Representational learning by optimization of neural manifolds in an olfactory memory network
Bo Hu, Nesibe Z. Temiz, Chi-Ning Chou, Peter Rupprecht, Claire Meissner-Bernard, Benjamin Titze, SueYeon Chung, Rainer W. Friedrich

TL;DR
This study explores how the brain learns to distinguish odors by analyzing the geometry of neural representations in zebrafish.
Contribution
The paper introduces manifold capacity as a novel analytical framework to link neural geometry with odor discrimination behavior.
Findings
Olfactory training increases separation of neural manifolds for task-relevant odors.
Manifold capacity better predicts odor discrimination than other population activity descriptors.
pDp stores information in the geometry of representational manifolds, supporting sensory and semantic maps.
Abstract
Cognitive brain functions rely on experience-dependent internal representations of relevant information. Such representations are organized by attractor dynamics or other mechanisms that constrain population activity onto “neural manifolds”. Quantitative analyses of representational manifolds are complicated by their potentially complex geometry, particularly in the absence of attractor states. Here we trained juvenile and adult zebrafish in an odor discrimination task and measured neuronal population activity to analyze representations of behaviorally relevant odors in telencephalic area pDp, the homolog of piriform cortex. No obvious signatures of attractor dynamics were detected. However, olfactory discrimination training selectively enhanced the separation of neural manifolds representing task-relevant odors from other representations, consistent with predictions of autoassociative…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Extended Data Fig. 1 |
Extended Data Fig. 1 |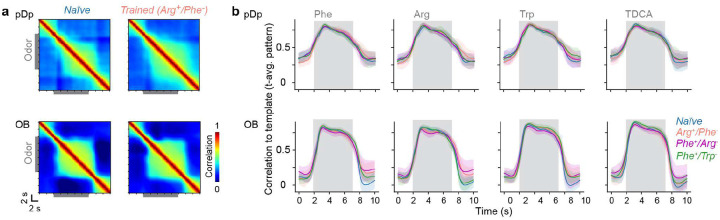 Extended Data Fig. 2 |
Extended Data Fig. 2 |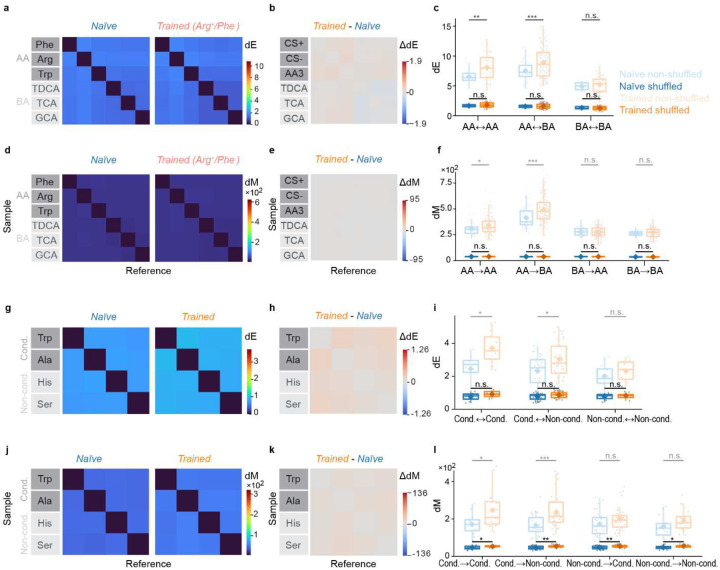 Extended Data Fig. 3 |
Extended Data Fig. 3 |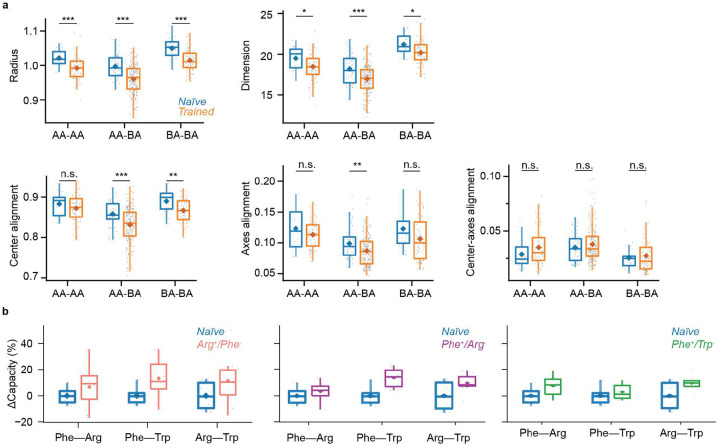 Extended Data Fig. 4 |
Extended Data Fig. 4 |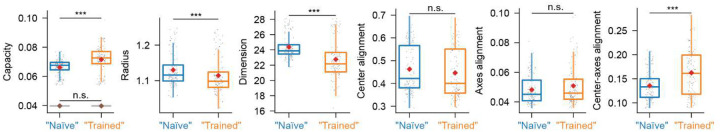 Extended Data Fig. 5 |
Extended Data Fig. 5 |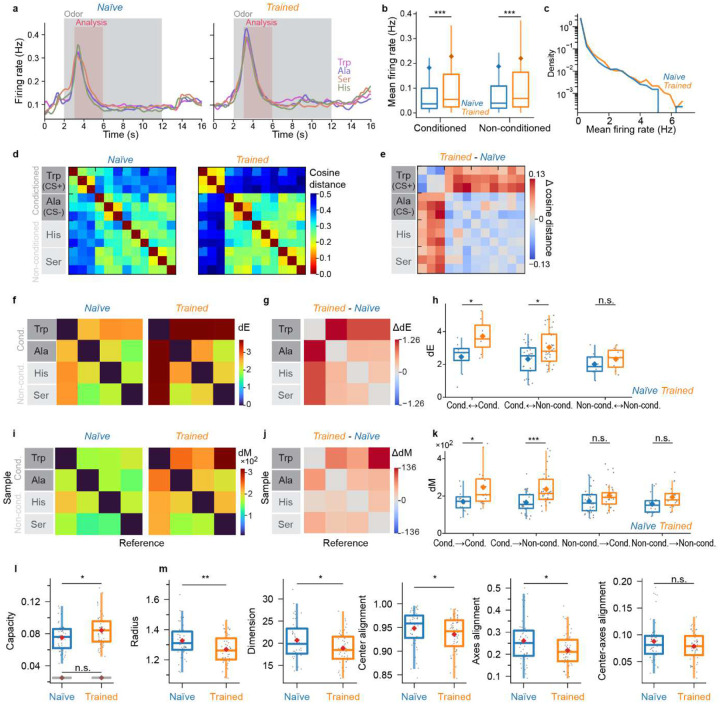 Extended Data Fig. 6 |
Extended Data Fig. 6 |Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNeurobiology and Insect Physiology Research · Neural dynamics and brain function · Zebrafish Biomedical Research Applications
INTRODUCTION
Learning generates organized representations of relevant information in the brain that generalize to novel inputs and serve as a basis for cognition. Representational learning is thought to depend on autoassociative memory networks that store information by activity-dependent modifications of recurrent synaptic connectivity. Classical models predict that learning enhances recurrent excitation among specific ensembles of neurons, resulting in convergent attractor dynamics that mediates pattern classification^1–5^. Recurrent networks can also give rise to a continuum of stable attractor states that may, for example, function as a cognitive map^2,6^. Alternatively, representational learning may be mediated by mechanisms that do not depend on attractor dynamics. Generally, networks may represent relevant information by mapping patterns of input activity to specific subspaces of neuronal state space according to semantic relationships between inputs^7,8^. Hence, learning may cause geometrical state space modifications that define continuous neural manifolds without establishing distinct attractor states^7–15^ but the possible functions of such manifolds in autoassociative memory remain to be explored.
Autoassociative memory has been proposed to be a primary function of piriform cortex, a paleocortical brain area that receives non-topographic sensory input from the olfactory bulb. Within piriform cortex, excitatory neurons are recurrently connected through an extensive “association fiber system” that is plastic and under neuromodulatory control^16–19^. These observations gave rise to the hypothesis that learning results in the formation of neuronal assemblies in piriform cortex that represent odor objects and mediate pattern classification by convergent dynamics^20^. However, recent analyses of neuronal activity in piriform cortex did not reveal obvious signatures of attractor states. For example, odor-evoked activity is not persistent but phasic and curtailed by inhibition^21–23^. Moreover, while convergent dynamics are expected to reduce variability, intra- and inter-trial variability of neuronal activity in piriform cortex appears high in comparison to inputs from the olfactory bulb^23–26^. Nonetheless, passive odor exposure and active learning modify odor-evoked population activity in piriform cortex depending on stimulus statistics or task structure^25,27–31^. For example, activity patterns evoked by different odors become more highly correlated after presentation of odors in a binary mixture^25^, or after association of multiple odors with a common reward^29^. Hence, experience may reorganize odor representations in piriform cortex in a task-relevant fashion in the absence of discrete attractor dynamics.
Autoassociative memory has recently been explored in computational models of recurrent networks that were constrained by data from telencephalic area pDp of adult zebrafish^32^, the teleost homolog of the piriform cortex^33,34^. When information was stored in these models by enhancing connectivity among small assemblies of excitatory and inhibitory neurons, a transition from discrete attractor dynamics to continuous representational manifolds occurred when networks entered a regime of precise synaptic balance^32^. This regime is characterized by strong and correlated excitatory and inhibitory synaptic currents in individual neurons and thought to be characteristic of cortical circuits^35,36^.
