Investigation of the Cyanothece nitrogenase cluster in Synechocystis: a blueprint for engineering nitrogen-fixing photoautotrophs
Deng Liu, Anindita Bandyopadhyay, Michelle Liberton, Himadri B. Pakrasi, Maitrayee Bhattacharyya-Pakrasi

TL;DR
This study identifies key genes from Cyanothece that enable nitrogen fixation in photosynthetic organisms, offering a blueprint for engineering green, solar-powered nitrogen-fixing systems.
Contribution
The study defines a minimal gene set from Cyanothece's nitrogenase cluster essential for nitrogen fixation in phototrophic systems.
Findings
The structural genes nifHDK, nifBSUENPVZTXW, and hesAB are crucial for optimal nitrogenase function in phototrophs.
Optimal gene expression is essential for efficient nitrogen fixation in engineered systems.
A refactored gene cluster in Synechocystis demonstrates the feasibility of synthetic nitrogen-fixing phototrophs.
Abstract
The nitrogenase gene cluster of unicellular diazotrophic cyanobacteria, such as Cyanothece, is frequently selected by nature for nitrogen-fixing partnerships with eukaryotic phototrophs. The essential cluster components that confer an advantage in such partnerships remain underexplored. To use this cluster for the development of synthetic, phototrophic nitrogen-fixing systems, a thorough and systematic analysis of its constituent genes is necessary. An initial effort to assess the possibility of engineering this cluster into non-diazotrophic phototrophs led to the generation of a Synechocystis 6803 strain with significant nitrogenase activity. In the current study, a refactoring approach was taken to determine the dispensability of the non-structural genes in the cluster and define a minimal gene set for constructing a functional nitrogenase for phototrophs. Using a bottom-up strategy,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
