate and nitrite was negligible, and total growth was similar to that of the Δcrp mutant (Fig. 3A). Therefore, deletion of crp in the ΔhapR mutant background restored the susceptibility of the ∆hapR mutant biofilm to inhibition by nitrate and nitrite. While these findings do not elucidate the mechanism underlying the resistance of the ΔhapR mutant to nitrate and nitrite, they do allow us to predict the contribution of V. cholerae VPS-dependent biofilm formation to surface colonization under conditions that might be encountered in the mammalian intestine provided the following two assumptions are true. (i) In a glucose-rich environment, CRP is inactive, and WT V. cholerae responds to nitrate as a ∆crp mutant would. (ii) At low cell density, HapR is inactive, and WT V. cholerae responds to nitrate as a ∆hapR mutant would. If this is correct, the following model shown in Figure 3B should be valid. In glucose-rich media, V. cholerae reduction of nitrate to nitrite should occur in static but not in aerated cultures. In glucose-rich environments, this leads to inhibition of biofilm formation by nitrate in static but not in aerated cultures independent of cell density (Fig. 3B [pathways Ia and b, and Ila and b]). In environments such as the terminal ileum, where glucose is limiting, V. cholerae reduces nitrate to nitrite regardless of aeration, but only high cell density cultures are susceptible to inhibition by nitrite (Fig. 3B [pathways Ic and llc]).
*CRP and HapR govern V. cholerae biofilm inhibition by nitrate. (A) Quantification of biofilm accumulation and total growth by WT V. cholerae as well as ∆crp, ∆hapR, and ∆hapR∆crp mutants cultured statically in LB alone or supplemented with 5 mM nitrate (NO3-) or nitrite (NO2-). The mean of biological triplicates is shown for all experiments. Error bars reflect the standard deviation. For biofilm accumulation, significance was calculated using a Brown-Forsythe and Welch’s ANOVA with a Dunnett’s T3 multiple comparisons test. For total growth, significance was calculated using an ordinary one-way ANOVA with a Dunnett’s multiple comparisons test. ****P < 0.0001; ***P < 0.001; **P < 0.01; P < 0.05; ns, not significant. (B) A model showing the impact of carbon source, cell density, and aeration on inhibition of V. cholerae biofilm formation by nitrate. This model is based on our observations using the Δcrp mutant, which mimics V. cholerae behavior in a glucose-rich environment (↑glucose), and the ΔhapR mutant, which mimics cells in a low cell density state (↓cell density). (Ia and lla) In a ∆crp mutant background or when glucose is plentiful (Δcrp [↑glucose]) and cultures are aerated, nitrate (NO3-) conversion to nitrite (NO2-) is blocked in cells with WT hapR (↑cell density) and ∆hapR (↓cell density), and biofilm accumulation proceeds. (lb and IIb) In a ∆crp mutant or when glucose is plentiful (Δcrp [↑glucose]) and cells are cultured statically, nitrate conversion to nitrite proceeds in both cells with WT hapR (↑high cell density) and ∆hapR (↓density), and biofilm accumulation is inhibited. (Ic and IIc) When glucose is scarce (WT crp [↓glucose]), nitrate is converted to nitrite in both cels with WT hapR (↑cell density) and ∆hapR (↓cell density). The ∆hapR mutant (↓cell density) biofilm is resistant to nitrite, while the WT hapR biofilm (↑cell density) is inhibited by nitrite. These pathways are highlighted in red because they are the only ones in which nitrate regulation of biofilms differs at high and low cell density. (Created in BioRender. Watnick, P. [2025], https://BioRender.com/n69b437.)