A prerequisite for the analysis of neural manifolds are simultaneous measurements of activity across large neuronal populations. Basic features of manifolds may be assessed using distance measures that provide information about the relative separation and orientation of manifolds^32^. Moreover, geometrical features of neural manifolds relevant for pattern classification can be analyzed using the mathematical framework of manifold capacity, a measure that quantifies the degree of separability - or untangleness - of manifolds as a function of geometrical parameters^8,15,37^. Recently, this framework has been extended into GCMC (Geometric measures from Correlated Manifold Capacity) theory, which also considers the impact of multiple forms of neuronal co-variability on manifold capacity^38^. Thus, manifold capacity quantifies the amount of linearly decodable information per neuron as a function of parameters describing manifold geometry and correlations. Recently, GCMC theory has been used successfully to quantify computationally relevant, interpretable features of neuronal population activity in different brain areas and tasks^38^. Manifold capacity analysis can therefore be used to determine how learning modifies geometrical features of representational manifolds, and how these modifications collectively affect the classification of activity representing learned and novel inputs.
To explore how learning modifies representational manifolds we measured population activity in pDp after training juvenile or adult zebrafish in an odor discrimination task. Previous studies showed that odor-evoked activity in pDp is distributed, variable, modified by experience, and under strong inhibitory control^39–44^, consistent with observations in piriform cortex^19–21,23–25,29–31,45,46^. Moreover, in both brain areas, synaptic currents are dominated by recurrent rather than afferent input during an odor response^18,40,42,47^. In pDp, voltage clamp recordings demonstrated that excitatory and inhibitory synaptic currents in individual neurons are strong and co-tuned, providing direct evidence for precise synaptic balance^42^. The analysis of neuronal population activity in pDp therefore allowed us to examine how learning modifies representational manifolds in a precisely balanced autoassociative network.
Consistent with various previous observations we found no obvious signatures of fixed-point attractor dynamics in pDp. Behavioral odor discrimination training had only minor effects on the global amplitude and variability of odor responses in pDp. Nonetheless, training modified activity manifolds representing learned and related odors, with minimal effects on representations of dissimilar odors. Training modified multiple geometrical features that, in combination, enhanced the separability of manifolds representing task-relevant odors from each other and from manifolds representing novel odors (Extended Data Fig. 1). The separability of activity patterns was correlated with behavioral odor discrimination across trained individuals, indicating that representational manifolds directly contribute to learned behaviors. These results indicate that pDp stores task-relevant information not as discrete items but in continuous neural manifolds that improve classification of sensory information along relevant directions in coding space. Hence, representational learning in pDp, and possibly in other recurrent balanced-state networks, integrates sensory and semantic information into a continuous map, which may support distributed learning processes.
RESULTS
Basic features of neural dynamics in pDp
We measured odor-evoked activity in an ex-vivo preparation of the juvenile and adult zebrafish brain by volumetric 2-photon calcium imaging^48,49^ in transgenic fish expressing the calcium indicator GCaMP6s (Tg[alphaTubulin:GCaMP6s]) throughout the telencephalon and other brain areas^50^ (Fig. 1a,b). 3D scanning was performed by remote focusing with a volume rate of 7.5 Hz and calcium signals (ΔF/F) were transformed into firing rate estimates using CASCADE^43^. Activity across populations of neurons was represented by n-dimensional activity vectors, each representing the estimated firing rates of n neurons within a time bin. Most experiments were performed in juvenile fish (46 – 56 days post fertilization; 9.0 – 13.5 mm body length), which allowed us to measure activity across the majority of neurons in pDp. Consistent with previous results from adult zebrafish^39–42^, odor-evoked activity in juvenile pDp exhibited an initial phasic increase followed by a slowly decaying plateau (Fig. 1c,d), and responses of individual neurons were scattered throughout pDp without an obvious odor-related topography.
Previous results from pDp and piriform cortex failed to provide strong evidence for convergent attractor dynamics in response to novel or learned odors^22–26,30,39–42,45^. To confirm this conclusion, we examined two signatures of attractor states (Fig. 1e): (1) as a consequence of convergent dynamics, trial-to-trial variability is expected to be lower in output activity patterns than in input patterns, and (2) because attractor states are stable, activity is expected to persist for some time when the input is switched off abruptly.
To test the first prediction, we measured activity evoked by a panel of odors in pDp and in the olfactory bulb of the same juvenile zebrafish. Trial-to-trial variability was quantified as the cosine distance between activity vectors evoked by the same odors in different trials, averaged over a 3 s time window starting 1 s after response onset. Consistent with previous observations^39,40,51–53^, variability was high in pDp but low in the olfactory bulb (Fig. 1f), contradicting expectations for discrete attractor states.
To test the second prediction we used Tg[dlx4/6:Chr2YFP] zebrafish, which express channelrhodopsin-2 fused to YFP in superficial inhibitory interneurons of the olfactory bulb^54,55^. In these fish, olfactory bulb output could be silenced by targeted full-field illumination with blue light within 10–20 ms (Fig. 1g). We then performed whole-cell current- and voltage-clamp recordings from individual pDp neurons to examine the evolution of network activity after rapid silencing of olfactory bulb output during an odor response. Consistent with previous observations^40,42^, odor stimulation evoked a barrage of excitatory synaptic currents and a depolarization that occasionally resulted in action potential firing of pDp neurons. Silencing of olfactory bulb output during an odor response resulted in a fast hyperpolarization of pDp neurons with a time constant of 65 ± 23 ms, as determined by exponential fits (Fig. 1h). Direct injection of a step current hyperpolarized the same pDp neurons with a time constant of 52 ± 16 ms. Excitatory synaptic currents measured by voltage clamp recordings decayed with a similar time course (time constant: 60 ± 21 ms; Fig. 1i). Hence, network activity in pDp did not persist after silencing of sensory input but decayed on a timescale similar to the intrinsic time constant of pDp neurons. Together, these observations support the notion that odor-evoked activity in pDp does not show obvious signs of fixed-point attractor dynamics.
Experience-dependent modification of neural dynamics in pDp
To explore how odor representations in pDp are modified by learning we trained juvenile zebrafish expressing GCaMP6s^56^ in the telencephalon in an odor discrimination task using procedures developed for adult fish^57^ with minor modifications. During training, tanks were continuously perfused with fish water and two amino acid odors (CS^+^, C2^−^) were each added to the perfusion 9 times per day for 30 s. Presentation of the CS^+^, but not the C2^−^, was followed immediately by food delivery at a specific location. Fish were initially pre-trained as a group for 11 days before individual fish were further trained for 2 to 4 days in separate tanks (Methods; Fig. 2a). Learning was monitored by quantifying appetitive behavior of individual fish during odor presentation as described^57^. After training, appetitive behavior was significantly more pronounced in response to the CS^+^ than to the CS^−^ (Fig. 2b), as observed in adult fish^57,58^.
One group of juvenile fish (Arg^+^/Phe^−^; N = 9 fish) was trained using arginine (Arg) as CS^+^ and phenylalanine (Phe) as CS^−^ while a second group (Phe^+^/Arg^−^; N = 10 fish) was trained on the opposite association. A third group (Phe^+^/Trp^−^; N = 6 fish) was trained on Phe as CS^+^ and tryptophan (Trp) as CS^−^ (Fig. 2c). After behavioral training, odor-evoked activity was measured in pDp of all fish and compared to odor responses in a control group of naïve fish (N = 6). The odor panel comprised three amino acids (Phe, Arg, Trp) and three bile acids (TDCA, TCA, GCA), representing two classes of natural odors for aquatic animals^59^. In the olfactory bulb, activity patterns evoked by these odorants were modestly correlated within each chemical class but uncorrelated across classes^39^. Each stimulus was applied three times for 5 s.
As observed in adult pDp^39–42^, mean odor responses in juvenile pDp followed a phasic-tonic time course, with tonic components being somewhat more pronounced in trained fish (Fig. 2d). Mean odor responses to amino acids were slightly but significantly larger in each of the trained groups (Fig. 2e), which could be attributed in part to an overrepresentation of large odor responses in a subset of individual neurons (Fig. 2f). The cosine distance between time-averaged activity patterns evoked by the same odor in different trials was only slightly lower than the cosine distance between activity patterns evoked by different odors from the same odor category (Fig. 2g). In trained fish, cosine distances between activity patterns evoked by the CS^+^ and the CS^−^ were similar to the corresponding distances in naïve fish, while other cosine distances were slightly higher (Fig. 2g–j). Hence, training modified relationships between odor-evoked activity patterns but response variability remained high.
While high variability is atypical for fixed-point attractors it does not exclude chaotic attractor states. Modeling studies showed that chaotic attractor states tend to result in persistent pattern correlations, even when variability is high^4^. However, we found that correlations between activity patterns at different time points dropped rapidly after stimulus offset, without an obvious difference between naïve and trained animals (Extended Data Fig. 2). Hence, our observations do not support the hypothesis that training establishes convergent attractor dynamics representing specific odors.
Basic analysis of manifold geometry in a computational model
We next analyzed odor-evoked population activity in pDp as neural manifolds. We first used two distance measures, Euclidean distance (dE) and Mahalanobis distance (dM), to characterize basic geometrical relationships between neural manifolds in activity space (the state space where each dimension represents activity of one neuron). dE was defined as the distance between the centers of two distributions (e.g., the mean of data points representing a manifold). Hence, dE quantifies the separation of prototypical odor representations in state space. dM, in contrast, also depends on the shape of distributions: it measures the distance between a point (e.g., an individual activity vector) and a reference distribution (e.g., a neural manifold) in units that depend on the shape and extent (covariance pattern) of the reference distribution. dM between two manifolds is thus computed by averaging over the dM between each datapoint in one manifold and the distribution representing the other manifold. As manifolds typically have different shapes, dM in the direction from manifold A to manifold B (dM_A→B_) is usually different from dM in the opposite direction (dM_B→A_), providing information about manifold geometry (Fig. 3a,b). For example, dM_A→B_ > dM_B→A_ implies that A is more elongated than B along the A-B axis. Because dM is normalized by covariance it is closely related to the separability of two distributions.