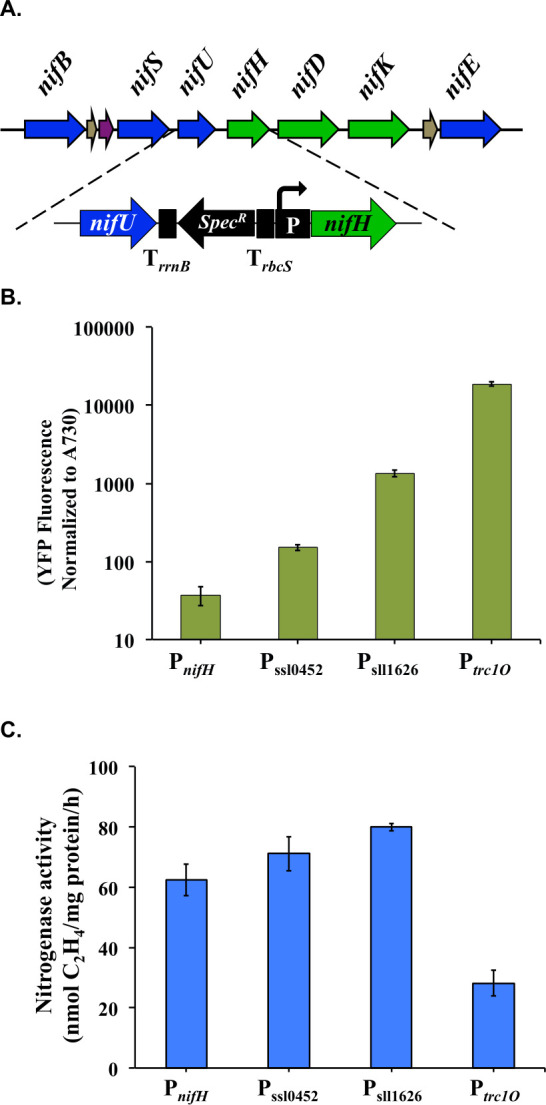 Fig 1
Fig 1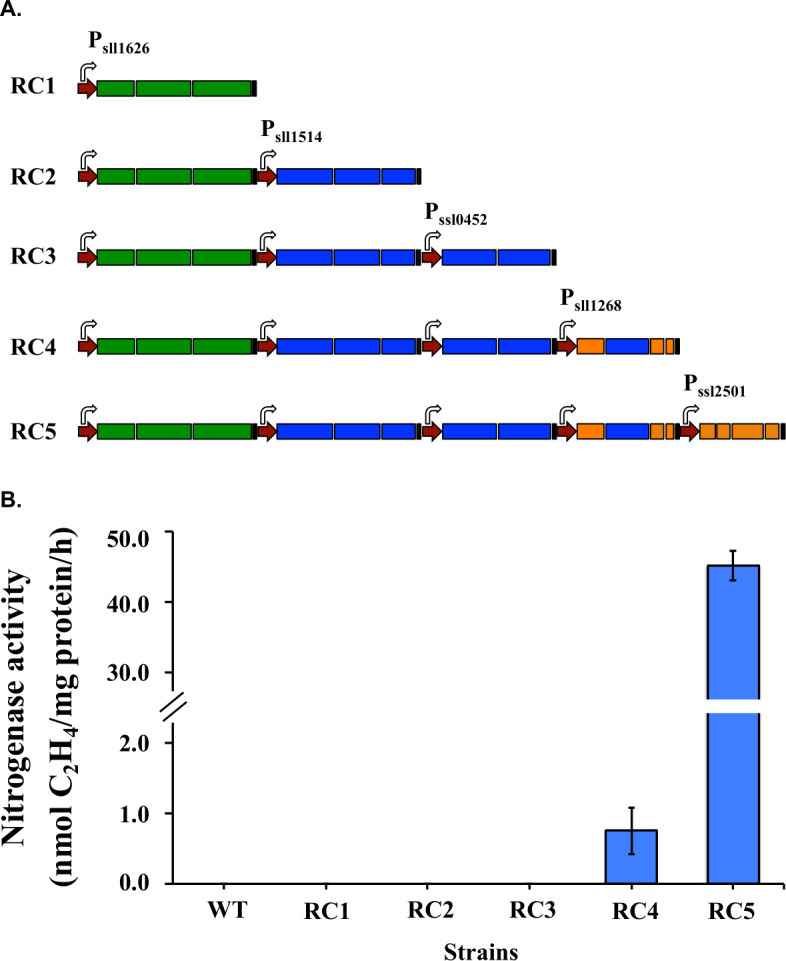 Fig 2
Fig 2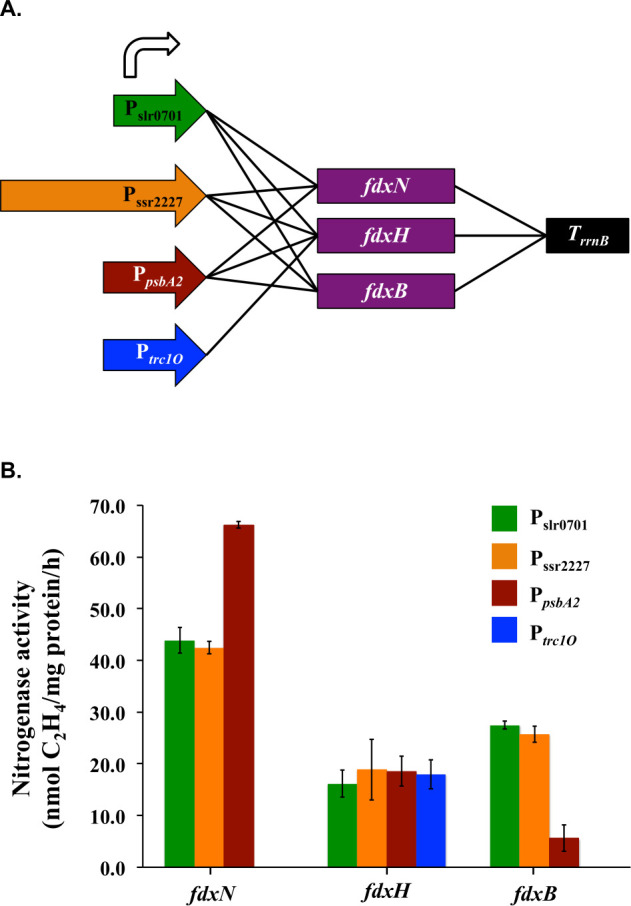 Fig 3
Fig 3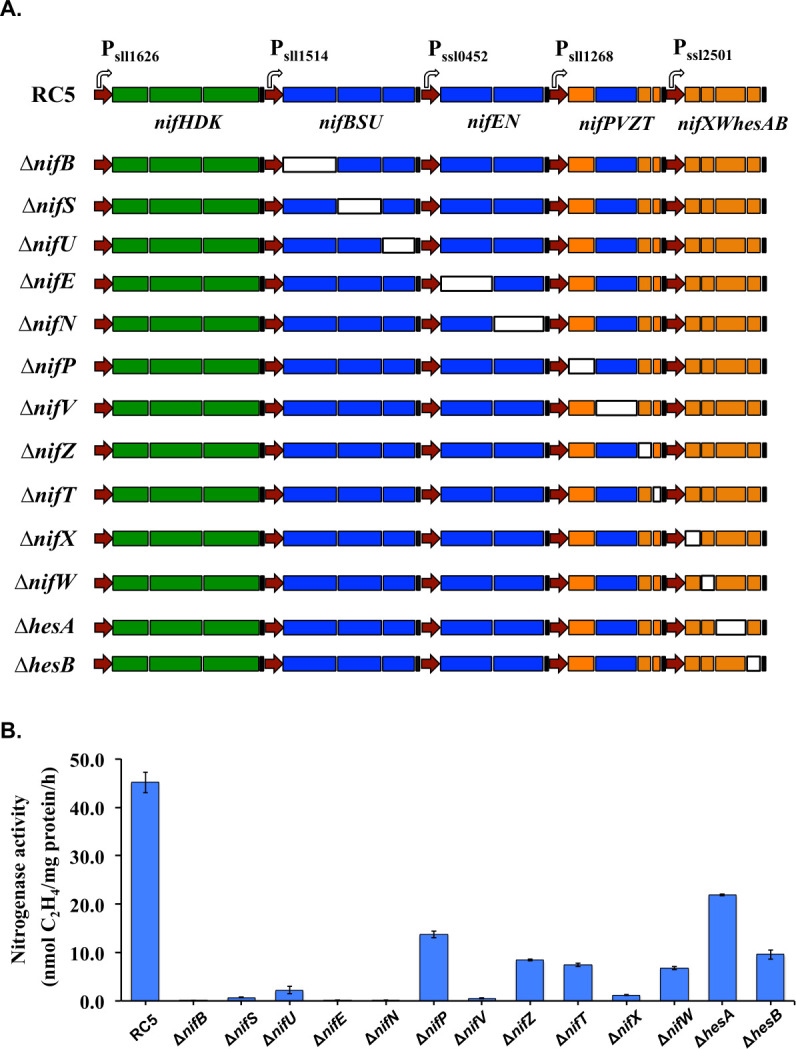 Fig 4
Fig 4Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMetalloenzymes and iron-sulfur proteins · Photosynthetic Processes and Mechanisms · Algal biology and biofuel production
INTRODUCTION
Multiple examples of natural endosymbiotic events suggest a propensity for the nitrogenase (nif) gene cluster of unicellular cyanobacteria of the order Chroococcales to be selected for photosynthetic–diazotrophic partnerships. Of these, the Cyanothece nif cluster has been referred to as a ‘privileged’ partner, forming associations with diatoms, such as Rhopalodia, Epithemia, and Climacodium, as well as the coccolithophore Braarudosphaera bigelowii (1–5). These naturally occurring instances suggest that the Cyanothece nif cluster harbors features that are conducive for nitrogenase to function in a photoautotroph and demands a thorough investigation as we intensify our efforts toward developing synthetic nitrogen- and carbon-fixing systems. In a first-of-its kind effort in this direction, the unicellular non-diazotrophic cyanobacterium Synechocystis sp. PCC 6803 (hereafter Synechocystis 6803) was engineered to fix nitrogen by introduction of the nitrogenase gene cluster from Cyanothece sp. ATCC 51142 (6). The resulting strain exhibited significant rates of nitrogen fixation, affirming the potential of this cluster to generate functional nitrogen-fixing machinery in photoautotrophs and initiating our next round of efforts into its more in-depth analysis.