Nitrate has a global impact on the V. cholerae transcriptome
We reasoned that genes involved in inhibition of V. cholerae biofilm accumulation by nitrate should be differentially regulated in response to nitrate in WT V. cholerae and the ∆crp and ∆hapR∆crp mutants but not in the ∆hapR mutant. To gain insight into the genetic basis of V. cholerae biofilm inhibition by nitrate, we carried out an RNA-seq experiment including WT V. cholerae and ∆crp, ∆hapR, and ∆hapR∆crp mutants cultured statically for 8 h in LB alone or supplemented with 5 mM nitrate (Tables S1 to S4). In general, more RNAs were differentially regulated in response to nitrate in the ∆hapR mutant than in the other genetic backgrounds tested. The ∆crp mutant had the fewest differentially regulated RNAs, and the ∆hapR∆crp mutant had an intermediate number (Fig. 4A). This suggests that CRP plays an important role in the transcriptional and post-transcriptional response of V. cholerae to nitrate, while HapR mutes the transcriptional response to nitrate. Only 74 coding sequences, 22 intergenic RNAs, and 14 antisense RNAs were commonly differentially regulated in the four strains in response to nitrate (Fig. 4B). Gene ontology (GO) analysis of coding sequences using the Panther Overrepresentation test showed significant overrepresentation of nitrate-regulated genes involved in aerobic respiration and iron transport but not in surface adhesion (Fig. 4C; Tables S1to S4) (52, 53). Genes involved in iron transport were overrepresented in all genetic backgrounds. One possible explanation is that the ferric uptake regulator Fur, which represses transcription of iron uptake genes in response to ferrous iron availability, is less active in the presence of nitrate (Fig. 4C) (54). We can conclude that the differential regulation of iron transport genes in response to nitrate is not responsible for the distinct responses of the ∆crp and ∆hapR mutant to nitrate.
Nitrate regulation of aerobic respiration is CRP dependent, while nitrate control of iron-regulated genes is CRP and HapR independent. (A) Table showing numbers of differentially regulated coding sequences (CDS), intergenic region RNAs (IGR), and antisense RNAs within coding sequences (AS) for the indicated strains when cultured statically for 8 h either alone (–) or in the presence of 5 mM nitrate (+). (B) Venn diagrams illustrating overlap in transcripts differentially regulated by nitrate in each of the genetic backgrounds studied. Transcripts found in coding sequences (CDS), intergenic regions (IGR), and antisense RNA (AS CDS) are illustrated separately. Genes that are differentially regulated in all but the ∆hapR mutant are circled in red. (C) Gene ontology analysis of differentially regulated genes using the Panther Overrepresentation test.
Surprisingly, nitrate supplementation activated not only transcription of the genes required for nitrate respiration but also those required for respiration of oxygen. However, the latter were only differentially regulated by nitrate when CRP was present (Fig. 4C). Indeed, transcription of bd-l and cbb3, the genes encoding the principal aerobic terminal oxidases of V. cholerae, was greatly reduced in the ∆crp mutant in both the presence and absence of nitrate (Fig. S4) (23). We have previously shown that oxygen inhibits nitrate reduction. Based on these findings, we hypothesize that, due to a paucity of aerobic terminal oxidases, the ∆crp mutant cannot efficiently deplete aerated cultures of oxygen, and this may underlie the inability of the ∆crp mutant to generate nitrate.
Nitrate does not decrease V. cholerae biofilm accumulation via known pathways
With the exception of Bap1 (VC1888), the genes responsible for forming the V. cholerae biofilm matrix are found in two islands, *vps-*I, which includes VC0916-27, and *vps-*II, which includes VC0934-9 (3, 6, 43, 55, 56). These genes are regulated by two transcription factors, VpsR (VC0665) and VpsT (VCA0952), both of which are activated by binding of the second messenger cyclic-diguanylate (c-di-GMP) (57–61). We first scrutinized transcription of these genes (Fig. 5A). Differential regulation of most of these genes did not meet our criteria for fold-change and statistical significance. Genes that were significantly regulated did not follow the pattern expected based on biofilm phenotype. For instance, while nitrate decreased WT V. cholerae biofilm accumulation, transcription of biofilm genes was increased in response to nitrate. Conversely, while nitrate had no impact on the ∆hapR mutant biofilm, transcription of biofilm genes was decreased in response to nitrate. To confirm these puzzling patterns of gene regulation, we undertook qRT-PCR analysis for a subset of these genes (Fig. 5B). Supporting our RNA-seq findings, nitrate activated the biofilm gene vpsA in WT V. cholerae and repressed the biofilm genes vpsR, vpsT, and vpsA in the ∆hapR mutant. These results suggest that nitrate does not inhibit V. cholerae biofilm accumulation by decreasing transcription of the biofilm synthesis genes.