We define a manifold representing a given odor as the distribution in state space of all activity vectors measured in response to the odor, pooled over time points and trials. To determine dE between two manifolds, we averaged activity patterns over each manifold, resulting in single points in state space (“manifold centers”). dE between representations of different odors was then determined as the Euclidean distance between manifold centers. Hence, dE is symmetric and depends both on the angular separation of manifold centers and their amplitudes (firing rates).
dM was determined between individual activity vectors x (activity evoked by an odor in an individual trial and time bin) and activity distributions Y (the set of activity vectors evoked by another odor in multiple trials and time bins). dM is defined as
where μ_Y_ and S_Y_ are the mean and the covariance matrix, respectively, of Y. Hence, dM is the effective distance between x and the mean of Y, taking into account the variability in Y. dM between manifolds representing odors X and Y in the direction X→Y (dM_X→Y_) was calculated by averaging over dM between each vector x in X and manifold Y. The same procedure was used to determine dM in the opposite direction (dM_Y→X_). Because dM_X→Y_ is usually not equal to dM_Y→X_ matrices quantifying dM between multiple manifolds contain information about their separation and geometry along selected directions (Fig. 3a,b).
We then jointly quantified dE and dM for manifolds representing different odors. As illustrated in Fig. 3bi, all dE and dM are equal when manifolds are equidistant and spherical with equal variance. When manifolds are scaled uniformly, dE changes while dM remains constant because Euclidean distance and variance co-scale (Fig. 3bii). When only the geometry of one manifold is modified, dE is unaffected but dM changes between specific subsets of manifolds (Fig. 3biii). Matrices of dE and dM therefore contain basic information about distances between representational manifolds and about their geometry in a subset of state space dimensions.
To predict effects of learning on dE and dM we simulated odor-evoked activity using recurrently connected networks of spiking neurons (Fig. 3c). Previous models of adult pDp^32,44^ consisted of 4000 excitatory and 1000 inhibitory neurons. Learning was assumed to enhance connectivity between small assemblies of strongly activated neurons (100 excitatory and 25 inhibitory neurons), which resulted in the separation of manifolds representing learned and related odors from other manifolds. To adapt these computational models to juvenile fish we reduced network size by a factor of four while maintaining precise synaptic balance. We then simulated responses to six input patterns with response amplitudes and correlations matching those of activity patterns evoked by the experimental panel of amino acid and bile acid odors across mitral cells of the olfactory bulb (Methods). Virtual odors were thus separated into inputs corresponding to amino acids (odors A1, A2, A3) or bile acids (B1, B2, B3) based on their correlations. In networks without assemblies, dE was higher between representations of A-odors than between representations of B-odors, and highest across odor classes (Fig. 3d). dM was also higher between A-odor than between B-odor representations (Fig. 3g). Across odor classes, however, dM was higher in the direction from vectors representing A-odors to reference distributions (manifolds) representing B-odors (dM_A→B_) than in the opposite direction (dM_B→A_) (Fig. 3g).
Introducing assemblies representing two A-odors increased dE between manifolds representing these “learned” and other odors (Fig. 3d,e), which can be attributed to a modest increase in firing rates of neurons within but not outside an assembly^32^. This effect generalized partially to the third A-odor, but not to B-odors. dM was increased for learned odors specifically in the direction dM_A→B_ (from A-odors to B-odors), indicating that manifold geometry changed non-uniformly (Fig. 3f,g). This asymmetric change in dM also generalized to the third A-odor, while no obvious effects were observed on distances between B-odors. In the computational model, the introduction of autoassociative memories therefore resulted in specific geometrical modifications of manifolds representing learned and related odors, as summarized in Fig. 3h.
Learning-related modifications of representational manifolds in pDp
To determine how discrimination training modified representational manifolds in pDp we first compared matrices of dE and dM in naïve and trained fish (Fig. 4). dE was increased between the CS^+^, the CS^−^, and the third amino acids (“AA3”), and between amino acids and bile acids, whereas dE between bile acids was not significantly different (Fig. 54a-c). This pattern was observed in all individual training groups. Matrices of dM were asymmetric, with higher dM in the direction from amino acids to bile acids (dM_AA→BA_) than in the opposite direction (dM_BA→AA_; Fig. 4d). In all groups of trained fish, dM was increased asymmetrically between specific classes of odor pairs (Fig. 4d–f): while dM was significantly increased between all amino acid representations (CS^+^, CS^−^ and AA3; AA-AA), dM between bile acid representations (BA-BA) was not significant different. Largest increases were observed between representations of amino acids and bile acids (AA-BA) in the direction from amino acid activity vectors to bile acid reference distributions (dM_AA→BA_), whereas dM in the opposite direction (dM_BA→AA_) remained almost constant. Shuffling of odor labels decreased all dE and dM and abolished significant differences between naïve and trained groups (Extended Data Fig. 3). Hence, effects of discrimination training on representational manifolds were consistent with model predictions.
These observations indicate that training had two major effects on representational manifolds of conditioned and related odors: first, manifold centers became more distant from each other and from other manifolds, as revealed by the increase in dE. Second, manifold geometry changed in a non-uniform, odor-specific manner. The finding that training increased dE_AA→BA_ but not dM_BA→AA_ implies that representational manifolds for amino acids expanded in the direction from amino acids to bile acids because the increase in dE must be accompanied by an increase in variability of similar magnitude in the same direction. In other directions, however, dM increased, indicating that variability increased less than dE. As dM directly reflects the linear separability of a given pattern from a reference distribution, learning enhanced the linear discriminability of activity vectors representing learned or related odors from manifolds representing other odors.
Changes in manifold geometry support pattern classification
The increase in dM in the direction from learned to other odors suggests that learning-related changes in manifold geometry support pattern classification. To further investigate this hypothesis we used the analytical framework of GCMC^38^. This framework quantifies the separability of manifolds using manifold capacity^8,15,37^, a metric that assesses how efficiently manifolds are stored in neural state space for retrieval via linear readout (Fig. 5a). Specifically, higher manifold capacity indicates greater discriminability of manifolds. In other words, a downstream neuron, modeled as a linear sum followed by thresholding, can distinguish between manifolds by accessing a smaller number of upstream neurons (see ref^38^ for additional interpretations of manifold capacity).
GCMC uses a set of parameters to describe geometrical features and correlations of manifolds (Fig. 6b). To compute manifold capacity, manifolds are repeatedly sampled from a distribution defined by these parameters and embedded in a state space of a given dimensionality. Individual manifolds are randomly assigned a binary label (1 or −1) and linear classifiers are trained to separate sets of manifolds with different labels. Manifold capacity is then defined as the maximum number of manifolds per dimension that can be embedded until linear classification fails (Fig. 5a). As GCMC theory analytically links geometrical parameters to classification error it provides a mathematical framework to understand how manifold capacity depends on geometrical features of representational manifolds and their correlations.
GCMC theory expresses manifold capacity as a function of five parameters that measure interpretable geometrical features of manifolds and their correlations (Fig. 5b): (1) effective manifold radius (related to “compactness”), (2) effective dimension (related to “flatness”), (3) center alignment (related to “pattern correlation”, i.e., the correlation between activity vectors representing manifold centers), (4) axes alignment (related to the “relative orientation” of manifolds), and (5) center-axes alignment (related to the interaction of “pattern correlation and relative orientation”). These measures provide a statistical description of representational manifolds that is linked to manifold capacity through a mathematical characterization of how linear readout interacts with manifold structure. The relationship between manifold capacity and parameters (1) – (4) is monotonic: manifold capacity increases when effective radius decreases, effective dimension decreases, effective center alignment decreases and effective axes alignment increases. The effect of center-axes alignment on manifold capacity (5) is more complex and depends on the other parameters.
We quantified manifold capacity based on sets of activity vectors across all pDp neurons within a 7 s time window starting 1 s after the odor response onset, pooled over the three trials for each odor. Across all odors and training groups, manifold capacity was significantly higher in trained (0.108 ± 0.017; n = 375 odor pairs from N = 25 fish; mean ± SD) than in naïve fish (0.098 ± 0.013; P = 2.9 × 10^−7^; n = 90 odor pairs from N = 6 fish; Fig. 5c). Shuffling manifold labels across all activity patterns substantially decreased the resulting capacity and abolished differences between naïve and trained groups (Naïve: 0.014 ± 0.0001; Trained: 0.014 ± 0.0001; P = 0.33; Fig. 5c). An increase in manifold capacity was consistently observed in each individual training group (Fig. 5c). To examine whether changes in manifold capacity were specific to an odor category we separately analyzed different classes of odor pairs (AA-AA, AA-BA, BA-BA). For all three classes, manifold capacity was significantly higher in trained than in naïve fish (AA-AA: P = 0.003; AA-BA: P = 3.4 × 10^−6^; BA-BA: P = 0.001; Fig. 5d). Largest relative differences in manifold capacity were observed for AA-BA pairs (10.80 ± 15.19 %), followed by AA-AA (8.84 ± 12.22 %) and BA-BA pairs (8.20 ± 8.83 %).
We next tested the hypothesis that training primarily modified manifolds representing the CS^+^ or CS^−^. We first compared pairs of manifolds representing conditioned odors to pairs of manifolds representing one learned amino acid (CS^+^ or CS^−^) and the third amino acid in the odor set (AA3). The relative increase in manifold capacity was significantly higher for classification of a conditioned odor versus AA3 (11.1 ± 11.7 %; P = 0.011e) than for classification of the CS^+^ versus the CS^−^ (4.4 ± 10.8 %; Fig. 5). These observations indicate that training enhanced the separability of both conditioned odors from other odors, as well as from each other. We therefore conclude that specific geometrical modifications of representational manifolds support pattern classification, particularly the discrimination of conditioned from other stimuli.