The commonly occurring nitrogenase enzyme is composed of two metalloproteins: the MoFe protein (dinitrogenase, NifDK) and the Fe protein (dinitrogenase reductase, NifH) (7). Besides these structural subunits of the enzyme, three iron–sulfur clusters (F, P, and M clusters) are required to form a mature nitrogenase enzyme complex (8, 9). The processes of assembly and insertion of these three iron–sulfur clusters into the nitrogenase enzyme are complicated, involving multiple steps (10). The genetic elements required for the nitrogenase enzyme to mature vary in different diazotrophic species (11, 12). Previous efforts to engineer Escherichia coli with nitrogen fixation experimented with various combinations of nif genes as a strategy to optimize the activity. The study identified nine genes from the nif cluster of Paenibacillus sp. WLY78, including nifHDK and nifBENXhesAnifV, as essential (13). A different study determined 10 genes (anfHDGK and nifJUSVFB) from the nif clusters of Azotobacter vinelandii and Klebsiella oxytoca to be essential for nitrogenase activity (14). On the contrary, a recent report by Yang et al. identified 13 genes, including nifHDK and nifENBYUSVFMJ, as the essential components of the nitrogenase cluster of Klebsiella oxytoca (15). Nitrogenase activity could be further improved 10-fold using a refactoring approach and expressing 18 nif genes in various combinations, along with additional genes, and by altering the expression ratios of genes (16).