*Repression of the vps genes is not responsible for inhibition of V. cholerae biofilm accumulation by nitrate. (A) Heat map illustrating differential regulation of biofilm formation and biofilm dispersal genes as measured by RNA-seq. Fold-change represents transcription in +nitrate/–nitrate conditions. White stars indicate genes that met the criteria for significance FC >2 and padj <0.05. (B) qRT-PCR confirmation of vps gene transcription in the presence (+) and absence (–) of nitrate. The indicated strains were cultured statically for 8 h. The mean of biological triplicates is shown. Error bars reflect the standard deviation. Significance was calculated using a Student’s t-test. **P < 0.01; P < 0.05; ns, not significant.
We then explored known post-transcriptional mechanisms of biofilm regulation. Two previously identified sRNAs, ryhB and qrrX, regulate V. cholerae biofilm formation. The Fur-regulated small RNA ryhB, whose transcription was increased with nitrate supplementation for all genetic backgrounds including the ∆hapR mutant, activates biofilm formation (62). Because ryhB activates biofilms, it is unlikely that ryhB is responsible for biofilm inhibition by nitrate (63). qrrX is an RNA sponge that destabilizes the qrr1-4 sRNAs, which maintain low cell density signaling and transcription of the vps genes (64, 65). qrrX transcription increased in the presence of nitrate in WT V. cholerae, which would be expected to decrease biofilm formation, but transcription of qrrX is quite low in ∆crp, ∆hapR, and ∆crp∆hapR mutants (Fig. S5). Therefore, qrrX also seems an unlikely candidate for biofilm inhibition. There were many additional coding and putative non-coding RNAs within this data set that have not previously been associated with biofilm inhibition and, therefore, warrant further exploration.
V. cholerae accumulates on surfaces at low cell density, and HapR contributes to biofilm dispersion at high cell density (48). Therefore, we entertained the alternative hypothesis that nitrate induces biofilm dispersal at high cell density rather than inhibiting biofilm matrix synthesis. Several studies have reported genes that are essential for V. cholerae biofilm dispersal (66–69). We reasoned that transcription of biofilm dispersal genes might be activated in response to nitrate in all genetic backgrounds tested except ΔhapR, which mimics a low cell density state. A total of 49 genes and non-coding RNAs were differentially regulated in all but the ∆hapR mutant background (Fig. 4B, red circle). However, this group did not include any genes known to be involved in biofilm formation or dispersal. Examining genes that were differentially regulated in the ∆crp and ∆hapR∆crp mutant backgrounds only and unchanged in WT V. cholerae and the ∆hapR mutant, we identified one gene VC0658 or cdgI previously identified to be involved in biofilm dispersal (Fig. 5A) (66). CdgI is a c-di-GMP phosphodiesterase and, therefore, would be predicted to decrease intracellular levels of c-di-GMP and thereby vps gene transcription. Because we did not observe a decrease in vps gene transcription in response to nitrate, it seems unlikely that CdgI is responsible for biofilm inhibition by nitrate unless it is acting via a novel, c-di-GMP-independent mechanism. We conclude that, if HapR regulates the biofilm response to nitrate by increasing biofilm dispersal, this is unlikely to occur through a previously described pathway.