To further understand the underlying modifications of representational manifolds we analyzed the contributions of different geometrical parameters (Fig. 5b,f; Extended Data Fig. 4). In trained fish, the effective radius, dimensionality, center alignment and axes alignment were all significantly lower than in naïve fish (radius: 1.01 ± 0.04 [naïve] versus 0.98 ± 0.04 [trained]; P = 1.3 × 10^−10^; dimensionality: 19.0 ± 2.0 versus 17.9 ± 2.1; P = 1.0 × 10^−5^; center alignment: 0.87 ± 0.03 versus 0.85 ± 0.04; P = 5.2 × 10^−6^; axes alignment: 0.108 ± 0.031 versus 0.096 ± 0.028; P = 0.001), while center-axes alignment was not significantly different (0.032 ± 0.013 versus 0.035 ± 0.017; P = 0.095). The increase in manifold capacity was therefore due to decreases in effective manifold radius, dimensionality, and center alignment that outweighed opposite effects of axes alignment on manifold capacity (Fig. 5b,f).
The effective radius and dimensionality of manifolds were decreased for all classes of odor pairs (Extended Data Fig. 4), indicating that training reduced within-manifold variability relevant for classification in all odor categories. Center alignment was decreased significantly for AA-BA and BA-BA pairs, but not for AA-AA pairs. Axes alignment was decreased only for AA-BA, but not for AA-AA and BA-BA. Hence, training had somewhat different effects on manifolds representing conditioned inputs, related inputs, and dissimilar inputs, but decreases in effective manifold radius (“compactness”) and dimensionality (“flatness”) occurred consistently. Similar changes in manifold capacity, radius and dimensionality were observed in computational models after introducing assemblies (Extended Data Fig. 5). A decrease in effective radius indicates that manifolds become more “compact”, thus enhancing their separability, while a decrease in effective dimensionality implies that manifolds become more constrained in specific state space dimensions. A decrease in center alignment supports pattern classification by decorrelating representations. Our results indicate that these are the most prominent changes that improve the capacity of manifolds for pattern classification after training.
Experience-dependent modification of representational manifolds in adult fish
We next examined odor representations in adult zebrafish, where pDp is substantially larger. Effects of training may thus be expected to generalize less from conditioned stimuli to related stimuli because a larger network allows for more specific local modifications of manifold geometry. Adult zebrafish were trained^57^ using Trp as CS^+^ and alanine (Ala) as CS^−^. After training, activity patterns were measured in response to Trp, Ala, and two additional amino acids, serine (Ser) and histidine (His), that are structurally similar to the conditioned odors and evoke correlated responses in the adult olfactory bulb^51,60^. As observed in juvenile fish, training slightly increased odor responses and strong responses were observed more frequently (Extended Data Fig. 6a–c). In naïve fish, the similarity between activity patterns was generally low (high cosine distance), even between responses to the same odors in different trials (Extended Data Fig. 6d, left). In trained fish, the similarity of activity patterns was higher for repeated applications of the same odors but slightly lower for dissimilar stimuli (Extended Data Fig. 6d,e). Hence, odor representations in pDp were somewhat more distinct in trained fish but response variability remained high in comparison to the olfactory bulb^41,51–53^. Basic effects of training were therefore similar to those observed in juvenile fish, possibly with more odor-specific changes in pattern similarity.
As observed in juvenile fish, training significantly increased dE and dM in adult pDp (Extended Data Fig. 6f–k). The increase in dE was largest between the conditioned odors (Trp, Ala) and smallest between non-conditioned odors (Extended Data Fig. 6g,h). dM was increased predominantly in the direction from conditioned to other odors (i.e., from an activity vector representing a conditioned odor to a distribution of activity vectors representing another conditioned or a novel odor), with largest increases in the direction from the CS^+^ to non-conditioned odors (dM_Trp→Ser_, dM_Trp→His_; Extended Data Fig. 6j,k). These observations were largely abolished after shuffling of odor labels (Extended Data Fig. 2g–l). Hence, training resulted in an odor-specific, asymmetric increase in dM that primarily enhanced the discriminability of responses to conditioned odors.
Consistent with these findings, manifold capacity in adult pDp was significantly higher in trained as compared to naïve fish (Extended Data Fig. 6l). Parameter-specific analyses showed that the effective radius, dimensionality, center alignment and axes alignment were all significantly lower in trained as compared to naïve fish (Extended Data Fig. 6m). Hence, as observed in juveniles, the increase in manifold capacity can be attributed to changes in the effective radius, dimensionality and center alignment of representational manifolds, which collectively outweighed a negative contribution of axes alignment. Together, these observations indicate that training resulted in consistent geometrical modifications of representational manifolds in juvenile and adult zebrafish that enhance the capacity for pattern classification.
Manifold geometry predicts discrimination behavior
To examine whether the geometry of representational manifolds in pDp affects behavior we asked whether geometrical features of manifolds predict behavioral odor discrimination. The Euclidean distance between manifolds representing conditioned odors (dE_CS+,CS−) was positively correlated with the behavioral discrimination score (difference between appetitive behavioral response to CS^+^ versus CS^−^) across individual juvenile fish from all training groups but this correlation was not statistically significant (P = 0.11; Fig. 6a). A significant positive correlation was, however, found between dM_CS+→CS− (dM between a vector representing CS^+^ and the distribution representing CS^−^) and the behavioral discrimination score (r = 0.42; P = 0.04; Fig. 6b), while the correlation between dM in the opposite direction (dM_CS-→CS+_) and the discrimincation score was not statistically significant (P = 0.60; Fig. 6c). The highest significant correlation was detected between manifold capacity, calculated based on representations of the CS^+^ and CS^−^, and the discrimination score (r = 0.57; P = 0.003; Fig. 6d). Hence, unlike Euclidean distance, geometrical features of representational manifolds relevant for pattern classification predicted olfactory discrimination behavior. These observations indicate that geometrical modifications of representational manifolds contribute to learned changes in behavior.
DISCUSSION
Neural manifolds in an autoassociative network
We explored the organization of neural manifolds representing task-relevant information in an autoassociative memory network. Activity evoked by novel or conditioned odors did not exhibit obvious signs of classical or chaotic attractor states. Hence, our experimental observations do not support the classical view of primary olfactory cortex as an autoassociative memory network that classifies odors by convergent attractor dynamics. In principle, associative learning may be mediated by any mechanism that maps relevant inputs to defined activity subspaces according to their semantic relations. For example, activity patterns representing task-relevant sensory stimuli (inputs) may be associated with different task outcomes (semantic categories) by mapping them to separate neural manifolds (activity subspaces). Stimuli may thus be classified by a systematic organization of neural manifolds according to semantic categories, even if manifolds are not attractors. In fact, the formation of manifolds representing semantic categories is the basis for pattern classification by convolutional networks, which lack attractor dynamics by design^14,61^. Unlike such convolutional networks, however, biological memory networks are often recurrent, pre-structured, and modified by local learning rules. The small size of pDp together with computational modeling and GCMC theory allowed us to directly analyze representational manifolds and their plasticity in an autoassociative area of the vertebrate brain.
The notion that neural manifolds organize relevant information in neuronal state space implies that geometrical and statistical properties of manifolds are relevant for neural computation^8^. Quantitative analyses of features such as the effective radius, dimension and alignments of neural manifolds should thus drive insights into neuronal computations at the population level, similar to analyses of receptive fields at the single-neuron level. Consistent with this notion, we found that defined manifold features were systematically modified by discrimination training, demonstrating that representational manifolds are interpretable objects that capture computationally relevant structure in neuronal population activity. The finding that manifold capacity predicted discrimination behavior across individuals further indicates that manifold geometry is directly linked to behaviorally relevant neuronal computations. These observations imply that GCMC theory provides valuable tools to extract computationally relevant information about neural manifolds from large-scale measurements of population dynamics.
Representational manifolds and olfactory memory
Our findings indicate that pDp contains a continuous representation of odor space that is modified, but not discretized, by constraining activity onto neural manifolds as a function of experience. This scenario is consistent with multiple experimental observations in pDp and piriform cortex: (1) Training of animals in olfactory memory tasks led to the emergence of task-specific responses but the continuous representation of odor space observed in naïve animals was not reorganized fundamentally (Figs. 1,2)^25,27,29–31^. This observation is consistent with the notion that pDp or piriform cortex do not store discrete items of information but jointly represent odor space and semantic information in continuous maps^19,20,28^. (2) Experimental results in pDp and piriform cortex revealed only minor effects of training on the variability of single-neuron responses or global population activity^25,30,31^. These observations are consistent with the assumption that representational manifolds constrain neuronal activity only in a subset of state space dimensions. (3) Accurate odor discrimination requires reliable readout of information from population dynamics despite high variability of neuronal activity in pDp and piriform cortex. The notion of representational manifolds can reconcile these issues because activity is constrained to subspaces and, thus, information can be retrieved reliably by integrating activity over the manifold^32,62,63^. (4) In pDp^42^, and possibly also in piriform cortex^64^, network activity enters a state of precise synaptic balance during an odor response that may not support discrete attractor states. Representational manifolds can directly account for this observation and allow for pattern classification without attractor dynamics.
Discrimination training primarily enhanced the ability to classify activity patterns representing conditioned or related odors. Modifications of representational manifolds therefore contain task-relevant information that is combined with a map of odor space, resulting in joint representations of sensory and semantic information. These observations could, in theory, be explained by the formation of neuronal assemblies in synaptically balanced autoassociative networks^32^ but detailed insights into the biological mechanisms that define manifold geometry will eventually require reconstructions of memory networks at synaptic resolution.