In our earlier work, we demonstrated the feasibility of expressing the native 24 gene nif cluster from Cyanothece 51142 in Synechocystis 6803. The additional seven genes in the cluster were not included in this study (6). Furthermore, except for the structural genes nifHDK, the essentiality of the other genes in the cluster in nitrogen fixation was not determined in this photosynthetic chassis. In the current study, we reconstructed nif gene operons using a bottom–up strategy in Synechocystis 6803. Promoters, ribosome binding sites (RBS), and transcriptional terminators previously identified (17) were used to control the expression of the nif genes. We found that 16 genes organized into five engineered operons formed a functional nitrogenase enzyme complex with remarkable activity. More importantly, using the CRISPR/cpf1 strategy (18), we strategically removed each gene, except nifHDK, from the reconstructed nif operons to determine their essentiality. Our study showed that deletion of any of these genes leads to a decrease in nitrogenase activity, thus establishing the minimal gene set required for optimal nitrogenase function in a non-diazotrophic photoautotroph.
RESULTS
Expression levels of structural genes affect nitrogenase activity
In the nif cluster of Cyanothece 51142, the transcriptional start sites and operons are not well defined. The only well-characterized operon in this cluster is the nifHDK operon, the expression of which is driven by the sequence before the nifH gene (19). In order to test the effect of the expression strength of these genes on nitrogenase activity, we cloned the sequence between nifU and nifH into cassettes containing different promoters (Fig. 1A).
Expression levels of nitrogenase structural genes affect nitrogen fixation activities in engineered Synechocystis 6803. (A) Schematic map of changes on the 5′-UTR sequence before the nifH gene. Three promoters were tested, Pssl0452, Psll1626, and Ptrc1O. (B) The strengths of the tested promoters were compared to the native promoter PnifH. The strengths were tested from the expression levels of the reporter protein EYFP. (C) Nitrogenase activity under different strength promoters preceding the nifH gene. Samples were collected from cultures under 12 h light/12 h dark conditions in BG110 medium. Nitrogen fixation activity was assayed by acetylene reduction, and error bars represent the standard deviations observed from at least three independent experiments.
Three promoters characterized in an earlier study (17), P_sll0452_, P_sll1626_, and Ptrc1O, were tested for their efficacy in driving gene expression in comparison to the natural promoter PnifH. Using EYFP as the reporter protein (Fig. S1), we found that the strength of PnifH was the lowest among all the promoters tested (Fig. 1B). Nitrogenase activities could be enhanced when the stronger promoters P_sll0452_ and P_sll1626_ were used. However, when the strongest promoter Ptrc1O was used to drive the expression of nifHDK, more than 50% reduction in nitrogenase activity was observed (Fig. 1C). These results suggested that optimal expression of the structural nif genes is important for achieving high nitrogenase activity, a finding in accordance with a study on Klebsiella (20).
Bottom–up strategy to re-organize nitrogen fixation genes
To find the essential genes for nitrogenase activity, we used a bottom–up strategy to build operons containing the genes from the native Cyanothece 51142 nif cluster (Fig. 2A). Promoters, RBS, and transcriptional terminators used in this study are native genetic elements of Synechocystis 6803 that were characterized earlier (17). The re-organized operons were inserted into the backbone of the shuttle vector pCB-SC101 (17).
Rebuilding the functional nitrogenase enzyme in Synechocystis 6803. (A) A schematic map of reconstruction of nif genes using a bottom–up strategy. Five operons were added sequentially into the plasmid pCB-SC101, which were transferred into Synechocystis 6803 as the CK strain. For each operon, promoters, RBS, and terminators were organized with ORFs to form an independent expression cassette. Shown are promoters (red), transcription terminators (black), and the genes for the three structural proteins, nifHDK (green), necessary cofactors (blue), accessory proteins (orange). (B) Nitrogenase activities of engineered strains containing sets of reconstructed nif operons. Samples were collected from cultures under 12 h light/12 h dark conditions in BG110 medium. Nitrogen fixation activity was assayed by acetylene reduction, and error bars represent the standard deviations observed from at least three independent experiments.