Paracoccus aminovorans, a microbe associated with the microbiomes of patients with cholera, rescues V. cholerae biofilm accumulation in the presence of nitrate
In a human host, inhibition of V. cholerae VPS-dependent biofilm formation due to nitrate reduction to nitrite might diminish colonization of the ileum, where the concentration of nitrate is high. We reasoned that a nitrite-reducing microbial community might mitigate inhibition of VPS-dependent biofilm formation and, thus, predispose to V. cholerae colonization and disease. To test this, we co-cultured V. cholerae with the aerobic gram-negative rod Paracoccus aminovorans. Sequenced strains of P. aminovorans JCM 7685 and DSM 8537 (NCBI Bioprojects PRJEB8789 and PRJEB17339) encode the genes for reduction of nitrite to nitric oxide, nitrous oxide, and finally nitrogen gas. Furthermore, P. aminovorans is found in the microbiota of patients with active cholera but not uninfected household contacts (8). In a later publication, P. aminovorans was found to increase colonization of abiotic surfaces and the neonatal mouse intestine via formation of a VPS-dependent biofilm (7). Using the P. aminovorans strain previously isolated from cholera stool and generously provided by the Weil and Ng laboratories, we first cultured P. aminovorans statically in LB supplemented with 5 mM nitrate or 5 mM nitrite, and quantified nitrite remaining after 48 h. As shown in Figure 6A and B, nitrite was not present in the medium after 48 h of culture, suggesting that P. aminovorans is able to reduce nitrite. To determine whether P. aminovorans might reduce the nitrite generated from nitrate by V. cholerae, we statically cultured WT V. cholerae and the ∆crp mutant alone or with P. aminovorans in LB supplemented with nitrate, and measured the nitrite concentration after 48 h. In both co-cultures, P. aminovorans greatly diminished nitrite concentrations in the medium at the end of the incubation period (Fig. 6B). To investigate whether reduction of nitrite by P. aminovorans might rescue V. cholerae surface attachment, we compared V. cholerae surface adhesion in LB alone or supplemented with nitrate in the presence and absence of P. aminovorans. As shown in Figure 6C and D, the presence of P. aminovorans rescued biofilm accumulation by WT and ∆crp mutant V. cholerae in the presence of nitrate. Interestingly, P. aminovorans also significantly improved the growth of the ∆crp mutant, suggesting it secretes an as-yet unidentified metabolite that can be utilized by ∆crp mutant. P. aminovorans made a very little biofilm on its own and did not increase surface adhesion by a ∆vps mutant. This demonstrates that P. aminovorans rescues VPS-dependent biofilm formation in the presence of nitrate rather than providing V. cholerae an alternate means of surface adherence. To determine the contribution of V. cholerae and P. aminovorans to the biofilm mass measured, we formed biofilms with V. cholerae, which is streptomycin (Sm) resistant, and a Sm-sensitive P. aminovorans strain. After 48 h, biofilms were dispersed and colony forming units (CFUs) were enumerated on LB agar alone or supplemented with Sm. As shown in Figure S6, CFUs were comparable regardless of the addition of antibiotics, suggesting that V. cholerae is the major contributor to measured biofilm biomass in co-cultures. We conclude that members of the microbiota that reduce nitrite can rescue V. cholerae biofilm accumulation in the presence of nitrate.
*Paracoccus aminovorans consumes nitrite produced by V. cholerae and rescues V. cholerae biofilm accumulation in the presence of nitrate. (A) Quantification of nitrite in sterile LB broth supplemented with 5 mM nitrite (Std) and the spent supernatant of a P. aminovorans (Pa) cultured statically for 48 h in LB supplemented with 5 mM nitrite (Pa). (B) Quantification of nitrite in the supernatants of WT V. cholerae and the indicated mutants cultured statically for 48 h alone or with P. aminovorans (Pa) in LB supplemented with 5 mM nitrate. The mean of biological triplicates is shown. Error bars reflect the standard deviation. Significance was calculated using a Student’s t-test. (C) Representative images of biofilms formed by WT V. cholerae and the indicated mutants cultured statically for 48 h alone or with P. aminovorans (Pa) in LB (–) or LB supplemented with 5 mM NaNO3 (+). (D) Quantification of biofilm formation and total growth by the indicated strains cultured as described in (C). The mean of six biological replicates is shown. Error bars reflect the standard deviation. For biofilm accumulation, significance was calculated using a Welch’s t-test. For total growth, significance was calculated using a Student’s t-test. ****P < 0.0001; ***P < 0.001; **P < 0.01; P < 0.05; ns, not significant.
DISCUSSION
Here, we report that V. cholerae vigorously respires nitrate in shaking cultures to produce nitrite, that CRP, a transcription factor that is active in the absence of glucose, is essential for aerobic reduction of nitrate to nitrite, and that nitrite inhibits V. cholerae biofilm accumulation when HapR is active at high cell density but not in a ∆hapR mutant, which mimics low density behavior. Based on our findings, we propose that inhibition of V. cholerae biofilm accumulation by nitrate is controlled by carbon source and cell density. Finally, we show that co-culture with P. aminovorans, an inhabitant of the human intestinal microbiota that is associated with the development of cholera, respires nitrite and augments V. cholerae surface accumulation in the presence of nitrate. These findings are significant because they highlight the role of the microbiota in regulation of surface colonization by V. cholerae in environments such as the mammalian terminal ileum, where both oxygen and nitrate are available as terminal electron acceptors. They also leave unanswered questions that are discussed below.