Behavioral training consistently increased manifold capacity and, thus, optimized representational manifolds for odor classification. Increased manifold capacity could, in part, be attributed to higher “compactness” (decreased effective radius) and lower correlation (decreased center alignment) of representational manifolds, in agreement with increases in dE and dM. Manifold capacity was further enhanced by a decrease in dimensionality. In computational models, a similar decrease in dimensionality could be attributed, in part, to a modest amplification of activity in an odor-specific direction^32^. Such a directional amplification of activity may also contribute to the decrease in axes alignment observed in pDp, which had a negative effect on manifold capacity. Hence, training may modify multiple geometrical features of manifolds with opposing effects on manifold capacity. In combination, however, positive effects on manifold geometry outweighed negative contributions, resulting in an overall enhancement of the capacity for task-relevant odor classification.
Importantly, geometrical features of manifolds, particularly manifold capacity, predicted behavioral odor discrimination. Hence, geometrical features critical for pattern classification were directly related to behavioral odor discrimination, consistent with the hypothesis that semantic information accumulated in the geometry of representational manifolds is read out by neural classifiers controlling behavior.
Putative computational functions of representational manifolds
The representation of sensory and semantic information by neural manifolds supports task-dependent pattern classification by facilitating access to relevant information. In computational models, for example, information about learned odors can be retrieved efficiently by integrating activity over assembly neurons^32,63^. In our analysis of dE and dM, we found that training enhanced the ability to classify conditioned odors even when activity was read out from subsets of neurons that were selected solely based on activity, indicating that information relevant for pattern classification can be retrieved by simple, biologically plausible mechanisms. The observed increase in manifold capacity further implies that fewer neurons are required for efficient classification of conditioned odors after training because the amount of relevant information per neuron increases^15,38,63^. One computational function of experience-dependent changes in manifold geometry may therefore be to facilitate the classification of meaningful sensory inputs. In addition, modifications of continuous representational manifolds support other computations such as the evaluation of metric relationships between relevant patterns^32^.
Interpretable changes in manifold capacity have recently been described also in brain areas of other species^38^. For example, manifold capacity was found to increase along the ventral stream of the visual system in datasets from monkeys and humans. In convolutional networks trained on image classification, manifold capacity increased systematically along layers prior to the final classification layer^14^. These studies further support the assumption that neural manifolds are informative objects critical for neuronal computation, implying that the systematic analysis of manifold geometry can uncover principles of information processing in the brain.
Since pDp does not exhibit persistent activity it is unlikely to function as an integrator network. Moreover, consistent with recent observations in piriform cortex^25^, pDp does not appear to classify odor objects by convergent attractor dynamics even after learning. Hence, pDp is functionally more closely related to the intermediate (“representational”) layers than to the final classification layer of deep convolutional networks. We therefore propose that pDp establishes “semantic maps” of odor space to support behaviorally relevant sensory pattern classification and potentially other computations as part of a larger network. This view is consistent with the concept of olfactory cortex as a memory-related structure that comprises multiple recurrent brain areas^19,20^. More generally, representational learning by experience-dependent reorganization of neural manifolds may be critical for cognitive processes that are mediated by distributed computations across the pallium.
METHODS
Animals and transgenic lines.
Zebrafish (Danio rerio) were raised and kept as groups in a standard facility at 26.5–27.5 °C on a 14/10 h light/dark cycle. Juvenile fish were kept in 1.1 L tanks (Tecniplast ZB11TK). Olfactory conditioning of juvenile fish in their home tank started 28–38 days post fertilization (dpf). Juvenile fish used for calcium imaging experiments were 46 – 56 dpf with a body length of 9.0 – 13.5 mm and not selected for gender, which is not yet determined at this stage. Adult fish were 5–6 months old and not selected for gender. The naïve group consisted of 4 females and 4 males while the trained group consisted of 5 females and 4 males. Juvenile fish expressed the calcium indicator GCaMP6s pan-neuronally under the control of the alpha-tubulin promoter (Tg[alphaTubulin:GCaMP6s]^50^). Adult fish were wildtype except for Tg[dlx4/6:ChR2-YFP]^54,55^ used in optogenetic experiments. All experimental protocols were approved by the Veterinary Department of the Kanton Basel-Stadt (Switzerland).
Ex-vivo preparation of the zebrafish brain.
Ex vivo preparation of the juvenile zebrafish brain.
In juvenile zebrafish, one of the olfactory bulbs and the ipsilateral side of the telencephalon were exposed to provide optical access. Before dissection, juvenile zebrafish were transferred into a beaker containing c.a. 10 mL system water at room temperature. The beaker together with the fish was cooled with ice to 4°C to anesthetize the fish until immobile. Heart rate was visually monitored and anesthesia was ensured by pinching the tail fin. The fish was the transferred into ice-cold teleost artificial cerebrospinal fluid (ACSF: 124 mM NaCl, 2 mM KCl, 1.25 mM KH_2_PO_4_, 1.6 mM MgSO_4_, 22 mM D-(+)-glucose, 2 mM CaCl_2_ and 24 mM NaHCO_3_, pH 7.2.) bubbled with O_2_/CO_2_ (95%/5%)^65^ and fully immersed in this medium during dissection. Peripheral structures (eyes, jaws, gills) were removed and the head was detached from the body. The head was pleased on a coverslip, the exposed lateral side was oriented upwards, and the preparation was stabilized using tissue glue (3M Vetbond Tissue Adhesives, No.1469SB). Fine forceps and scissors were used to expose the olfactory bulb and telencephalon by removing soft cartilage and connective tissue without damaging the olfactory nerve. Care was taken not to obstruct the nostrils. For fish with body length above 12 mm, contralateral bones were partially removed to increase exposure of the brain to aerated ACSF. The preparation was transferred together with the coverslip to a custom flow chamber for two-photon imaging that was continuously perfused with ACSF at room-temperature.
Ex vivo preparation of the adult zebrafish brain.
The ex vivo preparation of adult zebrafish for the electrophysiology experiment was performed as described^42,49^. Briefly, fish were anesthetized by cooling to 4°C and decapitated in ACSF. After dissection of the jaws, the eyes and the bones covering the ventral telencephalon, the dura mater over pDp was removed with fine forceps. The ventral bones covering the olfactory bulbs were removed if necessary (electrophysiological recordings from mitral cells, optogenetic silencing). After surgery, the preparation was slowly warmed up to room temperature under constant perfusion with ACSF as described for juvenile fish.
Injection of a synthetic calcium indicator into adult pDp.
Fish were mounted in a custom-made holder for imaging in a horizontal orientation to ensure high reproducibility of the injection procedure as described (REF Frank et al., NN paper). Bolus loading of Oregon Green 488 BAPTA-1-AM (OGB-1; ThermoFisher Scientific) was performed as described^66^ with minor modifications. 50 μg of OGB-1-AM was dissolved in 30 μL of DMSO/Pluronic F-127 (80/20; ThermoFisher Scientific), vortexed for 1 min and sonicated for 15 min before being stored in 4 μL aliquots at −20°C. Prior to each experiment, an aliquot was diluted in 15 uL of ACSF, vortexed for 1 min, sonicated for 5 min and purified for 5 min at 5000 rcf in centrifugal filter units (PVDF membrane with 0.22 μm pore size; Millipore). A slowly tapering glass pipette was pulled from a borosilicate glass capillary (borosilicate glass capillaries, 1 mm outer diameter, wall thickness 0.21 mm, capillary length 100 mm; Hilgenberg article number 1810021) with a laser puller (P-2000; Sutter). The tip was broken under a microscope (MF-900 Microforge; Narishige) to a tip diameter of approximately 4 μm.
Pressure injections were targeted using a non-resonant two-photon scanning microscope with a 20x objective that provided both a fluorescent channel for detection of OGB-1 (PMT H7422P-40MOD, Hamamatsu) and a transmitted light channel (PSD, position-sensitive detector) as described^41^. pDp was identified based on its location in the lateral telencephalon, posterior to the prominent furrow. One primary injection was made ~210 μm dorsal from the ventral-most aspect of Dp and ~130 μm from the lateral surface of Dp in two intervals of each ca. 2 min. A second injection was performed in the same entry channel of the pipette but less deep (ca. 180 μm dorsal, 60 μm lateral) for 1–2 min. Dye injection was monitored by snapshots of multiphoton images. Pressure was adjusted to prevent fast swelling of the tissue that could be caused if the applied pressure was too high. The entire injection procedure typically took 6–10 min.
After injection, the pipette was retracted from the brain and the fish head was quickly (<2 min) transferred to another holder for imaging in a sagittal orientation using a 2-photon microscope equipped for volumetric resonance scanning. The approximately sagittal orientation allowed for improved optical access to areas of pDp close to the lateral brain surface but deeper from the ventral surface. The orientation of the brain was adjusted to minimize optical aberrations due to tissue curvature. Odor application and calcium imaging started ca. 1 h after dye injection.
Optogenetic stimulation of the olfactory bulb.
Blue light was targeted to the ipsilateral olfactory bulb through an optical fiber (200 μm diameter, Thorlabs) using a 457 nm laser (500 mW before attenuation) as described^49^. Laser intensity was adjusted to obtain 200 μW at the fiber tip. Optical stimulation was restricted to the olfactory bulb with possible minor off-target effects onto adjacent areas.
Odors were applied for 10 s through a tube in front of the ipsilateral nostril using a peristaltic pump system as described^42^. The onset of optical stimulation (300 ms) was targeted at the plateau phase of the odor response in pDp, typically 400–500 ms after response onset. Current clamp recordings from superficial mitral cells (n = 4) showed that the latency between blue light onset and cessation of action potential firing was approximately 10–20 ms. Recovery of mitral cell activity was observed approximately 500 ms after the end of the light pulse.