Of the 24 genes in the native Cyanothece nif cluster, 16 genes have been annotated as nif genes in the JGI/IMG microbial database (genome.jgi.doe.gov) (21). The others are mostly genes of unknown function, except one that is annotated as a ferredoxin gene. The 16 annotated genes were sequentially organized into five independent operons, nifHDK, nifBSU, nifEN, nifPVZT, and nifXWhesAB (Fig. S2). Five engineered strains (RC1 to RC5) were generated by transforming Synechocystis 6803 with plasmids containing the above operons in different combinations. The strain RC4 containing 12 genes (nifHDKBSUENPVZT) showed significant activity, generating 0.75 nmol ethylene per mg total protein in 1 h(Fig. 2B). Complementing these 12 genes with the nifXWhesAB operon generated the RC5 strain, which exhibited a 60-fold increase in activity, thereby confirming these genes as essential components of the nitrogen-fixing machinery. This observation was in accordance with our earlier findings (6), and the RC5 strain was selected for further analyses of the cluster.
Influences of ferredoxins on nitrogenase activity
As direct electron donors to Fe protein, ferredoxins are known to significantly affect nitrogenase activity (22, 23). There is one ferredoxin annotated as fdxN in the 24-gene native nif cluster of Cyanothece 51142 and two more ferredoxins, fdxH and fdxB, in the 35-gene extended cluster. To evaluate the role of ferredoxins in nitrogen fixation, the three ferredoxin genes, fdxN, fdxH, and fdxB, were introduced into the RC5 strain. Four promoters, P_slr0701_, P_ssr2227_, PpsbA2, and Ptrc1O were used to independently drive the expression of each ferredoxin gene (Fig. 3A). One interesting observation was that, when driven by the Ptrc1O promoter, only the fdxH gene could be transformed into Synechocystis. Therefore*,* for genes fdxN and fdxB, only three promoters were tested.
Ferredoxins affect nitrogenase activity in engineered Synechocystis 6803. (A) Scheme showing the combination of promoters and ferredoxin genes. The expression cassettes were integrated into the plasmid pRSF1010. (B) Nitrogenase activities are affected by type and expression level of ferredoxins in engineered Synechocystis 6803. Samples were collected from cultures under 12 h light/12 h dark conditions in BG110 medium. Nitrogen fixation activity was assayed by acetylene reduction, and error bars represent the standard deviations observed from at least three independent experiments.
The ferredoxins tested had a diverse effect on nitrogenase activity (Fig. 3B). When the fdxN gene under the PpsbA2 promoter was introduced into RC5, a 50% increase in nitrogenase activity was observed. In contrast, when fdxB under a strong promoter was introduced into RC5, a significant reduction in activity was observed. Interestingly, the introduction of fdxH into RC5 leads to a 60% decrease in activity, irrespective of the promoter used. This implied that the fdxH gene product negatively impacts nitrogen fixation.
Essential nif genes for nitrogenase activity in Synechocystis 6803
Of the 16 annotated genes in strain RC5, only the structural genes nifHDK have been functionally characterized. The role of the other 13 genes in nitrogen fixation remains largely unknown. To determine the essential set of genes for nitrogenase activity in Synechocystis 6803, we used the CRISPR/cpf1 strategy (18) to seamlessly delete each of the 13 genes after the nifHDK operon (Fig. S3). The 13 mutant strains based on RC5 were generated as shown in Fig. 4A.
Deletion analysis of the genes shown in the re-constructed nif operons. (A) Scheme showing the deletion of specific genes in the operons using the CRISPR/cpf1 system. The hollow rectangles represent the deleted genes, and the colored rectangles represent the remaining genes. (B) Nitrogenase activity of the deletion strains in engineered Synechocystis 6803. Samples were collected from cultures under 12 h light/12 h dark conditions in a BG110 medium. Nitrogen fixation activity was assayed by acetylene reduction, and error bars represent the standard deviations observed from at least three independent experiments.