We have not elucidated the mechanism by which CRP activates nitrate reduction in aerated cultures, although we have ruled out transcriptional activation of the narQ and nap genes as the basis for this. Our RNA-seq data suggest that expression of the principal terminal oxidases bd-I and cbb3 is decreased in a ∆crp mutant, leading to impaired oxygen respiration. Previous work suggests that aerobic bacterial cultures progress toward a hypoxic state as cell density increases. We predict that this would occur more slowly in a ∆crp mutant (70). Our data show that aeration delays nitrate reduction by wild-type V. cholerae. If this is a direct response to oxygen, nitrate reduction should be further delayed in a ∆crp mutant. Two conserved and well-studied transcription factors respond to oxygen availability and could contribute to the inability of the ∆crp mutant to reduce nitrate in aerated cultures. The first is the fumarate nitrate reduction regulator FNR, which is active as a dimer when its (4Fe-4S)^2+^ cluster cofactor is bound. In the presence of O_2_, FNR dimerization is inhibited by sulfur oxidation of this co-factor (71). Therefore, FNR is a direct sensor of oxygen availability. In a previous study, FNR was identified as essential for nitrate reduction by V. cholerae in anaerobic cultures, but transcriptional targets of FNR were not further explored (30). In other organisms, FNR regulates additional genes found to be essential for V. cholerae nitrate reduction including the moa genes, encoding the molybdenum cofactor biosynthesis proteins, and the nqr genes, encoding the NADH:ubiquinone oxidoreductase (72–74). The ArcAB two component system also controls gene transcription in response to oxygen availability (75). ArcB is a membrane-associated sensor that responds to anaerobiosis by activating the transcription factor ArcA via phosphorylation. ArcB is thought to respond to oxygen availability by sensing the oxidation state of quinones that shuttle electrons in aerobic electron transport chains (76). In V. cholerae, ArcA activates biofilm formation via vpsT, represses flagellar synthesis, and increases expression of the virulence factors cholera toxin and the toxin co-regulated pilus (77–79). Because this regulation occurs at the transcriptional level, it does not explain the observed response to nitrate. In other organisms, both FNR and ArcAB also control expression of small RNAs (sRNAs) that regulate protein expression (27, 80–82). In our RNA-seq experiment, many putative sRNAs were differentially transcribed in response to nitrate, and it is quite likely that one or more of these contributes to regulation of nitrate reduction at the post-transcriptional level.
We also have not uncovered the mechanism by which nitrite inhibits V. cholerae VPS-dependent biofilm formation nor the basis for the resistance of the ∆hapR mutant biofilm to nitrite. Our experiments rule out transcriptional regulation of the vps genes as a mechanism, and it seems unlikely that nitrite would directly poison the enzymes involved in WT and ∆crp biofilm formation but not that of the ∆hapR mutant*,* because biofilm formation by all depends on the same suite of enzymes. While HapR has previously been associated with activation of biofilm dispersal at high cell density, we have not identified a plausible role for known biofilm dissolution genes in HapR-dependent inhibition of V. cholerae biofilm accumulation by nitrite (48). Our RNA-seq experiments suggest that HapR regulates many putative sRNAs in response to nitrate. We hypothesize that one of these is responsible for post-transcriptional inhibition of biofilm formation in the presence of nitrite.
Finally, we have not explored the basis of increased transcription of iron scavenging genes in nitrate-supplemented V. cholerae cultures. These genes are regulated by the ferric uptake repressor Fur. Fur represses transcription of iron transport genes by binding reduced or ferrous iron (Fe^2+^) (83). Under the conditions of our RNA-seq experiment, while we expect most of the available iron to be in the ferric or oxidized state (Fe^3+^), there is sufficient Fe^2+^ to promote repression of transcription by Fur (84, 85). One possibility is that addition of nitrate decreases the availability of Fe^2+^, but this cannot be explained by differential regulation of V. cholerae vciB (VC0283), which is essential for iron reduction in the periplasm, as this gene was not differentially regulated in the presence of nitrate (Tables S1to S4) (86, 87). Furthermore, while NapC has been shown to participate in iron reduction, its contribution is relatively small (87, 88). Therefore, competition between Fe^3+^ and nitrate for reduction by NapC is not a likely mechanism. We have discussed possible roles for FNR and ArcAB in the response to nitrate. Highlighting the complexity of the bacterial response to electron acceptor availability, in other organisms, molybdenum cofactor biosynthesis and nitrate metabolism is under control of Fur, FNR, and ArcA, and ArcA regulates both Fur and uptake of ferrous iron (89–92). A methodical examination of this complex regulatory network will be required to understand activation of iron-regulated genes by nitrate.