Electrophysiology.
Patch-clamp recordings of neurons in pDp were performed as described^42^ using borosilicate pipettes (pipette resistance: 4–8 MΩ) and a Multiclamp 700B amplifier (Molecular Devices). Pipettes were filled with intracellular solution containing (in mM): 132 Cs methanesulfonate, 10 Na_2_-phosphocreatine, 4 MgCl_2_, 4 Na_2_-ATP, 0.4 Na-GTP, 5 L-glutathione, 0.1 EGTA, and 10 HEPES (pH 7.2, 300 mOsm; all from Sigma). Current-clamp recordings were performed with an intracellular solution containing (in mM): 129 K-gluconate, 10 HEPES (free acid), 0.1 EGTA, 4 Na_2_-ATP, 10 Na_2_-phosphocreatine, 0.3 Na-GTP, 5 L-glutathione, and 13.1 KOH (pH 7.2, 305 mOsm; all from Sigma).
Neurons were targeted using the shadow-patching technique^67^ as described^42^, with 0.05 mM Alexa488 or Alexa594 (Invitrogen) included in the internal solution using a custom-designed video-rate multiphoton microscope^48^. When the dura mater over Dp was not completely removed, the pipette was advanced through the dura mater with transient high pressure (100 mbar) to avoid contamination of the pipette tip. Before forming a seal, neurons were approached within the tissue using low pressure (20 mbar). After break-in, series resistance and input resistance were continuously monitored. To measure the cellular time constant of Dp neurons by direct hyperpolarization, brief voltage steps (500 ms, step size ΔV = −5 mV) were applied through the patch pipette during voltage clamp experiments.
Voltage traces triggered on the onset of optical stimulation or hyperpolarizing current injections were averaged for each neuron and normalized by subtracting the pre-event current (mean of the 200 ms window before stimulation onset) and dividing by the asymptote (400–500 ms after stimulation onset). The resulting traces were fitted with an exponential function to estimate the decay time constant. An equivalent procedure was used to estimate decay time constants from normalized current traces.
2-photon calcium imaging.
2-photon calcium imaging was performed using a custom-built multiphoton microscope^48^ using a 20x objective (NA 1.0; Zeiss) and custom-written Scanimage software^42,68^. Laser pulses for two-photon excitation were centered around 930 nm, with a temporal pulse width of 180 fs below the objective as measured with an autocorrelator (CARPE; APE Berlin). Fluorescence was detected by a GaAsP photo-muliplier tube (PMT, H7422P-40MOD; Hamamatsu) without bandpass filtering of the emitted light to maximize fluorescence yield. The average laser power was gradually adjusted to 42–52 mW for juveniles and 23–24 mW for adults at imaging planes closest to the brain surface and 4 mW higher for deepest imaging planes using a Pockels cell (350–80LA; Conoptics) synchronized with the voice coil motor used for fast z-scanning.
Imaging was performed in 8 planes (256 × 512 pixels each) at 7.5 Hz as described^48^. Ca. 7.3 of these 8 planes were scanned during the linear trajectory of the z-scanning, while the remainder was acquired during the fast flyback of the z-scanning unit. The peak-to-peak maximum extension of the imaging volume was ca. 150 μm in juveniles and 100 μm in adults, whereas the extent of the FOV in x and y was around 250 μm and 125 μm in juveniles and 200 μm and 100 μm in adults, respectively, slightly varying with the relative z-position of the plane due to remote z-scanning^48^.
After every 1 or 2 trials, the microscope stage was repositioned in order to compensate for potential drifts. For this purpose, a small z-stack of ± 6 μm around the current location (step size, 2 μm) was acquired and the optimal shift in x, y and z was determined based on the correlation with a reference stack. Sub-resolution interpolation using a Gaussian fit of the correlation values allowed to achieve a correction accuracy well below the step size of 2 μm. For most experiments, the corrected drift was substantial over the time course of an experiment. In juveniles, the total drift ranged in 10–15 μm in pDp and 5–10 μm in the OB in z-direction, each region recorded for 80–90 min. In adults, the total drift ranged in 18–25 μm in z-direction over a full experiment within 50–60 min. During each trial, average projections of each imaging plane were created and inspected for artifacts (e.g., degraded signal due to bubbles or imaging plane drifts with respect to the reference stack). Defective trials were discarded and re-acquired.
Odor application.
Amino acids (Ala, Arg, His, Phe, Ser, Trp; Sigma) were prepared as 100x stock solutions in double-distilled water, vortexed, sonicated, stored at −20°C, and diluted to a final concentration of 10^−4^ M in ACSF immediately before the experiment. Bile acid odorants (TDCA, TCA, GCA; Sigma) were also prepared as 100x stock solutions and diluted to a final concentration of 10^−5^ M. Food extract, which was used as a positive control stimulus, was generated by heating fish food (Gemma Micro 300) in double-distilled water and filtering the product through 0.22 μm pore size filters. The product was then stored as a stock solution and diluted 500x before the experiment.
Odors were applied to the nasal epithelium through a constant stream of ACSF using a computer-controlled odor-application system based on peristaltic pumps as described^42^. In experiments using juvenile fish, a circular symmetric 9-channel plastic manifold (Darwin microfluidics Manifold 9 Ports 1/4–28 PEEK, 1/16” OD) was used to combine the flow of multiple odor channels to the main ACSF perfusion channel. A bubble trap (Diba Omnifit^®^ Bubble Traps, 21940–38) was placed in the tube delivering ACSF to the manifold. In experiments using adult fish, odors were applied using a linear manifold as described^42^. The time of response onset was determined by application of fluorescein in the absence of fish and fine-adjusted based on neural activity measurements in each experiment. Both in juvenile and adult experiments, 1–3 odor applications were performed prior to the start of data acquisition to identify the odor-responsive brain region. Odors used in these pre-trials were food odor (adult fish) or a bile acid (juvenile fish).
To measure odor-evoked activity in pDp of juvenile fish, 6 odor stimuli (Phe, Arg, Trp, TDCA, TCA, GCA), a control stimulus (ACSF), and another control with no odor delivery were applied in a pseudo-random sequence. This procedure was repeated three times with different pseudo-random sequences. Odors were applied for 5 s with an inter-trial interval of at least 2 min. The same odor application procedure with new pseudo-randomized application sequences was repeated to measure odor-evoked activity in the olfactory bulb of the same fish. In experiments on adult fish, an equivalent protocol was used to measure activity in pDp using Trp, Ala, His, Ser, Food odor, TDCA, and ACSF as stimuli. Odors were applied for 10 s.
Odor discrimination training.
Adult zebrafish were trained in an odor discrimination task for six days as described^57^. Juvenile fish were trained using a similar procedure after pre-training in the home tank as a group (Temiz et al., manuscript in preparation). Naïve fish used as controls were obtained from the same parental stock. During the group training phase, 10–15 juvenile zebrafish (28–38 days post fertilization) were transferred into a housing tank (Tecniplast 3.5L tank, ZB30TK) that was continuously perfused with system water (26.5°C, 120–130 ml/min flow rate). During each trial, a CS^+^ or CS^−^ odor was delivered for 30 seconds using an Arduino-controlled peristaltic pump (Adafruit Peristaltic Liquid pump). Following CS^+^ delivery, fish food (Tetra TetraMin Flocken) was dispensed into a feeding ring floating in a specific location after a 30-second delay, whereas no food was dispensed following presentation of the CS^−^. Fish received 7 CS^+^ and 14 CS^−^ applications per day, presented in an alternating sequence with 40-minute intervals.
After 11 days of group training, fish were transferred individually to small tanks (Plastic-Haus AG, 12×6×6 cm) and allowed to acclimate for 1–3 days. Subsequently, each fish underwent individual training using the same procedure as for adult animals^57^ with minor modifications to account for differences in body size. CS^+^ and CS^−^ odors were the same as during the group training phase. Each odor was delivered in an alternating sequence for 9 trials with a 20-minute intertrial interval. Swimming behavior was monitored by 3D video imaging throughout the individual training phase, which lasted 2–4 days. The appetitive behavior score ζ for each odor was computed as described based on quantitative analyses of swimming speed, the z-position in the water column, the presence in the reward zone, water surface sampling, the distance to odor inflow tube and rhythmic circular swimming during the 30 seconds between odor onset and reward delivery^57^. ζ scores are combined and normalized measures similar to z-scores that quantify appetitive behavior in response to each odor. The behavior discrimination score was computed as the difference between the ζ scores for the CS^+^ and CS^−^, summed over trials and divided by the number of individual training days.
Processing of image data.
Automatic region of interest (ROI) extraction was performed using StarDist (https://github.com/stardist/stardist) combined with manual correction to segment neuronal somata in the juvenile calcium imaging dataset. The model was trained with 20 RGB images (512 × 512), each color channel containing the anatomy image, ΔF/F map, and spatial correlation map as input and manually segmented neuron somata as ground-truth output. The ground-truth data was obtained from the same fish line using the same microscope as in the juvenile calcium imaging experiments. The threshold for ROI detection in StarDist was adjusted to avoid cross-neuron signal contamination. The ROIs detected by StarDist were imported into a custom MATLAB program (https://github.com/fmi-basel/neuRoi). ROIs were aligned between trials with fast Fourier transform (FFT) cross-correlation and individual neurons were manually redrawn or deleted depending on the quality of trial-by-trial alignment.
The raw fluorescence traces for each neuron were extracted by averaging the pixel intensity within the ROI and subtracting the background fluorescence. The F_0_ for each neuron was taken as the median fluorescence intensity within a 2 s time window prior to stimulus onset.