As anticipated, deletion of nifB, nifE, or nifN leads to a complete cessation of nitrogenase activity (Fig. 4B). This observation is in agreement with the fact that nifENB genes are required for M cluster biosynthesis, and they are the most conserved genes besides nifHDK among all diazotrophs (11, 24). Similarly, the deletion of any of the three genes nifSU and nifV resulted in over 100-fold decrease in nitrogenase activity. NifSU proteins are required for synthesis of iron–sulfur clusters (25), while nifV encodes the enzyme homocitrate synthase. Homocitrate is a component of the M cluster present in the catalytic center of dinitrogenase (26). NifP encodes a serine acetyltransferase, which has been identified as a key enzyme in cysteine biosynthesis in E. coli (27). Deletion of nifP (cysE2) resulted in approximately 60% reduction in nitrogenase activity in Synechocystis, implying its essential role in the process. Genes nifZTW encode proteins that are suggested to be involved in MoFe protein maturation, but none were found to be essential for nitrogen fixation in E. coli (13–15). However, our results showed that deletion of nifZ leads to 70% decrease in nitrogenase activity, while deletions of nifT and W lead to about 80% decrease, implying their essential roles in nitrogen fixation in a photoautotroph. NifX is an interesting protein that has been identified as having various roles in different diazotrophs. It is a negative regulator of nitrogenase activity in Klebsiella (28), while in Azotobacter, it is required for the biosynthesis of the M cluster (29). Deletion of nifX in Synechocystis 6803 resulted in an over 100-fold decrease in nitrogenase activity, implying its essential role. Genes hesAB are not widely found in diazotrophs, and their functions are yet to be determined. Our results suggested that they play an important role in nitrogenase activity in Synechocystis 6803, especially hesB, a finding corroborated by studies in the diazotrophic cyanobacterium Anabaena sp. PCC7120 (30).
DISCUSSION
Members of the genus Cyanothece that engage in aerobic nitrogen fixation harbor one of the largest nif clusters known among cyanobacteria (31). The fact that these unicellular cyanobacteria are preferred by higher photoautotrophs as their nitrogen-fixing partners suggests that their nif cluster might confer certain advantages. This hypothesis is further strengthened by recent studies that revealed that close relatives of Cyanothece with highly similar nif clusters are now functioning as early-stage nitrogen-fixing organelles, called diazoplasts or nitroplasts (1, 4, 32). Therefore, efforts to engineer photoautotrophic systems involving eukaryotic algae, lichens, or plants for the production of nitrogen-rich compounds will benefit from a detailed analysis of the Cyanothece nif cluster. In an earlier study, we investigated the efficacy of the Cyanothece 51142 nif cluster in imparting nitrogen-fixing ability to non-diazotrophic photoautotrophs by introducing it into Synechocystis 6803. The engineered Synechocystis strain exhibited significant rates of nitrogen fixation, laying the foundation for the current study, which further investigates the functioning of the cluster in photoautotrophs. We focused on identifying a set of essential genes and the optimal expression ratios of some of the key genes involved in the process of nitrogen fixation as the next step toward designing an efficient synthetic cluster for nitrogenase. We successfully re-organized the genes of this cluster to form independent operons in Synechocystis 6803 and demonstrated significant nitrogenase activity. Although the highest nitrogenase activity was observed in strain RC5, strain RC4 harboring only 12 nif genes also fixed nitrogen at low levels. In a study that introduced the nif gene cluster from Leptolyngbya boryana into Synechocystis 6803, nitrogenase activity around 4 nmol ethylene per mg cell dry weight per hour was demonstrated (33). We determined that under the testing conditions, the ratio of total protein per cell dry weight is 0.25 (data not shown), which translates into an activity of about 1 nmol ethylene per mg total protein in 1 h, a rate very close to that observed in the RC4 strain. Although our engineered RC5 strain exhibits significantly lower activities (45 nmol per mg total protein in 1 h) compared to Cyanothece 51142 (1262 nmol per mg total protein in 1 h), the activities are significantly higher compared to the Synechocystis 6803 strain expressing genes from Leptolyngbya boryana (1 nmol per mg total protein in 1 h) (33). Thus, the nitrogen fixation rates observed in the Synechocystis strain expressing genes from Cyanothece 51142 are very encouraging and validate our hypothesis that the Cyanothece nif cluster offers an optimal starting point for designing synthetic solar-powered nitrogen fixing systems.
In order to convert N_2_ into ammonia, reducing power is required by the nitrogenase enzyme complex. Ferredoxins have been identified as one of the primary electron donors to the connected Fe protein of nitrogenase (34). However, in strain RC5, none of the ferredoxins from Cyanothece 51142 were transferred into Synechocystis 6803, which indicates that the endogenous ferredoxins in Synechocystis 6803 recognize the introduced nitrogenase Fe protein and function as the electron bridge for dinitrogen gas reduction. It has not been determined which ferredoxin gene is essential for electron transfer to nitrogenase in Cyanothece 51142; therefore, all three annotated genes in the extended cluster were tested in the RC5 strain. Our results indicated that only the fdxN gene positively impacted nitrogenase activity when expressed at a higher level, suggesting that FdxN could be involved in electron transfer to nitrogenase. Studies have revealed that pathways of electron transfer to nitrogenase in metabolically diverse strains differ, and the oxygen dynamics of the cell is an important factor that determines the ferredoxins involved in the process (34). In our study, FdxH and FdxB negatively impacted nitrogenase activity, implying that these ferredoxins could be involved in diverting electrons from nitrogen fixation to other processes in Synechocystis 6803, where the cellular oxygen dynamics are very different from Cyanothece 51142. It has been shown that electron transport chains from plant organelles function to support nitrogenase activity in engineered E. coli (23). The fact that the RC5 strain fixes nitrogen without any ferredoxins from Cyanothece suggests that transferring nif genes without the ferredoxins from the native cluster might endow higher phototrophs with nitrogenase activity, minimizing the number of genes to be transferred.