Our studies highlight key unexplored aspects of V. cholerae surface association in nitrate-rich environments that are likely to have implications for disease. V. cholerae colonization of the ileum, a region of the intestine where oxygen is present and nitrate concentrations reach 6 mM, is critical for expression of V. cholerae virulence factors (25). Previous studies have shown that CRP is essential for robust V. cholerae colonization of the mammalian intestine (93). We propose that in the glucose-poor environment of the ileum, V. cholerae CRP is active, available oxygen is consumed, and nitrate is reduced (Fig. 7). Thus, V. cholerae colonizes the ileum in the presence of nitrite until high density is reached. HapR then activates a response to nitrite that inhibits or disperses the biofilm. Because HapR also represses expression of the toxin co-regulated pilus, cholera toxin, and the biofilm synthesis genes, we propose that HapR enacts a regulatory program that limits V. cholerae colonization of the intestine and prolongs survival of the host (94–96). Despite the actions of HapR, some hosts are differentially susceptible to cholera (9). We suggest that intestinal microbes such as P. aminovorans, which convert the nitrite generated by V. cholerae nitrate respiration to inactive byproducts, short circuit HapR-induced biofilm inhibition in response to nitrate.
A model for V. cholerae high cell density colonization of the ileum in the presence and absence of a nitrite-reducing microbe such as P. aminovorans. Schematic showing the impact of a nitrite-reducing organism such as P. aminovorans on V. cholerae intestinal colonization at high cell density. In a resistant host, V. cholerae generates nitrite (NO2-) from host nitrate (NO3-) in the terminal ileum in a process that is dependent on the transcription factor CRP. At high cell density, when HapR is active, nitrite inhibits VPS exopolysaccharide-dependent biofilm formation on the intestinal surface either by preventing accumulation or accelerating dispersal. This reduces the burden of V. cholerae in the intestine and increases the likelihood of pathogen clearance without symptomatic infection. In a susceptible host, the intestinal microbiome includes an organism such as P. aminovorans, which can further reduce nitrite to nitrogen gas (N2). Mobilization of nitrite allows VPS-dependent biofilm accumulation to proceed. This results in a greater burden of V. cholerae in the gut and an increased likelihood of symptomatic infection. (Created in BioRender. Santos, R. [2025], https://BioRender.com/p56l530.)
MATERIALS AND METHODS
Bacterial strains and media
The quorum sensing-competent V. cholerae strain C6706, which is Sm-resistant, served as the parental strain for all experiments. Sm-sensitive and Sm-resistant P. aminovorans strains were generously provided by Drs. Wai-Leung Ng and Ana Weil. The Sm-sensitive strain was previously isolated from stool swabs of a family member of a cholera patient in Bangladesh (7, 8). Both V. cholerae and P. aminovorans strains were cultured with shaking in LB broth (Difco, Miller formulation) supplemented with Sm (100 µg/mL, Gibco) unless otherwise noted. Escherichia coli strains used for cloning were cultured in LB broth supplemented with ampicillin (100 µg/mL, Sigma). For rescue experiments, 0.5 mM IPTG (MedChemExpress) was used to induce expression. V. cholerae was cultured at 27°C, while P. aminovorans and E. coli were cultured at 37°C. Except for the ∆napC mutant, all plasmids and strains used were previously published and are shown in Table S5. The ∆napC mutant was generated by double homologous recombination as previously described (33). The following primers were used to generate the deletion construct, which was cloned into pWM91 (97): primer 1: AATTGGGTACCGGGCCCCCCCCTGGCTGGCATTGCTCAT, primer 2: TCCTTATTGCCAGCCTTCTGGGCTCGATAGACGAAG, primer 3: GAAGGCTGGCAATAAGGA, primer 4: CTTATCGATACCGTCGACCTCGAGCAGGAAACCACCATGAG.