Spike probability was inferred from ΔF/F traces using the CASCADE algorithm^43^. Global_EXC_7.5Hz_smoothing200ms_causalkernel was chosen as the spike inference model. The noise level was determined by pooling ΔF/F traces during a 2s time window from all trials and used to choose the most appropriate noise level parameter used in the inference function. The output of CASCADE is the spike probability within the 133 ms time bin corresponding to the 7.5 Hz frame rate. The spike probability of the CASCADE algorithm was converted to firing rate by multiplication with the frame rate.
Segmentation of neuronal somata in the adult calcium imaging dataset was performed manually using a custom MATLAB program (https://github.com/PTRRupprecht/Drawing-ROIs-without-GUI). Spike probability was inferred using the OGB_zf_pDp_7.5Hz_smoothing200ms model from CASCADE.
Spiking network model of pDp.
The spiking network model of pDp was the same as in a previous study^32^ except for the number of neurons and synaptic weights, to account for the smaller size of the juvenile zebrafish brain. The model of pDp consists of 1000 excitatory (E) and 250 inhibitory (I) adaptive leaky integrate-and-fire neurons with conductance-based synapses. The neurons in pDp receive inputs from 1500 excitatory mitral cells in the OB.
Neuronal dynamics.
The membrane potential Vx of a neuron x in pDp evolved according to:
where X is the excitatory or inhibitory population to which x belongs. C_X_ is the membrane capacitance, g_rest,X_ is the leak conductance, and E_rest,X_ is the resting potential. The capital X in the subscripts means that the same values apply to all neurons in population X. g_OB,x_ is the conductance of the synapse from an olfactory bulb input to neuron x and g_P,x_ is the synaptic conductance from population P to neuron x. E_P_ is the reversal potential of a synapse of population P. z_x_ is the adaptation current if x is an excitatory neuron^69^:
with z_x_ set to z_x_ + b after each spike. Parameters a, b and τ_a_ were the same as in a previous model^32^. The conductances g_P,x_ evolved according to
where τ_syn,P_ is the synaptic time constant, w_yx_ is the synaptic weight from neuron y to neuron x, and t_spike,y_ is the spike time of neuron y.
Olfactory bulb input.
The input from the olfactory bulb to pDp was modeled by directly modulating the baseline 6 Hz firing rates of mitral cells in the olfactory bulb. Odor-evoked activities in the olfactory bulb were simulated by increasing the firing rate of 150 mitral cells and decreasing the firing rate of another 75 mitral cells. The activity pattern of mitral cells for each odor was generated to approximate the response amplitudes and pattern correlations observed in experiments.
Network connectivity and E/I assemblies.
Connections between two neurons x and y from populations X and Y, respectively, were drawn from a Bernoulli distribution with parameter p_XY_. The strength of all existing connections was set to w_XY_. While parameters p_XY_ are the same as in a previous model^32^, w_XY_ were fitted to account for the reduced network size. To explore a broad parameter space, 4 connectivity matrices were fitted, each with 2 instantiations drawn, with parameter values shown in Table 1. The simulation results from these 8 instantiations were used in further analysis.
E/I assemblies were introduced in pDp as described^32^. The E-assembly comprised the 100 excitatory neurons receiving the highest degree of inputs from the mitral cells activated by the associated odor. The corresponding I-assembly comprised the 25 inhibitory neurons receiving the highest degree of inputs from the E-assembly. E/I assemblies were formed by increasing the number of connections within the E-assembly and between the E- and I-assembly. To maintain a constant number of connections, an equal number of connections was eliminated randomly between non-assembly neurons and assembly neurons. The network with assemblies in Fig. 3 has two sets of E/I assemblies corresponding to odor A1 and A2.
Manifold construction and distance between manifolds.
In experimental data, neural manifolds were constructed as follows: In each fish, for each odor, the spike probabilities of each neuron at each time point within the 3 s analysis time window from 3 trials were pooled, resulting in a total of 72 datapoints per manifold for each odor and fish (24 timepoints × 3 trials). The time window started 1 s after response onset to include the peak and the plateau of the response. Different starting points (2 s to 6 s with interval 1 s;odor application starting at 2s) and durations (3 s, 5 s, 7 s) of the analysis window gave similar results.
In simulated data, manifolds were constructed by computing spike rate in 100 ms time windows and pooling the activity patterns across all excitatory neurons, including the assembly and non-assembly neurons, from a 2 s time window from 4 trials for each odor.
Distance between each pair of manifolds was computed in each fish. To ensure that the covariance matrix is invertible we selected subsets of 70 neurons, repeated the sampling 50 times, and averaged the resulting dM over the 50 repeats. dE was computed using the same procedure.
dE between two manifolds X and Y was defined as:
where and are the centers of the manifolds X and Y, respectively. dM with X as sample and Y as reference was defined as:
where |X| is the number of points in the manifold X, x is a point in the manifold X, μY is the center of the manifold Y, and SY is the covariance matrix of the manifold Y.
To analyze distances between a pair of manifolds after shuffling of odor labels, data points of both manifolds were pooled and randomly assigned to two new sets of equal size. dE and dM were then computed between these sets. This procedure was repeated 50 times and results were averaged.
Manifold capacity analysis.
Data preprocessing and capacity estimation.
Manifolds were constructed from experimental data as described above using an analysis time window of 7 s starting from the onset of the odor response. The time window was longer than for the analysis of dE and dM because manifold capacity can quantify separability even when manifolds exhibit curvature. Shorter time windows or different starting time points produced similar results. To ensure linear separability as required by GCMC, we subsampled neurons and time points, to make sure the number of neurons is larger than the number of activity patterns, so that the dimension of the space is large enough for a valid linear separator to exist. In juvenile fish, 700 neurons and 140 activity patterns were sampled for each manifold, while in adults, 400 neurons and 80 activity patterns were sampled. The number of neurons for juvenile and adults were chosen to match the minimal number of neurons recorded in each developmental stage. Capacity analysis of simulated data was performed using a 2 s time window, 1000 neurons, and 50 repeats.
Capacity was computed separately for each pair of manifolds representing an odor pair using the GCMC framework^38^. Subsampling was repeated 50 times for each manifold pair and capacity and geometric measures were averaged over the 50 repeats. As a control, manifold points were shuffled by pooling data points from both manifolds and re-assigning labels randomly prior to the calculation of capacity. The global centering and bias parameters were both set to true during capacity computation, which translated the mean of the manifold centers to the origin and allowed for the hyperplane classifier to have an offset from the origin^38^.
Manifold capacity: overview.
The algorithm for computing manifold capacity is detailed in^38^. Here we briefly describe the key steps. In the manifold capacity theory (MCT), a manifold is modeled as a convex set residing in an N-dimensional state space. It is represented by its center, its K axes stretching out from the manifold center, and the set of coordinates with respect to the manifold axes, specifying the location of the points belonging to this manifold. That is, denoting the manifold by M, a point x in the manifold can be expressed as
where u0 is the manifold center, ui ∈ R^N^ for 1 ≤ i ≤ K < N is the set of manifold axes, and s ∈ R^K^ is the coordinate of x with respect to the manifold axes. We use S ⊂ R^K^ to denote the set of all possible coordinates of points contained in the manifold.
Consider P manifolds simultaneously residing in the N-dimensional state space. Each manifold M^μ^ is indexed with μ for 1 ≤ μ ≤ P. Consider a set of dichotomies . As derived from MCT^38^, the manifold capacity α is:
where DT is the zero-mean Gaussian measure and T is a random vector with dimension N. Finally, A is a convex set of vectors reflecting the geometry of the manifold shapes:
The set Ay is the collection of all linear classifiers for the dichotomy y, where V corresponds to the normal vector that uniquely defines a separating hyperplane.
The overline in equation (1) denotes the average with respect to the labels y.
Effective geometric measures for linear classification.
According to the MCT, capacity is affected by geometric measures including effective radius, dimension, and alignments. “Effective” emphasizes that the measures are analytically connected to the capacity value; hence, this approach can be used to analyze geometrical changes underlying changes in manifold capacity. Thus, manifolds with different intrinsic geometries can have the same effective measures due to their relative configurations.
Detailed mathematical expressions of these measures are introduced in^38^. Here we only give an intuitive overview of how they are computed.
To understand the effective geometric measures, it is important to introduce the concept of anchor points. The capacity formula (Equation 1) is a Gaussian average (over T) of a quadratic programming problem (a convex optimization problem). According to the strong duality theory, the solution of a quadratic program is equal to the solution of its dual problem. In this case, the dual problem is a function of the points from the manifolds. Concretely, let , the duality theory gives that there exists a function g such that F(T) = g(T, xx^1^(T), …, x^P^(T)) for some x^μ^(T) ∈ M^μ^ for each μ. These points x^1^(T), …, x^P^(T) are known as anchor points. As a result, the randomness of T Induces a distribution over each manifold. These distributions are the anchor point distributions. Notice that the anchor point distributions are analytically connected to the capacity value by the following equation:
Finally, the effective geometric measures from GCMC are simply geometric terms extracted from the function g in the above formula. For reference, the following are the intuitive definitions of the geometric measures. We refer interested readers to the GCMC paper for a comprehensive understanding.
- Effective radius measures how far the points spread away from the manifold center, and is normalized by the length of the manifold center. It can be seen as the amplitude of internal variability of the manifold.
- Effective dimension measures the number of directions along which the points extend, with respect to the manifold center. It represents the degree-of-freedom of the variability.
- Effective center-alignment measures the correlation between the centers of different manifolds. It is the cosine similarity under the anchor-point geometry.
- Effective axes-alignment measures the correlation between the internal axes of different manifolds.
- Effective center-axes alignment measures the correlation between internal axes of one manifold and the center of another manifold.
These geometric measures were computed simultaneously with the capacity. Each odor pair from each fish is characterized with these five measures for further statistical analysis.