In this work, we have described the minimal set of nif genes essential for nitrogenase activity in a nondiazotrophic cyanobacterium. However, the complete cellular machinery needed for robust and efficient nitrogen fixation under aerobic conditions, a key piece of the puzzle for engineering photoautotrophs, remains largely unknown. More recently, a transposon sequencing (Tn-Seq) strategy has been used in various non-diazotrophic species, including cyanobacteria, to identify essential gene sets under special conditions (35, 36). Extending such studies to diazotrophic cyanobacterial strains can reveal a more complete picture of the entire cellular machinery needed for high levels of nitrogen fixation.
MATERIALS AND METHODS
Microorganisms, culture conditions, and media
All cyanobacterial strains were cultured in 100 mL flasks with fresh BG11 medium and appropriate antibiotics (20 µg/mL kanamycin, 3 µg/mL gentamycin, or 20 µg/mL spectinomycin). Cells were pre-cultured at 30°C under 50 µmol photons⋅m^−2^⋅s^−1^ constant light, with shaking at 150 rpm. For nitrogenase assays, pre-cultured cells were collected and washed with fresh BG11 medium without nitrate (BG11_0_) and resuspended in 500 mL fresh BG11_0_ medium. Cells were grown at 30°C with air bubbling under 12 h light/dark conditions under 50 µmol photons⋅m^−2^⋅s^−1^ of light. All cloning was performed in E. coli strain XL1-Blue grown in LB medium in culture tubes or on agar plates at 37°C and supplemented with 50 µg/mL kanamycin, 20 µg/mL gentamycin, or 30 µg/mL spectinomycin, as needed.
Construction of recombinant plasmids and engineered strains
Plasmids and strains used in this study are listed in Table S1, and all primers are listed in Table S2. Plasmids were constructed by the Gibson isothermal DNA assembly method (37) using linear fragments purified from PCR products. Genomic DNA from the strain Cyanothece 51142 and Synechocystis 6803 were used as templates for PCR. The genetic elements, including promoters, ribosome binding sites, and transcriptional terminators, were chosen from our previous study (17). The plasmid pUC118 (38) was used as the backbone to construct the plasmids used in the promoter assay for structural genes. The broad host plasmid pRSF1010 (39) was used as the backbone for the promoter—EYFP assays—and for the evaluation of the ferredoxin coding genes. The shuttle vector described in our previous study, pCB-SC101, was used for the re-construction work using the bottom–up strategy (17). The plasmid pSL2680 constructed in our lab (18) was used for the knock-out experiments.
Plasmids based on pUC118 and pCB-SC101 vectors were all transformed by natural transformation (40) into the Synechocystis 6803 TSyNif-2 and wild-type strains, respectively. A tri-parental conjugation method was used to transfer all pRSF1010 derivative plasmids into Synechocystis 6803 cells (41) using a helper strain of E. coli containing the pRL443 and pRL623 plasmids (42). Transformants were selected on BG11 agar plates containing 20 µg/mL kanamycin, 3 µg/mL gentamycin, or 20 µg/mL spectinomycin, as needed. Isolated Synechocystis 6803 transformants were checked by PCR to confirm the presence of the desired constructs.
All PCR amplifications were performed using Phusion high-fidelity DNA polymerase (Thermo Scientific). Plasmids and PCR products were purified using the GeneJET (Thermo Scientific) plasmid miniprep and gel extraction kits, respectively. Oligonucleotides were designed using SnapGene software (GSL Biotech LLC) and synthesized by IDT (Coralville, IA). All of the plasmids constructed in this study were checked by sequencing to confirm the correct DNA coding information (Genewiz, NJ).