Biofilm assays
Cells were cultured overnight, diluted 1:100 into fresh LB broth, and cultured to mid-exponential phase using the conditions described above. Mid-exponential cells were diluted in the indicated medium to yield an OD_600_ of 0.05 as measured on a SpectraMax ABS microplate reader (Molecular Devices). For V. cholerae biofilm assays, 300 µL of this diluted culture was aliquoted into 10 × 75 mm borosilicate glass tubes (Fisher) and incubated for approximately 20 h at 27°C either statically or with agitation at 200 rpm as noted. Tubes were positioned vertically for static cultures and at an angle for aerobic cultures to increase exposed surface area. A similar procedure was followed for co-cultures with P. aminovorans with the following variations. Where noted, a 1:1 ratio of P. aminovorans to V. cholerae was diluted into a 300 µL volume in a 10 × 75 mm borosilicate tube and cultured statically at room temperature (RT) for 48 h. Planktonic cells were removed and replaced with an equal volume of phosphate-buffered saline as well as several 1 mm glass beads. Biofilm-associated cells were then dispersed by vortexing. The density of biofilm and planktonic cells was quantified by measuring an OD_600_. Total growth was calculated as the sum of the planktonic and biofilm OD_600_. A minimum of three biological replicates was performed, and each experiment was repeated at least once with similar results. Biofilms were imaged using a Nikon D3400 digital camera.
Nitrite measurements
Overnight cultures were diluted 1:100 in LB medium supplemented with 5 mM sodium nitrate or nitrite as noted, and cultured at 27°C for the indicated time. Prior to the measurements, cultures were centrifuged at 12,000 × g for 1 min at room temperature. The supernatants were collected and filtered through a 0.2 µm filter. A kit based on the Griess reagent (Biotium) was used to measure nitrite concentrations per the manufacturer’s instructions. Briefly, 50 µL of the Griess reagent was combined with 150 µL of the sample and 1.3 mL of ddH_2_O, and incubated at RT for at least 30 min prior to measurement of A_548_ using an SpectraMax iD5 microplate reader (Molecular Devices). The concentration of nitrite was determined by plotting the experimental values against a standard curve generated from a 1 mM stock of nitrite solution. Biological triplicates were performed, and each experiment was repeated at least once with similar results.
RNA isolation
Total RNA was isolated from 3 mL cultures cultivated at 27°C without agitation for 8 h. Bacterial pellets were collected and resuspended in 500 µL TriZol (Invitrogen), incubated at 60°C for 10 min, followed by purification using Direct-zol RNA Miniprep kit (Zymo R2052) following the manufacturer’s instructions. RNA for both RNA-seq analysis and qRT-PCR was prepared using this method.
Generation of RNA-Seq data
Illumina cDNA libraries were generated using a modified version of the RNAtag-seq protocol (98). Briefly, 0.5–1 μg of total RNA was fragmented, depleted of genomic DNA, dephosphorylated, and ligated to DNA adapters carrying 5′-AN8-3′ barcodes of known sequence with a 5′ phosphate and a 3′ blocking group. Barcoded RNAs were pooled and depleted of rRNA using the riboPOOLS rRNA depletion kit (siTOOLs Biotech). Pools of barcoded RNAs were converted to Illumina cDNA libraries in two main steps: (i) reverse transcription of the RNA using a primer designed to the constant region of the barcoded adaptor with addition of an adapter to the 3′ end of the cDNA by template switching using SMARTScribe (Clontech), and (ii) PCR amplification using primers whose 5′ ends target the constant regions of the 3′ or 5′ adaptors and whose 3′ ends contain the full Illumina P5 or P7 sequences. cDNA libraries were sequenced on the Illumina NovaSeq 6000 platform to generate paired-end reads.