Statistical and correlation analysis.
Comparisons between naïve and trained groups were performed using a Mann–Whitney U test. For multiple comparisons between naïve and trained subgroups we used a nonparametric Kruskal–Wallis test followed by a post hoc Dunn’s test for multiple comparisons with the naïve group. Reported P values are adjusted for multiple comparisons with Bonferroni correction.
The correlation between variables was evaluated using ordinary least squares (OLS) linear regression (implemented with statsmodels package in Python), treating one variable as the independent predictor and the other as the dependent outcome. The slope, coefficient of determination (R2), and p-value from the regression model were reported to quantify the strength and statistical significance of the relationship.
In all statistical tests (implemented in Python with scipy.stats or scikit_posthocs package), P < 0.05 was considered statistically significant. In graphical displays, standard significance levels are indicated with asterisks (n.s.: P ≥ 0.05; * P < 0.05; ** P < 0.01; *** P < 0.001).
Extended Data
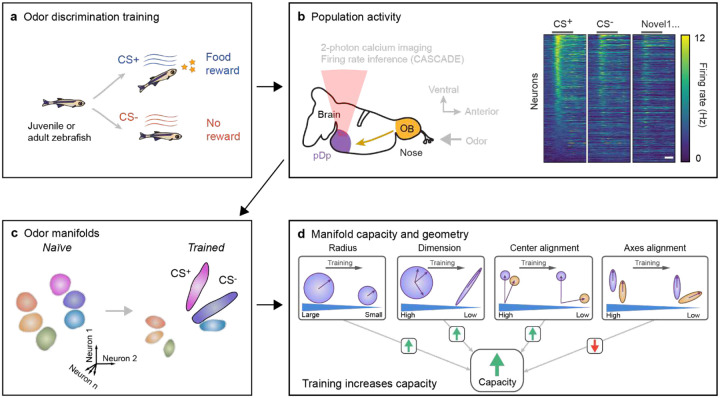
Graphical summary of approach and main results.a, Juvenile or adult zebrafish were trained in an odor discrimination task. b, Population activity evoked by conditioned (CS+, CS−) and novel odors was measured in telencephalic area pDp, the homolog of piriform cortex, using 2-photon calcium imaging and firing rate inference. c, Training enhanced the separation of neural manifolds representing conditioned odors (CS+, CS−) from representations of other odors. d, Analyses based on manifold capacity theory demonstrated that training enhanced the linear separability, or “untangledness”, of manifolds representing conditioned odors. Increased manifold capacity (separability) could be attributed to changes in multiple geometrical features: (1) a decrease in the effective radius (manifolds become more “compact”), (2) a decrease in dimensionality (manifolds become more “flat”) and (3) a decrease in center alignment (manifolds become more decorrelated). A concomitant decrease in axes alignment (manifolds become less aligned) had a negative effect on manifold capacity that was, however, outweighed by changes in the other features. The increase in manifold capacity was correlated to odor discrimination performance across individuals, indicating that geometrical features of representational manifolds are closely related to the behavioral readout of odor representations.
Odor-evoked activity is not persistent.a, Pearson correlation between activity patterns at different time points in pDp (top) and in the olfactory bulb (bottom), averaged over all Arg trials of naïve (left; N = 6) and Arg+/Phe−-trained fish (right; N = 9). b, Mean Pearson correlation between activity patterns at different time points and the mean activity during the odor application period (gray shading) in pDp (top) and in the olfactory bulb (bottom). Curves show correlations of activity evoked by four odor stimuli (Phe, Arg, Trp, TDCA) in each training group (colors; mean ± SD).
*Distances between pairs of manifolds after shuffling manifold labels.a, Pairwise dE between manifolds constructed after shuffling labels of datapoints representing different odors. Left: naïve juvenile fish (matrix averaged over N = 6 fish); right: Arg+/Phe−-trained fish (N = 9 fish). The shuffling was performed such that for each pair of manifolds, all points from both manifolds were polled and randomly split into two new manifolds. In all panels, color scales are the same as in the figures showing corresponding results without shuffling (Fig. 4). b, Difference between dE matrices from trained and naïve fish after shuffling. Trained fish were from different training groups were combined (N = 25 in tital). Amino acid odors were reordered by reward assignment (CS+, CS−, third amino acid) prior to averaging over training groups. c, dE between manifolds representing different odor classes (AA: amino acids; BA: bile acids) after shuffling of data points from naïve (N = 6) and trained (N = 25) fish. Pale box plots show dE before shuffling (same data and axis scaling as in Fig. 4). For all panels: ns, P ≥ 0.05; *, P < 0.05; **, P < 0.01; **, P < 0.001; see Supplementary Notes for more details. d-f, analysis of dM after shuffling of datapoint labels. Same procedures and plot conventions as in a-c. g-l, same analysis of dE and dM for data from adult fish after shuffling of datapoint labels. Color and axis scales are the same as in the figures showing corresponding results without shuffling (Extended Data Fig. 6). In i and l, manifold distances were analyzed separatedly for conditioned odors (Cond.; Trp and Ala) and non-conditioned odors (Non-cond.; Ser and His).
*Further analysis of geometrical measures contributing to manifold capacitya, Effective geometric measures of manifolds representing different classes of odor pairs (AA: amino acids; BA: bile acids) based on data from naïve (N = 6) and trained fish (N = 25; all training groups combined). For all panels: ns, P ≥ 0.05; *, P < 0.05; **, P < 0.01; **, P < 0.001; see Supplementary Notes for more details. b, Change in manifold capacity in each training group relative to the naïve group for each pair of amino acids. Capacity was consistently higher in trained fish but statistical significance was reached only in one case (Phe-Trp in Phe+/Arg− fish; P = 0.002), possibly because the number of datapoints for comparison of single odor pairs in individual training groups is low. See Fig. 5 for statistical comparisons of pooled data.
*Manifold capacity analysis of simulated activity.Manifold capacity and geometric measures of neural manifolds in randomly connected spiking network models (“naïve”; n = 120 odor pairs from N = 8 network configurations) and in network models containing E/I-assemblies representing two input patterns (“trained”; Fig. 3; n = 120 odor pairs from N = 8 network configurations). For all panels: ns, P ≥ 0.05; *, P < 0.05; **, P < 0.01; **, P < 0.001; see Supplementary Notes for more details. Gray: results after shuffling of labels.
*Learning-related plasticity of odor representations in pDp of adult zebrafisha, Mean time course of inferred firing rates in response to each odor in naïve (N = 8 fish; left) and trained adult zebrafish (N = 9; right), averaged across all neurons, trials and fish. Gray shading indicates odor presentation (10 s); red shading shows time window for standard analyses (3 s). b, Response amplitude evoked by conditioned (CS+: Trp, CS−: Ala; left) and non-conditioned odors (Ser, His; right) was significantly higher in trained than in naïve fish. For all panels: ns, P ≥ 0.05; *, P < 0.05; **, P < 0.01; **, P < 0.001; see Supplementary Notes for more details. c, Amplitude histogram of responses to odors used for conditioning (Trp, Ala) in naïve (blue) and trained (orange) fish. d, Mean cosine distance between activity patterns evoked by different odors (3 trials each) in naïve (left) and trained fish (right). e, Difference between the cosine distance matrices in (d). f, Matrix of mean dE in naïve and trained adult zebrafish. g, Difference between dE matrices in f. h, dE between pairs of manifolds representing different categories of odors (Cond: conditioned odors [Trp, Ala]; Non-cond: other odors [His, Ser]). i, Matrix of mean dM in naïve (left) and trained fish (right). j, Difference between dM matrices in i. k, dM between pairs of manifolds representing different categories of odors. l, Manifold capacity was significantly higher in trained fish (gray: after shuffling of labels). m, Comparison of geometric measures contributing to manifold capacity in naïve and trained fish.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Hopfield J. J. Neural networks and physical systems with emergent collective computational abilities. Proc. Natl. Acad. Sci. U. S. A. 79, 2554–2558. (1982).6953413 10.1073/pnas.79.8.2554 PMC 346238 · doi ↗ · pubmed ↗
- 2Khona M. & Fiete I. R. Attractor and integrator networks in the brain. Nat Rev Neurosci 23, 744–766 (2022).36329249 10.1038/s 41583-022-00642-0 · doi ↗ · pubmed ↗
- 3Kohonen T. Self-organization and associative memory. (Springer, 1984).
- 4Pereira U. & Brunel N. Attractor Dynamics in Networks with Learning Rules Inferred from In Vivo Data. Neuron 99, 227–238 e 224 (2018).29909997 10.1016/j.neuron.2018.05.038PMC 6091895 · doi ↗ · pubmed ↗
- 5Aljadeff J., Gillett M., Pereira Obilinovic U. & Brunel N. From synapse to network: models of information storage and retrieval in neural circuits. Curr Opin Neurobiol 70, 24–33 (2021).34175521 10.1016/j.conb.2021.05.005PMC 8688181 · doi ↗ · pubmed ↗
- 6Burak Y. & Fiete I. R. Accurate path integration in continuous attractor network models of grid cells. P Lo S computational biology 5, e 1000291 (2009).19229307 10.1371/journal.pcbi.1000291 PMC 2632741 · doi ↗ · pubmed ↗
- 7Nieh E. H. Geometry of abstract learned knowledge in the hippocampus. Nature 595, 80–84 (2021).34135512 10.1038/s 41586-021-03652-7PMC 9549979 · doi ↗ · pubmed ↗
- 8Chung S. & Abbott L. F. Neural population geometry: An approach for understanding biological and artificial neural networks. Curr Opin Neurobiol 70, 137–144 (2021).34801787 10.1016/j.conb.2021.10.010PMC 10695674 · doi ↗ · pubmed ↗