Fluorescence measurements
Fluorescence intensities were measured as described earlier (17). Briefly, all cultures were adjusted to similar cell densities, with an OD_730 nm_ of 0.2 (about 1 × 10^8^ cells/mL) at the start of the experiment. Three independent replicates of each culture were then transferred to six-well plates and grown for 3 days, followed by fluorescence measurements. The fluorescence intensity and the optical density of each culture were determined on a BioTek Synergy Mx plate reader (BioTek, Winooski, VT). The excitation and emission wavelengths were set to 485 and 528 nm for EYFP, respectively. All fluorescence data were normalized by culture density.
Measurement of nitrogen fixation activity
Nitrogen fixation activity was measured by an acetylene reduction assay modified from a previously published method (43). The assay was performed as follows: 25 mL of cyanobacterial culture grown in BG11_0_ medium was transferred to a 125 mL air-tight glass vial; 10 µM DCMU was added to the culture; vials were flushed with pure argon; and cultures were incubated in 12 h light/dark conditions. Cells in the sealed vials were cultured overnight, and, at the time point D1, 5 mL acetylene was added into the sealed vials, followed by 3 h of incubation in light at 30°C. 200 µL of gas was sampled from the headspace and injected into an Agilent 6890N gas chromatograph equipped with a Poropak N column and a flame ionization detector using argon as the carrier gas. The temperature of the detector, injector, and oven were 200, 150, and 100°C, respectively.
Determination of cellular total proteins and cell dry weight
Total protein levels were determined on a plate reader (Bio-Tek Instruments, Winooski, VT) using a bicinchoninic acid assay kit (Pierce, Rockford, IL) according to the manufacturer’s instructions. Cell dry weight was determined from cell pellets of 100 mL culture aliquots that were centrifuged for 15 min at 4°C and 9,000 × g, washed twice with distilled water, and dried at 85°C until the weight was constant.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Moulin SLY, Frail S, Braukmann T, Doenier J, Steele-Ogus M, Marks JC, Mills MM, Yeh E. 2024. The endosymbiont of Epithemia clementina is specialized for nitrogen fixation within a photosynthetic eukaryote. ISME Commun 4:ycae 055. doi:10.1093/ismeco/ycae 05538707843 PMC 11070190 · doi ↗ · pubmed ↗
- 2Prechtl J, Kneip C, Lockhart P, Wenderoth K, Maier U-G. 2004. Intracellular spheroid bodies of Rhopalodia gibba have nitrogen-fixing apparatus of cyanobacterial origin. Mol Biol Evol 21:1477–1481. doi:10.1093/molbev/msh 08614963089 · doi ↗ · pubmed ↗
- 3Carpenter EJ, Janson S. 2000. Intracellular cyanobacterial symbionts in the marine diatom Climacodium Frauenfeldianum (Bacillariophyceae). J Phycol 36:540–544. doi:10.1046/j.1529-8817.2000.99163.x 29544002 · doi ↗ · pubmed ↗
- 4Coale TH, Loconte V, Turk-Kubo KA, Vanslembrouck B, Mak WKE, Cheung S, Ekman A, Chen J-H, Hagino K, Takano Y, Nishimura T, Adachi M, Le Gros M, Larabell C, Zehr JP. 2024. Nitrogen-fixing organelle in a marine alga. Science 384:217–222. doi:10.1126/science.adk 107538603509 · doi ↗ · pubmed ↗
- 5Hagino K, Onuma R, Kawachi M, Horiguchi T. 2013. Discovery of an endosymbiotic nitrogen-fixing cyanobacterium UCYN-A in Braarudosphaera bigelowii (Prymnesiophyceae). P Lo S ONE 8:e 81749. doi:10.1371/journal.pone.008174924324722 PMC 3852252 · doi ↗ · pubmed ↗
- 6Liu D, Liberton M, Yu J, Pakrasi HB, Bhattacharyya-Pakrasi M. 2018. Engineering nitrogen fixation activity in an oxygenic phototroph. M Bio 9:e 01029-18. doi:10.1128/m Bio.01029-1829871920 PMC 5989072 · doi ↗ · pubmed ↗
- 7Tezcan FA, Kaiser JT, Mustafi D, Walton MY, Howard JB, Rees DC. 2005. Nitrogenase complexes: multiple docking sites for a nucleotide switch protein. Science 309:1377–1380. doi:10.1126/science.111565316123301 · doi ↗ · pubmed ↗
- 8Hu Y, Ribbe MW. 2013. Nitrogenase assembly. Biochim Biophys Acta 1827:1112–1122. doi:10.1016/j.bbabio.2012.12.00123232096 PMC 3622157 · doi ↗ · pubmed ↗