RNA-seq data analysis
Sequencing reads from each sample in a pool were demultiplexed based on their associated barcode sequence using custom scripts. Up to one mismatch in the barcode was allowed, provided it did not make assignment of the read to a different barcode possible. Barcode sequences were removed from the first read as were terminal G’s from the second read that may have been added by SMARTScribe during template switching. Reads were aligned to Vibrio cholerae N16961 genome, and read counts were assigned to genes and other genomic features using custom scripts. Differential expression analysis was conducted with DESeq2 (99) and EdgeR (100). Genes were considered significantly differentially regulated if a twofold or greater change in expression was measured and if the adjusted P value was less than or equal to 0.05. RNA-seq data were visualized using the IGV (v2.10.3) developed at the Broad Institute. Genome ontology analysis utilized Panther version 19.0 (52, 101). Significantly overrepresented biological functions were identified using a Fisher exact test for significance and P < 0.05.
qRT-PCR
Total RNA (500 ng) served as a template to make c-DNA using the iScript reverse transcriptase supermix (Bio-Rad). A QuantStudio 5 Real-Time PCR machine was used to measure amplification of cDNA added to an iTaq Universal SYBR Green Supermix reaction (Bio-Rad). rpoA was used as a reference gene for static cultures, while clpX was used for aerated cultures. Technical duplicates and biological triplicates were performed.
Data analysis
Data were graphed and analyzed for statistical significance using Prism (Graphpad). A mean and standard deviation were calculated. For qRT-PCR, nitrite generation, and total growth measurements, a Student’s t-test or one-way analysis of variance (ANOVA) was used to evaluate significance. For biofilm accumulation measurements, a Welch’s t-test or Brown-Forsythe and Welch’s ANOVA was used to evaluate significance.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Reidl J, Klose KE. 2002. Vibrio cholerae and cholera: out of the water and into the host. FEMS Microbiol Rev 26:125–139. doi:10.1111/j.1574-6976.2002.tb 00605.x 12069878 · doi ↗ · pubmed ↗
- 2Walton MG, Cubillejo I, Nag D, Withey JH. 2023. Advances in cholera research: from molecular biology to public health initiatives. Front Microbiol 14:1178538. doi:10.3389/fmicb.2023.117853837283925 PMC 10239892 · doi ↗ · pubmed ↗
- 3Absalon C, Van Dellen K, Watnick PI. 2011. A communal bacterial adhesin anchors biofilm and bystander cells to surfaces. Plos Pathog 7:e 1002210. doi:10.1371/journal.ppat.100221021901100 PMC 3161981 · doi ↗ · pubmed ↗
- 4Berk V, Fong JCN, Dempsey GT, Develioglu ON, Zhuang X, Liphardt J, Yildiz FH, Chu S. 2012. Molecular architecture and assembly principles of Vibrio cholerae biofilms. Science 337:236–239. doi:10.1126/science.122298122798614 PMC 3513368 · doi ↗ · pubmed ↗
- 5Huang X, Nero T, Weerasekera R, Matej KH, Hinbest A, Jiang Z, Lee RF, Wu L, Chak C, Nijjer J, Gibaldi I, Yang H, Gamble N, Ng WL, Malaker SA, Sumigray K, Olson R, Yan J. 2023. Vibrio cholerae biofilms use modular adhesins with glycan-targeting and nonspecific surface binding domains for colonization. Nat Commun 14:2104. doi:10.1038/s 41467-023-37660-037055389 PMC 10102183 · doi ↗ · pubmed ↗
- 6Yildiz FH, Schoolnik GK. 1999. Vibrio cholerae O 1 El Tor: identification of a gene cluster required for the rugose colony type, exopolysaccharide production, chlorine resistance, and biofilm formation. Proc Natl Acad Sci USA 96:4028–4033. doi:10.1073/pnas.96.7.402810097157 PMC 22414 · doi ↗ · pubmed ↗
- 7Barrasso K, Chac D, Debela MD, Geigel C, Steenhaut A, Rivera Seda A, Dunmire CN, Harris JB, Larocque RC, Midani FS, Qadri F, Yan J, Weil AA, Ng WL. 2022. Impact of a human gut microbe on Vibrio cholerae host colonization through biofilm enhancement. Elife 11:e 73010. doi:10.7554/e Life.7301035343438 PMC 8993218 · doi ↗ · pubmed ↗
- 8Midani FS, Weil AA, Chowdhury F, Begum YA, Khan AI, Debela MD, Durand HK, Reese AT, Nimmagadda SN, Silverman JD, Ellis CN, Ryan ET, Calderwood SB, Harris JB, Qadri F, David LA, La Rocque RC. 2018. Human gut microbiota predicts susceptibility to Vibrio cholerae infection. J Infect Dis 218:645–653. doi:10.1093/infdis/jiy 19229659916 PMC 6047457 · doi ↗ · pubmed ↗